
Abstract
We have used high-resolution field emission scanning electron microscopy with backscatter electron imaging to detect immunogold-labeled C5a and interleukin-8 (IL-8) receptors on human blood neutrophils. The receptors were labeled with receptor-specific antibodies in combination with secondary antibody conjugated to immunogold. When neutrophils were isolated in a “nonactivated” state, both of these receptor populations were expressed primarily in clusters on nonprojecting domains of the cell membrane. When these cells were double labeled for C5a and IL-8 receptors, intermixing of these receptor species in a common cluster was not found. When neutrophils were isolated in an “activated” state, by mixing the blood with N-formylmethionyl-leucyl-phenylalanine, the cells were seen to be elongated and ruffled at their anterior pole, but the C5a receptors did not disperse or redistribute on the surface of the peptide-activated cells. Analysis of the distribution of human C5a receptors expressed by transfected mouse L-cell fibroblasts showed the C5a receptors to be clustered, but expressed on nonprojecting and projecting domains of the cell surface. These observations provide new information on the topographical expression of leukocyte receptors involved in directing cell migration.
Keywords
S
This report describes the application of FESEM and immunogold labeling to studies of the spatial distribution of two chemoattractant receptors on human neutrophils: receptors for C5a, a product of activation of the fifth component of complement, and receptors for interleukin-8 (IL-8), a cytokine. These receptors belong to the “serpentine” family characterized by seven trans-membrane domains and signal transduction through G-proteins (Gerard and Gerard 1994; Horuk 1994). Experiments describe the distribution of C5a and IL-8 receptors on the surface of nonactivated and activated human neutrophils, and the distribution of human C5a receptors expressed by transfected mouse L-cell fibroblasts.
Materials and Methods
Leukocyte Isolation
Blood specimens were collected using EDTA as anticoagulant from seven healthy human male and female volunteers, with approval from the Committee on Use of Human Subjects in Research at the University of Minnesota. To obtain “nonactivated” neutrophils, 10-ml aliquots of the blood specimens were added immediately to 4 volumes of cold (4C) Dulbecco's calcium-free phosphate-buffered saline (DPBS) containing 0.25% paraformaldehyde (v/v). After incubation for 10 min at 4C, the diluted specimen was layered over 3-ml aliquots of cold Ficoll–Hypaque (sp. gr. 1.119 g/ml) and centrifuged for 30 min at 400 x g (Erlandsen et al. 1993). Addition of the fixative did not affect the separation of leukocyte subpopulations or the purity of the layered mononuclear cells or neutrophils. After washing, the leukocytes were re-suspended in DPBS containing calcium and 0.5% bovine serum albumin (BSA; Sigma, St Louis, MO) to a concentration of 3 × 106 cells/ml. To obtain “activated” neutrophils, the 10-ml blood specimens were immediately added to 0.1 ml 10-6 M N-formylmethionyl-leucyl-phenylalanine (FMLP; Sigma) in DPBS (Fernández–Segura et al. 1995). After 1 min, the blood was added to 40 ml 0.25% paraformaldehyde in DPBS and the neutrophils isolated as described.
Labeling and Processing of Leukocytes for FESEM
Chemotactic receptors were labeled by a two-step process involving primary receptor-specific antibody and an appropriate secondary antibody conjugated to 6-, 12-, or 18-nm colloidal gold (Jackson Immuno Research Laboratory; West Grove, PA). Primary antibodies recognizing C5a receptor were mouse monoclonal antibody or rabbit polyclonal antiserum (Morgan et al. 1993). Primary antibodies recognizing IL-8 receptors A and B were mouse MAbs 9H1 and 10H2, respectively (Chuntharapai et al. 1994), kindly supplied by Dr. K.J. Kim, (Genentech; South San Francisco, CA). The anti-IL-8 receptor antibodies were used in combination for total labeling of these receptors, on the basis of the preliminary observation that both IL-8 receptors exhibited the same clustered distribution pattern on the neutrophil surface. The specificity of the primary anti-C5a and -IL-8 receptor antibodies and their respective saturating doses have been established (Morgan et al. 1993; Chuntharapai et al. 1994). Secondary antibodies were used at a dilution of 1:10 (A520 = 2.0 for antibodies conjugated to 18- or 12-nm gold and 1.0 for antibody conjugated to 6-nm gold). Cells did not label with the secondary antibodies when preincubated with iso-type control antibody or antiserum, or when receptor-specific antibody was omitted.
Immunolabeling of leukocytes for chemoattractant receptor expression was done after the cells were attached to 4 × 9-mm glass chips coated with 0.1% poly-
Transfection of Mouse L-cells to Express Human C5a Receptors
The human C5a receptor gene (kindly provided by Dr. Craig Gerard, Harvard Medical School) was stably transfected into mouse L-cell fibroblasts (CCL 1.1; American Type Culture Collection, Rockville, MD), which have been transfected to express N-formyl peptide receptor for evaluation of ligand binding and signal transduction (Prossnitz et al. 1991). Transfection of the C5a receptor gene, ligated into the pSFFV-neoexpression vector (Fuhlbrigge et al. 1988), was performed using a calcium phosphate precipitation method kit (Stratagene; La Jolla, CA). Stable transfectants were selected by their resistance to 0.25 g/ml G418 sulfate (Life Technologies; Grand Island, NY). Transfected and nontransfected cells for microscopy were obtained by seeding sterilized chips with the cells in 24-well tissue culture plates. After 24 hr, the chips were removed, rinsed in DPBS, and immersed in 0.2% paraformaldehyde in DPBS. Labeling of C5a receptors was done as described for neutrophils. Nontransfected cells did not label with immunogold after exposure to anti-human C5a receptor antibody.
Scanning Electron Microscopy
Cells were examined at accelerating voltages of 3.0–4.0 keV in a Hitachi S-900 field-emission SEM equipped with a YAG crystal for high-resolution backscatter electron detection. Images were recorded on Polaroid type 55 film.
Results
C5a and IL-8 Receptors on Nonactivated Neutrophils
Neutrophils were isolated by immediate addition of the blood specimen to paraformaldehyde fixative, to avoid or minimize cell activation during cell isolation and processing, which might alter cell surface morphology and/or cause a redistribution of cell surface molecules.
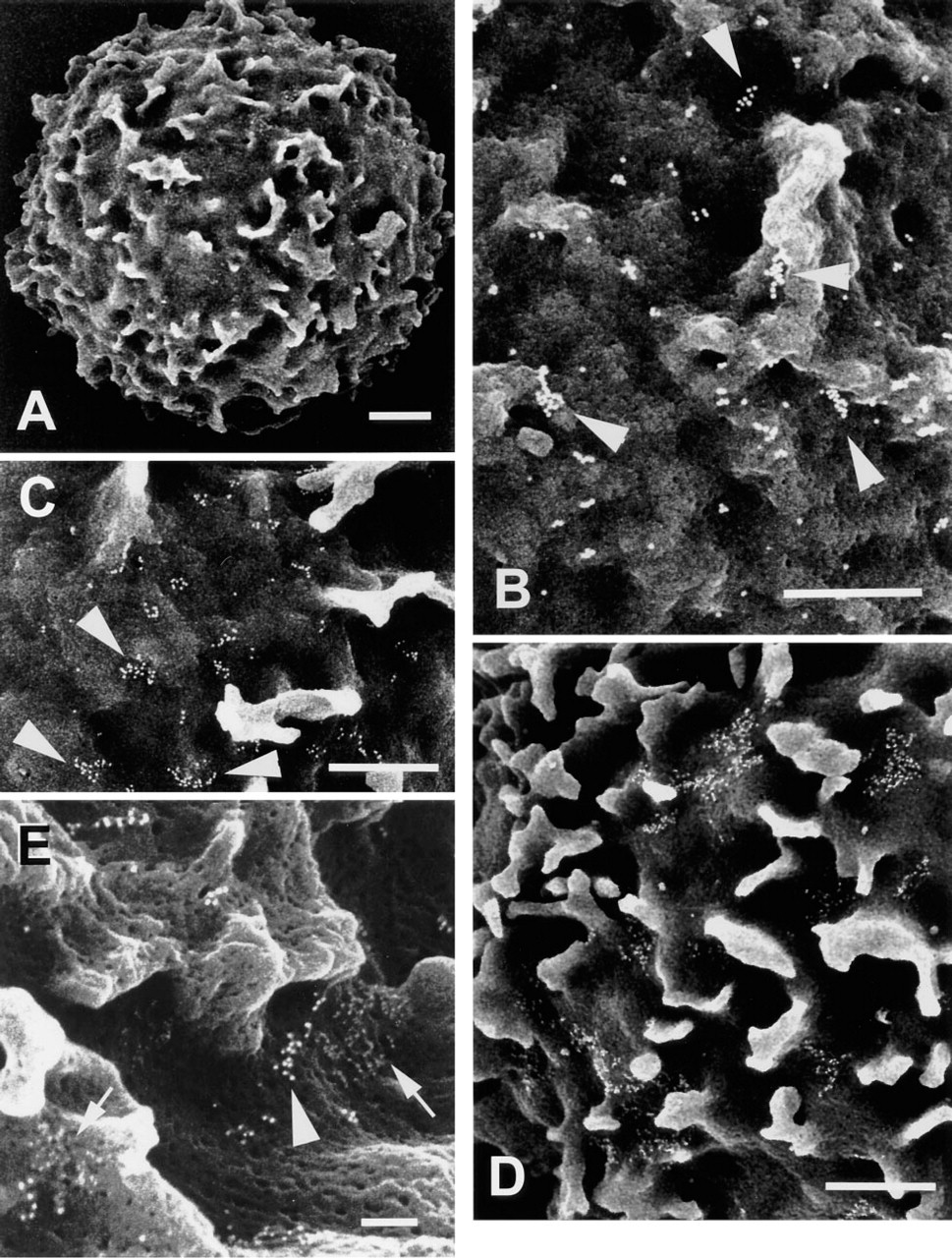
FESEM analysis of immunogold-labeled C5a and IL-8 receptors on nonactivated human neutrophils. (
Figure 1A illustrates the ruffled topography typical of neutrophils prepared by this methodology. Figures 1B–1D show typical distribution patterns of immunogold-labeled C5a receptors on neutrophils isolated from three of seven individuals (five men/two women) studied. Labeled receptors on these cells were occasionally seen as single gold particles but were most often seen as clusters composed of 3–15 (Figure 1B), 16–25 (Figure 1C), or <40 gold particles (Figure 1D). A similar diversity of labeling of C5a receptors was observed between cells isolated from the different donors. To obtain an objective measure of C5a clustering, the numbers of gold particles per cluster were counted on images of six different cells isolated from one of these donors (Figure 1B). Defining “cluster” as a group of gold particles in which individual particles are separated by a distance less than one particle diameter, 10% of the particles in these images were defined as singular, 30% were in clusters of 2–5, 22% were in clusters of 6–10, and 34% were in clusters of ≥11 particles.
The distributions of C5a and IL-8 receptor molecules on nonactivated neutrophils were compared by incubating the cells with primary rabbit polyclonal anti-C5a receptor and mouse monoclonal anti-IL-8 receptor antibodies, followed by secondary donkey anti-rabbit and rabbit anti-mouse immunoglobulin antibodies conjugated to immunogold. In Figure 1E, gold particles labeling either C5a receptors (12 nm) or IL-8 receptors (6 nm) are seen to be clustered. Further, no intermixing of these particles was observed in areas where particles representing the respective receptor populations were in close proximity (arrows) in this and three replicate experiments involving neutrophils isolated from different donors.
Influence of Cell Activation on the Distribution of C5a Receptors
To determine if cell activation might induce a redistribution of C5a receptors, neutrophils were stimulated by adding the blood specimen to DPBS containing the tripeptide chemoattractant FMLP (Fernández–Segura et al. 1995) before mixing with paraformaldehyde and cell isolation. Figures 2A–2C illustrate the morphology of a typical FMLP-activated cell (Figure 2A) and the comparable distribution of gold-labeled C5a receptors on the midbody (Figure 2B) and blunt posterior (Figure 2C) of such cells. Gold-labeled C5a receptors on the ruffled cell anterior (not shown) were distributed in a similar pattern. This observation indicates that brief activation of neutrophils by FMLP does not cause either a redistribution of C5a receptors along the long axis of the polarized cell or dissociation of the clustered receptors.
Distribution of Human C5a Receptors on Transfected Mouse L-cells
This experiment was conducted to determine if the clustered distribution noted for C5a receptors on neutrophils also occurs when these receptors are expressed in a “foreign” environment, represented here by mouse L-cell fibroblasts. Figure 2D demonstrates the multipolar morphology of these cells. Figures 2E–2G illustrate the distribution of the gold-labeled receptors on flat, ridged, or microvillous domains of the cell membrane, respectively. Note that the gold particles are predominantly in clusters on flat (Figure 2E), as well as ridged (Figure 2F) and microvillous domains (Figure 2G) of the cell membrane.
Discussion
In this report we describe the use of high-resolution FESEM in conjunction with immunogold labeling to visualize the cell surface distributions of receptors for the chemoattractants C5a or IL-8 on human neutrophils. To define the distribution of these receptor populations on naive circulating neutrophils, the neutrophils were isolated from blood specimens mixed with paraformaldehyde before cell isolation and exposure to processing conditions that might influence cell topography and/or surface molecule distribution (Hasslen et al. 1996). This procedure was chosen on the basis of our preliminary observation that neutrophils fixed by this methodology cannot be stimulated by chemotactic agents to polarize and the use of aldehyde fixation by others to prevent the redistribution of chemotactic peptide receptors on rabbit peritoneal exudate neutrophils (Sullivan et al. 1984).
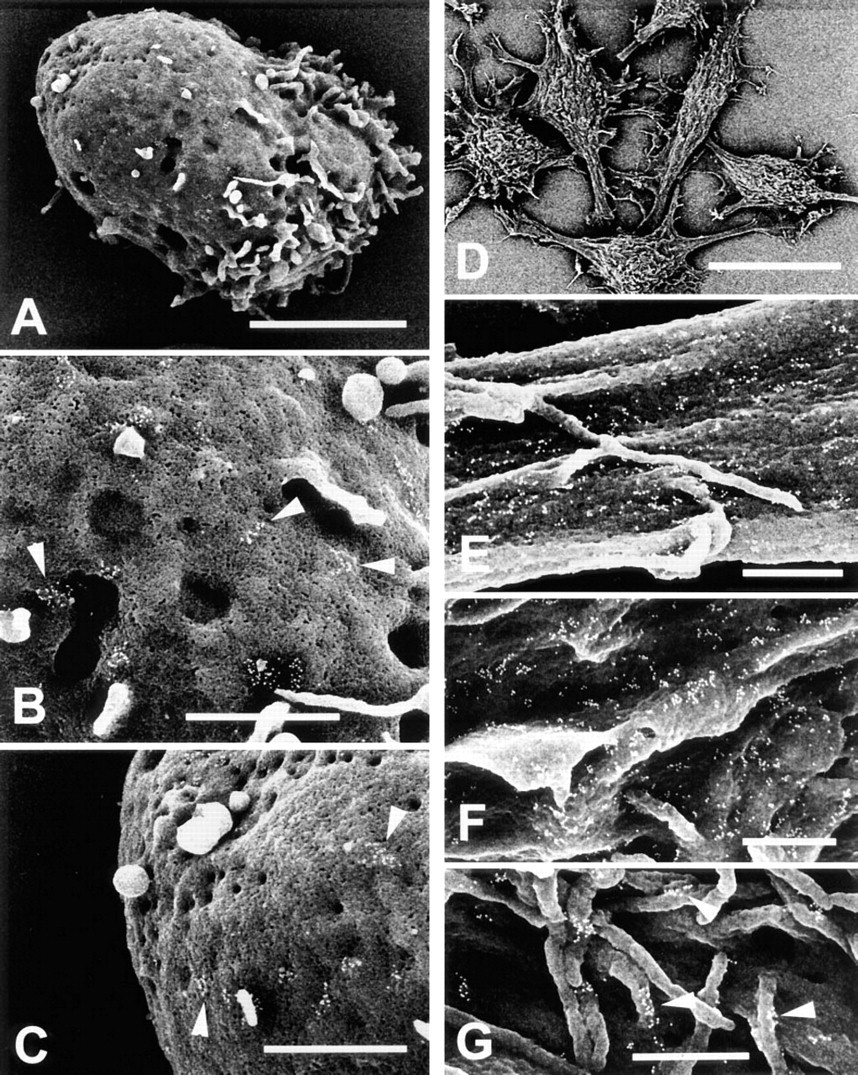
FESEM analysis of immunogold-labeled C5a receptors (
For neutrophils prefixed with paraformaldehyde, the immunogold-labeled C5a receptors were consistently observed as clusters of gold particles limited to the cell body, represented by nonruffled domains of the cell membrane (Figures 1B–1B). We have chosen “cluster” to define the observed arrangement of gold particles by comparison with computer-simulated arrangements of random, clustered, and regularly distributed points generated by nearest-neighbor analysis (Schwarz and Exner 1983). The number of gold particles per cluster showed some variation from cell to cell and donor to donor, but all cells examined showed a paucity of singlet gold particles. Cells double labeled for C5a and IL-8 receptors showed similar clustered distributions of gold particles bound to these receptor populations (Figure 1E), indicating that clustering is characteristic of receptors for two major neutrophil chemoattractants (Girard and Girard 1994; Horuk 1994).
One might argue that the clustered arrangement observed for these receptors is an artifact of cell activation or immunolabeling, but several observations provide circumstantial evidence that this is not so. The isolated, prefixed cells exhibited no morphological features of activation (polarization, increased ruffling, or spreading on the glass chip), indicating that fixation effectively preserved native neutrophil topography and receptor distribution. Thy-1 molecules expressed in very high density on murine lymphoid cells are diffusely distributed after labeling by the same methodology (Hasslen et al. 1996), suggesting that clustering of the C5a and IL-8 receptors on neutrophils was not induced by antibody bound to these cell surface molecules. The clustered arrangement of human C5a receptors expressed on the surface of transfected mouse L-cells (Figures 2E–2E) suggests that clustering is a native property of these receptor molecules. Finally, receptors for nerve growth factor on responsive cells are found preclustered on responsive cells, providing a precedent for receptor preclustering and immobility on a cell surface (Venkatakrishnan et al. 1991).
The clustered arrangement of C5a and IL-8 receptors on circulating neutrophils must reflect the common function and structural homology of these receptor populations (Girard and Girard 1994; Horuk 1994), but the functional significance of this molecular arrangement is less clear. The more random distribution of ferritin-labeled, serpentine N-formyl peptide receptors observed on rabbit neutrophils (Sullivan et al. 1984) suggests that clustering may not be essential to signaling for directed cell migration. However, accepting that C5a and IL-8 receptors are constitutively clustered, it appears likely that this arrangement serves the same role in signal transduction as ligand-induced oligomerization of single-pass hormone receptors (Wells 1994).
Information presently available also does not immediately identify the mechanism(s) involved in the clustered arrangement of C5a and IL-8 receptors. These mechanisms might include receptor–receptor interactions and/or interactions of receptors with accessory molecules in the cell membrane or components of the cytoskeleton in the cytoplasm. Whatever these mechanisms, they must prevent intermixing of receptor molecules (Figure 1E), and allow cell-specific distributions of the clustered receptors, i.e., limited to unruffled areas of the neutrophil surface (Figures 1B–1B), but including the cell body and projecting membrane domains of the transfected fibroblasts (Figures 2E–2E).
Although answers are not available to questions raised by observations described in this report, it is clear that knowledge of distributions of cell surface molecules obtained by FESEM can offer clues to how these molecules function and identify aspects of receptor expression and function that have not previously been considered.
Footnotes
Acknowledgements
Supported in part by grants from NIH (RO1 22374; RDN) and the Minnesota Medical Foundation (RDN, SLE).