
Abstract
The nuclear matrix, a proteinaceous network believed to be a scaffolding structure determining higher-order organization of chromatin, is usually prepared from intact nuclei by a series of extraction steps. In most cell types investigated the nuclear matrix does not spontaneously resist these treatments but must be stabilized before the application of extracting agents. Incubation of isolated nuclei at 37C or 42C in buffers containing Mg++ has been widely employed as stabilizing agent. We have previously demonstrated that heat treatment induces changes in the distribution of three nuclear scaffold proteins in nuclei prepared in the absence of Mg++ ions. We studied whether different concentrations of Mg++ (2.0–5 mM) affect the spatial distribution of nuclear matrix proteins in nuclei isolated from K562 erythroleukemia cells and stabilized by heat at either 37C or 42C. Five proteins were studied, two of which were RNA metabolism-related proteins (a 105-kD component of splicing complexes and an RNP component), one a 126-kD constituent of a class of nuclear bodies, and two were components of the inner matrix network. The localization of proteins was determined by immunofluorescent staining and confocal scanning laser microscope. Mg++ induced significant changes of antigen distribution even at the lowest concentration employed, and these modifications were enhanced in parallel with increase in the concentration of the divalent cation. The different sensitivity to heat stabilization and Mg++ of these nuclear proteins might reflect a different degree of association with the nuclear scaffold and can be closely related to their functional or structural role.
Keywords
I
Transmission electron microscopic investigations have demonstrated that the matrix is composed of three distinct domains: an outer lamina, an inner fibrogranular network, and residual nucleoli (Maraldi et al. 1986). However, depending on the cell type and on the isolation procedure, the inner network and/or nucleolar remnants can be absent, whereas the lamina is always present (Stuurman et al. 1992c). The nuclear matrix has been prepared from many tissues and cell lines but its existence in vivo is still under debate. This is mainly because nuclei are subjected to extensive manipulations before the final structure is obtained, and there is a substantial risk for creating in vitro artifacts (Cook 1988; Jack and Eggert 1992). Evan and Hancock (1985) demonstrated that when isolated nuclei are briefly exposed to the physiological temperature of 37C (e.g., during the nuclease digestion) a distinct subset of proteins became insoluble in buffers of high ionic strength. The term “heat-stabilization” of the nuclear matrix was introduced to denote this effect of temperature on solubility of nuclear polypeptides (Martelli et al. 1991). It occurs in many cell lines, but despite quite an intensive research the molecular mechanisms underlying it are yet to be elucidated (Martelli et al. 1994). Moderate heat exposure of isolated nuclei prevents the inner matrix network and residual nucleoli from dissociating when exposed to high ionic strength solutions.
Nevertheless, it has been argued that heat stabilization of the matrix could lead to the formation of artifacts (McConnell et al. 1987; Jack and Eggert 1992). Changes have been observed in the distribution of three nuclear scaffold proteins when heat treatment was employed to stabilize nuclei and nuclear scaffolds (Neri et al. 1997b). During the thermal stabilization of isolated nuclei, different concentrations of Mg++, ranging from 1 to 5 mM, have been employed (Boyle and Baluda 1987; Humphrey and Pigiet 1987; McConnell et al. 1987; Martelli et al. 1990; Belgrader et al. 1991; Kaufmann and Shaper 1991; Martelli et al. 1991; Neri et al. 1994). It is established that different concentrations of Mg++ have a marked effect on nuclear structure (e.g., Leake et al. 1972; Laval and Bouteille 1973; Dixon and Burkholder 1984; Papa et al. 1988). The addition of 5 mM Mg++ was able to prevent the redistribution of DNA topoisomerase II β after in vitro stabilization at 37C but not at 42C, showing that heat stabilization in a buffer without Mg++ cannot maintain the original distribution of the protein within the nucleus (Neri et al. 1997b). With the above in mind, we decided to investigate by indirect immunofluorescence experiments and confocal laser scanning microscopy analysis whether different Mg++ concentrations present during incubation of isolated nuclei at 37C or 42C could differently affect the spatial distribution of several polypeptides that are among the constituents of the nuclear matrix. The proteins we have analyzed are components of the inner network: 125- and 160-kD polypeptides (de Graaf et al. 1992; Marugg 1992), a 126-kD component of a class of nuclear bodies (Stuurman et al. 1992b) also known as ND10 or Kr bodies (Lamond and Carmo–Fonseca 1993; Zuber et al. 1995), an RNP antigen, and a constituent of splicing complexes (Clevenger and Epstein 1984).
Marked differences were detected in the behavior of the various antigens related to different stabilizing temperatures and/or Mg++ concentrations. The results emphasize the necessity to carefully perform adequate morphological controls at every step of the isolation protocols to obtain the insoluble nuclear matrix.
Materials and Methods
Sources of Antibodies
Monoclonal antibodies (MAbs) to inner nuclear matrix proteins of 160 kD (referred to as p160) and 126-kD component of nuclear bodies (clone 5E10) were a kind gift from Drs. R. van Driel and L. de Jong (E. C. Slater Institute for Biochemical Research, Amsterdam, The Netherlands). The antibodies have been previously described (de Graaf et al. 1992; Stuurman et al. 1992b). The MAb to 125-kD inner matrix protein (referred to as p125) has been described elsewhere (Marugg 1992). MAbs recognizing the 105-kD splicing component or the RNP polypeptide (Clevenger and Epstein 1984) were purchased from Chemicon International (Temecula, CA).
Cell Culture
K562 human erythroleukemia cells were grown in RPMI-1640 medium supplemented with 10% newborn calf serum at 37C in a humidified atmosphere containing 5% CO2. They were seeded at a density of 105/ml and used 4 days later, when they reached a density of 106/ml. It is important to stress that at this stage most of the cells (88%) were arrested in the G0/G1 phase of the cell cycle, as detected by flow cytometric control (data not shown).
Isolation of Nuclei
Cells were washed once in Dulbecco's PBS, pH 7.4, free of Mg++ and Ca++. They were then resuspended (1.5 × 107 cells/ml) in TM-2 buffer (10 mM Tris-HCl, pH 7.4, 2 mM MgCl2, 1 μg/ml aprotinin, 1 μg/leupeptin) and incubated for 5 min at 0C. Triton X-100 was added to 0.5% and cells were sheared by one passage through a 22-gauge needle fitted to a 30-ml plastic syringe. Nuclei were recovered by low speed centrifugation (8 min at 400 × g) and washed once in TM-2. They were then resuspended at 2 mg/ml in ST buffer (10 mM Tris-HCl, pH 7.4, 0.25 M sucrose, and protease inhibitors as above) containing different concentrations of Mg++ (2, 3.5, 5 mM) and incubated for 20 min at 0C, 37C, or 42C.
Immunofluorescent Staining
Cells in PBS and nuclei in ST buffer (containing the appropriate Mg++ concentration) were immediately plated onto 0.1% poly-
Confocal Laser Scanning Microscopic (CLSM) Analysis
Samples were imaged by a PHOIBOS 1000–SARASTRO (Molecular Dynamics; Sunnyvale, CA) CLSM mounted on a Nikon Optiphot microscope (Nikon; Tokyo, Japan). This confocal system was coupled with a 25-mW multiline argon ion laser as a light source, with a major line at 488 nm selected with a bandpass filter to reveal FITC signal. The laser power was tuned at 10 mW to obtain the highest light stability and the laser beam was attenuated to 30% of transmission with a neutral density filter to limit bleaching of the FITC fluorescence. Samples were observed with a × 100, 1.4 NA planapochromat objective lens. To obtain the highest resolution we have employed the lowest laser light wavelength suitable and the highest NA objective [(0.46 λ)/NA]. The refractive index of the immersion oil was 1.518 (Nikon). The oil was dissolved up to a concentration of 20% in the mounting medium to minimize distortion of the confocal spot during the laser beam penetration inside the specimen. In the detection path, the emitted fluorescent light was focused on a back pinhole aperture with a diameter of 50 μ in front of the detector, a photomultiplier tube (PMT). To block any unwanted contribution signal, when FITC was observed, a 515-nm longpass filter was inserted before the PMT as a barrier filter. The PMT was set at 912 mV. Settings were rigorously maintained for all experiments. Images were acquired, frame by frame, with a scanning mode format of 512 × 512 pixels. Pixel values were recorded in the range of 0–255 (8 bits) as previously described (Neri et al. 1997b). Images were reconstructed as follows. Slides were scanned from left to right on the x–y plane moving down with steps of 0.3 μ from top to bottom in the z direction of samples, using the motor drive focusing system to a position unaffected by the last horizontal pass. Each frame had a scan time of 0.5 sec and every optical section was the result of a single scan. The microscope table was set manually so that the first z section was collected just at the top of the structures to be analyzed.
Image Processing Analysis
Digitalized optical sections, i.e., z series of confocal data (“stacks”), were transferred from the CLSM to the graphics workstation Indigo Irix x524 (Silicon Graphics; Mountain View, CA) and stored on the graphics workstation with a scanning mode format of 512 × 512 pixels and 256 gray levels. The image processing was performed using the Image-Space software (Molecular Dynamics). To reduce the unwanted background noise generated by the photomultiplier signal amplification, all the image stacks were treated with a two-dimensional filter (Gaussian filtering) that was carried out on each voxel, with a mask of 3 pixels in the x, y and z direction (3 × 3 × 3). The FITC signal was elaborated to optimize the contrast, the brightness, and the intensity of the images.
Photographs were taken by a digital video recorder Focus ImageCorder Plus (Focus Graphics; Foster City, CA) using 100 ASA TMax black and white film (Kodak; Rochester, NY).
Results
CLSM Analysis
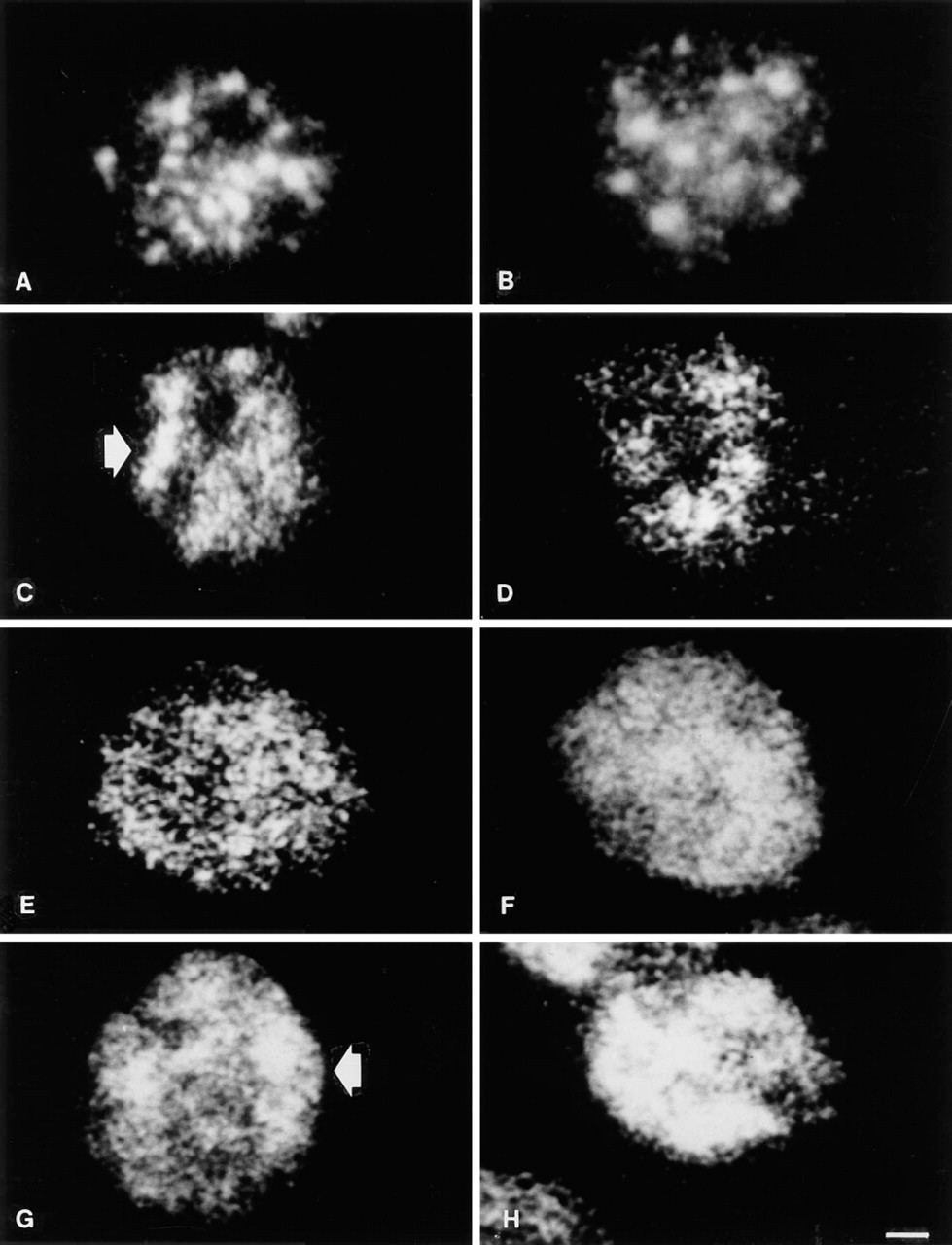
p125. In intact cells the distribution of the p125 nuclear matrix component was characterized by the presence of large and brilliant spots mainly located in the inner nucleoplasm, together with a faint fluorescence dispersed also at the nuclear periphery but not in the nucleolar areas as detected by phase contrast (not shown) (Figure 1A). Nuclei exposed to 0C showed an almost identical pattern (Figure 1B). In 2 mM MgCl2-prepared nuclei, after the stabilization at 37C the large dots were almost abolished and were substituted by enlarged and irregular fluorescent areas (Figure 1C). Many dots were present in the nucleoplasm, either rounded or irregularly shaped, that replaced the large spots. Nucleoli were still observable as negative areas. At 42C the pattern was characterized by very large and irregular spots associated with smaller dots of irregular size and shape that made it difficult to distinguish nucleoli (Figure 1D). The effect of 37C on nuclei prepared in 3.5 mM MgCl2 resulted in the distribution in the whole nucleus of dots that in some cases were fused together to form larger masses of irregular shape (Figure 1E). Nucleoli were hardly detectable. Homogeneous labeling was observable in 42C nuclei prepared with 3.5 mM MgCl2 in the buffer. It was also diffused at the periphery and showed some accumulation in the center. Nucleoli were not recognizable (Figure 1F). Five mM MgCl2 showed, after 37C stabilization, the scattering of the 125-kD protein all over the nucleoplasm, with a few areas of condensation mainly in the peripheral region (Figure 1G). The same MgCl2 concentration in 42C exposed nuclei determined the clumping of most of the fluorescent spots into large irregular masses distributed both in the inner nucleoplasm and at the nuclear periphery (Figure 1H). Nucleoli were undetectable in 5 mM MgCl2-treated nuclei.
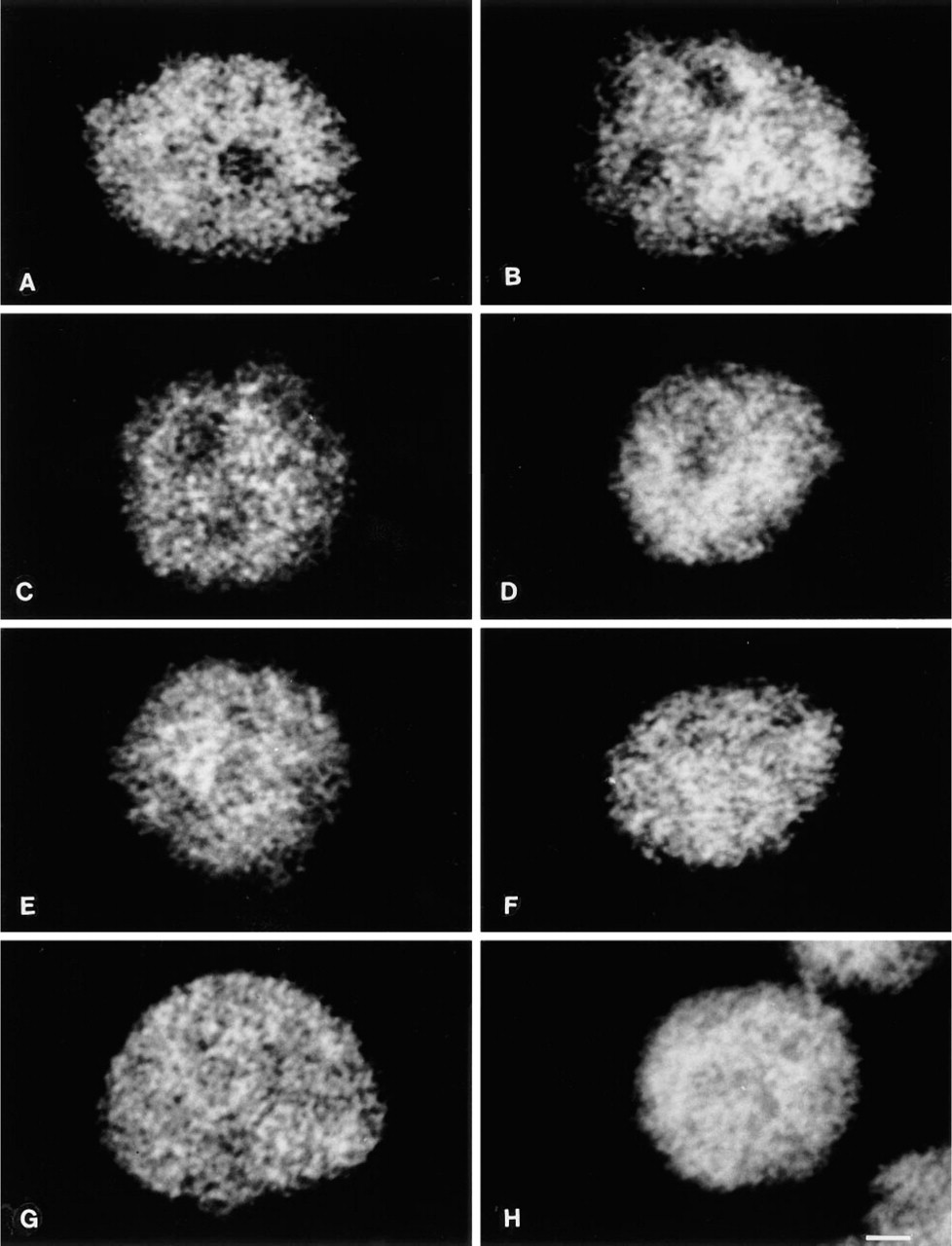
p160. The distribution of the 160-kD nuclear matrix protein in intact cells revealed a diffuse granular distribution with tiny dots throughout the nucleus, with no significant staining of the nucleoli (Figure 2A). The treatment at 0C did not modify this pattern except for a moderate degree of fluorescence condensation in the center of the nucleus (Figure 2B). When treated at 37C in a buffer containing 2 mM MgCl2, nuclei maintained the same protein distribution (Figure 2C) which was also observed at 42C (Figure 2D). Exposure at both 37C and 42C in 3.5 mM MgCl2 solution showed the diffusion of the granular meshwork also in the nucleolar areas that were barely detectable at 37C and not detectable at 42C (Figures 2E and 2F). A very similar aspect was observed after nuclear stabilization with 5 mM MgCl2 buffer: 37C and 42C treatment distributed the granules all over the nucleoplasm (Figures 2G and 2H), with the presence at 42C of a very faint fluorescence associated with the granules.
Single confocal optical sections of intact cell (
Single confocal optical sections of intact cell (
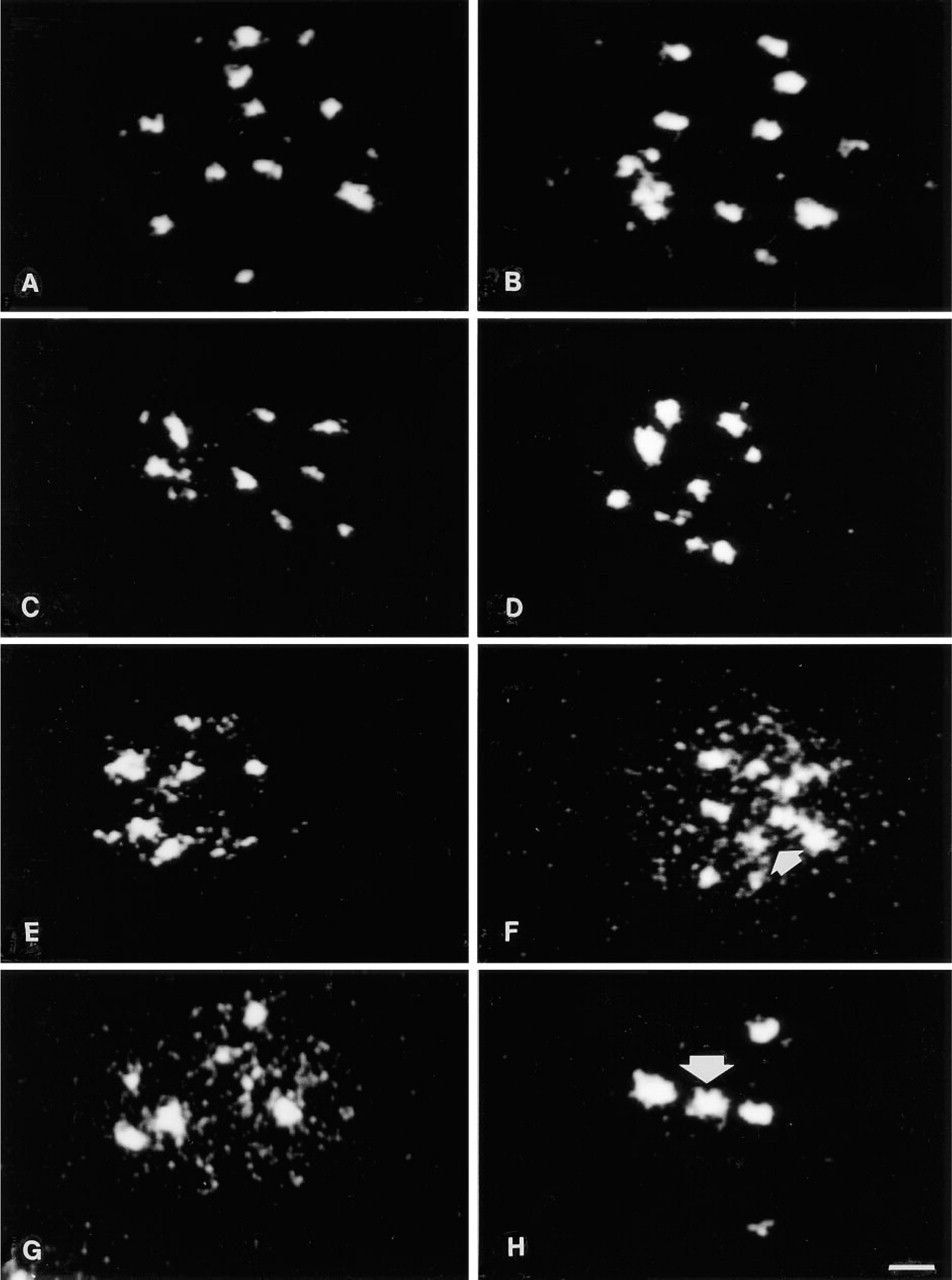
126-kD Component of Nuclear Bodies. In intact cells the staining was concentrated in 10–12 isolated spots distributed both in the center and in the periphery of the nucleus. It should be remembered that we selected the equatorial plane section through the nucleus that contains the majority but not all of the spots. The spots were similar in size but they displayed a different shape, with nonlinear edges (Figure 3A). The treatment at 0C did not significantly alter the described pattern (Figure 3B). The distribution area of the spots and their number were reduced by the stabilization at 37C and at 42C in buffer containing 2 mM MgCl2 (Figures 3C and 3D). A similar reduction of spot number and distribution area was induced by 3.5 mM buffers at both 37C and 42C (Figures 3E and 3F). In addition, we detected the appearance of some tiny dots at 37C that became more numerous at 42C. The latter stabilization was also characterized by the presence of large and very irregular spots, mainly located in the inner nucleoplasm. Many granules and tiny dots were distributed all over the nucleoplasm in 5 mM MgCl2-treated and 37C-exposed nuclei (Figure 3G). A few isolated spots (four or five) remained both in the nuclear center and periphery. Stabilization at 42C abolished the granules and the dots and induced a localization of the spots, mainly in the inner nucleoplasm (Figure 3H). The spot edges were irregular, their number was reduced, and their size was clearly enlarged in comparison with intact cells.
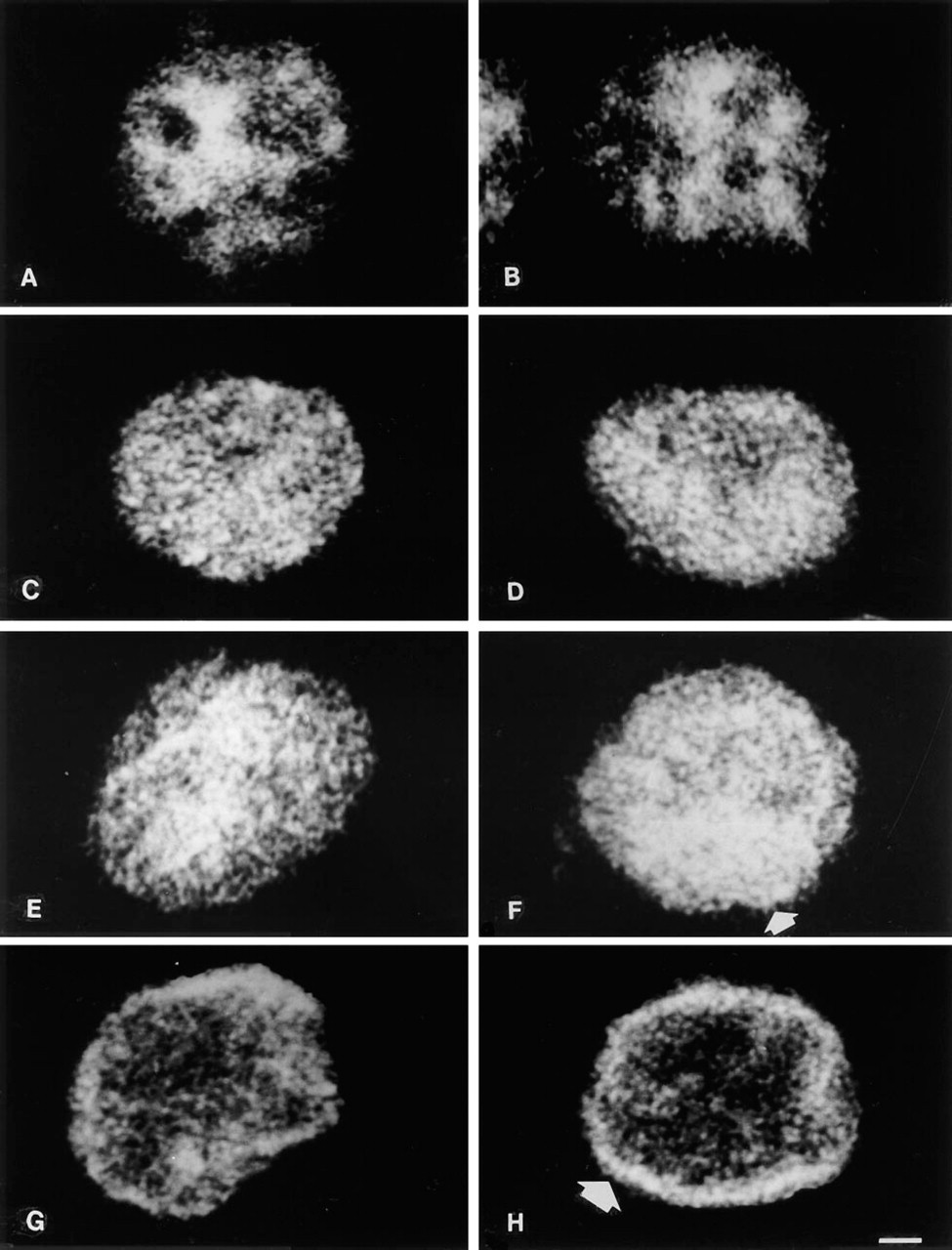
RNP. Discrete labeling of the nucleoplasm with tiny dots was observed in intact cells reacted with the anti-RNP antibody (Figure 4A). Condensation of fluorescence in the inner part of the nucleus was often detectable. Nucleoli were not stained as well as irregular areas at the nuclear periphery. Nuclei incubated at 0C showed an identical pattern (Figure 4B). Marked changes were induced by 2 mM MgCl2 at 37C and at 42C. The tiny dots were replaced by larger granules of different sizes scattered throughout the entire nucleoplasm, including the nucleolar areas that were no longer detectable (Figures 4C and 4D). A very similar pattern was observed after stabilization in 3.5 mM buffer. At 37C the granules were more concentrated in the center of the nucleus (Figure 4E), but at 42C this concentration was seen also at the nuclear periphery (Figure 4F). Fluorescence distribution was altered dramatically in the presence of 5 mM MgCl2. Exposure at 37C confined most of the nuclear fluorescence to the periphery, associated with some irregular granules also located in the center (Figure 4G). The inner nucleoplasm was almost empty after 42C treatment, and an evident ring of fluorescence was concentrated at the nuclear edge (Figure 4H), very reminiscent of the patterns given by antibodies to nuclear lamins (Hozak et al. 1995).
105-kD Splicing Component. Fifteen to 20 discrete large specks were observable in intact cells. They were brilliant and alternated with unlabeled areas in the nucleoplasm, some of which corresponded to nucleoli (Figure 5A). A comparable antigen distribution was seen in nuclei exposed to 0C (Figure 5B). In the case of 2 mM MgCl2 buffers, rare and large specks (two to four) were still observable associated with many brilliant granules of irregular size and shape. Both at 37C (Figure 5C) and at 42C (Figure 5D), the unlabeled areas were very much reduced and barely detectable. The use of buffers containing 3.5 mM MgCl2 completely disarranged the large spots, which were substituted by wide irregular masses concomitantly with the appearance of brilliant granules in 37C-exposed nuclei (Figure 5E). Conversely, in 42C-stabilized nuclei only the granules were observable, sometimes partially fused together at the nuclear periphery (Figure 5F). The appearance of large and homogeneously stained masses was caused by the combination of 37C incubation and 5 mM MgCl2 buffers. Between the masses an irregular fibrogranular meshwork was observable (Figure 5G). Treatment at 42C was characterized by large irregular masses in the center of the nucleus, where a fibrogranular aspect could also be distinguished. Isolated granules were present in all the remaining nucleoplasm (Figure 5H).
Quantitative Analysis
Table 1 shows the results of quantitative analysis performed on the various samples we analyzed, immunostained for the five antigens studied. To minimize as much as possible any bias in the analysis, we selected a standard set of cell images and then randomly compared individual test images of cells and nuclei with the standard set. For each experiment, 50 cells and nuclei were manually counted. A statistical test was performed by comparing the number of structures exhibiting the typical immunofluorescent pattern identified in intact cells with the number of those showing a different pattern. The data revealed that highly significant differences were detected in almost all of the samples after different treatments, thus demonstrating that the changes in the distribution of antigens discussed above were indeed present in the great majority of isolated nuclei.
Single confocal optical sections of intact cell (
Intact cell (
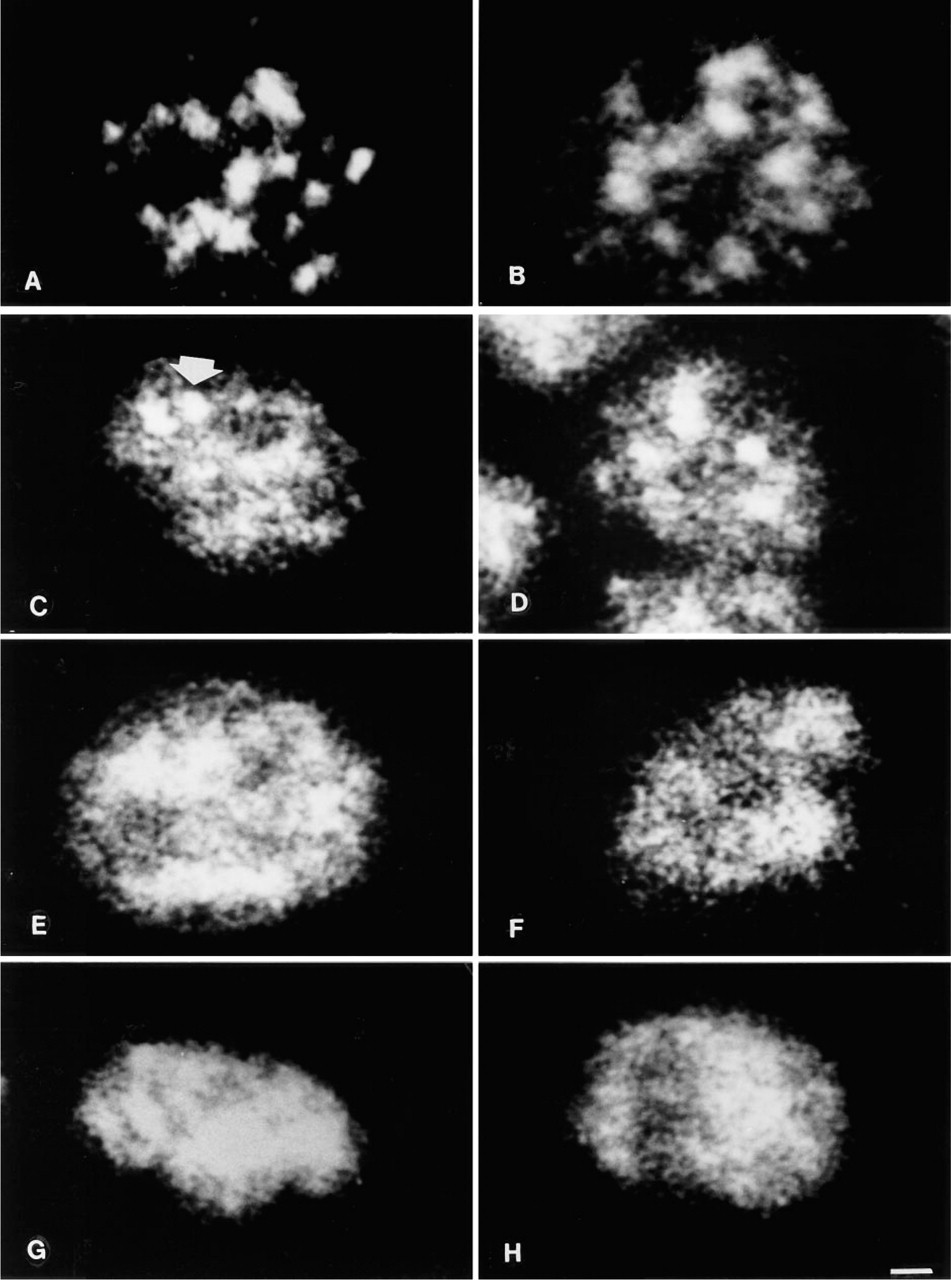
Intact cell (
Percentage of cells and isolated nuclei displaying immunofluorescent staining different from the typical pattern described for intact cells, as described in the Results section for each of the antibodies employed in this study a
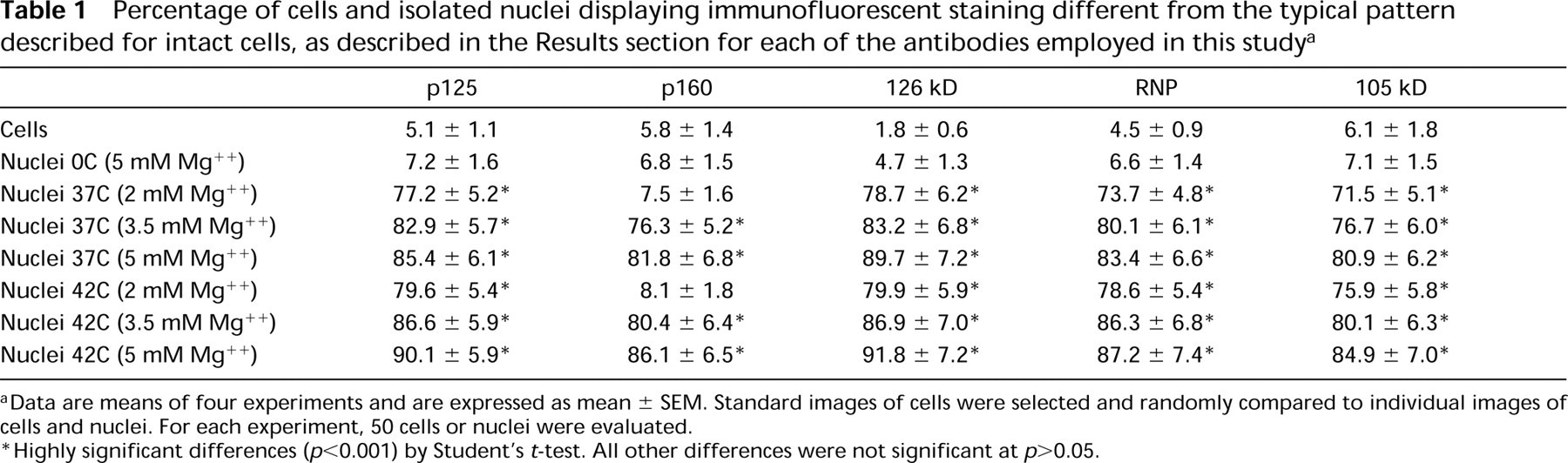
aData are means of four experiments and are expressed as mean ± SEM. Standard images of cells were selected and randomly compared to individual images of cells and nuclei. For each experiment, 50 cells or nuclei were evaluated.
∗Highly significant differences (p<0.001) by Student's t-test. All other differences were not significant at p>0.05.
Discussion
Nuclei are usually isolated by homogenizing cells in hypotonic buffers because chromatin aggregates and gels in isotonic salt solutions. Such an unphysiological environment has adverse effects on 30-nm chromatin fiber organization and on heterochromatin structure, and extracts many nuclear proteins. To circumvent these problems, stabilizing divalent cations such as Mg++ (Cook 1988) are added to the solutions, but the concentration of Mg++ ions employed varies over a wide range, thus affecting in a different manner the ultra-structural appearance of the nucleus (Papa et al. 1988).
Several reports have been published dealing with artifacts that may occur during nuclear matrix preparations. In these reports the investigators have taken into consideration the effect on nuclear matrix structure of RNAse A (Kaufmann et al. 1981; Belgrader et al. 1991), of reducing or oxidative chemicals (Dijkwel and Wenink 1984; Kaufmann and Shaper 1984; Stuurman et al. 1992a), and of different extracting agents (Belgrader et al. 1991; Eberharter et al. 1993). Surprisingly, however, the effect of different concentrations of Mg++ ions during nuclear preparation to obtain the matrix has never been investigated. Only the effect of the ionic strength in general has been examined (Boyle and Baluda 1987). Moreover, all of the aforementioned investigations dealt with the ultra-structural morphology of the nuclear matrix prepared under different conditions and its overall protein composition. They never tackled the issue of studying the effect of the preparation protocols on the subnuclear distribution of specific nuclear matrix components.
In this study we have systematically examined whether or not different concentrations of Mg++ ions also affect the spatial distribution of the protein component of the nuclear matrix during heat-induced stabilization of isolated nuclei. In fact, it has previously been reported that not only the 2D but also the 3D spatial distribution of DNA Topoisomerase II β is heavily modified after stabilizing nuclei either at 37C or 42C in the absence of Mg++ (Neri et al. 1997b). To this end we used confocal laser scanning microscopy, which was able to reveal marked differences in the fluorescent patterns when nuclei were exposed to temperatures causing stabilization of the matrix. In the past we had already demonstrated that CLSM analysis reveals changes in the distribution of two of the antigens (p125 and p160) analyzed in the present work, which could not be fully appreciated with a conventional epifluorescence microscopy (Neri et al. 1994, 1995a).
We analyzed structures derived from a homogeneous cell population mostly in the G0/G1 phase of the cell cycle. In the future it would be interesting to study if the observed effects occur in all the various stages of the interphase, using a synchronized cell population.
The statistical analysis corroborated the data obtained by visual inspection, thus confirming the quality of the microscopic procedure and the significance of the results.
It appears that for some antigens (p160, RNP antigen, 105-kD splicing component) the Mg++ concentration is more important than the temperature at which the stabilization is performed. Changes in the distribution of these antigens were seen concomitantly with the increase in the Mg++ concentration. However, the two different temperatures employed did not have any effect. On the other hand, for other antigens (such as p125 and the 126-kD constituent of nuclear bodies) there is an additional effect of temperature because at the same Mg++ molarity one can see marked differences between samples incubated at 37C and those treated at 42C. It is also important to emphasize that two RNA metabolism-related proteins (RNP and 105-kD) show different sensitivities to the stabilization method employed. For RNP, dramatic changes occur even at 2 mM Mg++, whereas in the case of the 105-kD they are more subtle because some large specks are preserved at this concentration. Such a different behavior might reflect a tighter association of 105-kD with the structural components of the scaffold. In this sense, our results are in good agreement with the data reported recently by Bisotto et al. (1995), showing a tighter association of SC-35 splicing component with the nuclear matrix than that of snRNP antigens.
A comparison between two nuclear matrix proteins, p160 and p125, also showed in this case not only a different fluorescent pattern but also a different sensitivity to Mg++ concentrations. At 2 mM p160 was unaltered, whereas p125 was already completely disarranged. In the past these proteins have been shown to undergo similar modifications during in vitro heat stabilization at 37C (Neri et al. 1994) or different variations after heat shock in vivo (de Graaf et al. 1992; Neri et al. 1995b).
It should also be emphasized that when the nuclei are kept at 0C, even the highest Mg++ concentration employed (i.e., 5 mM) never induces redistribution of nuclear components. This indicates that Mg++ ions do not have an inherent capacity to induce changes but rather that they act only when they are present during the thermal stabilization, also taking into account that heat changes the subnuclear distribution of nuclear matrix or scaffold proteins prepared from nuclei isolated in a buffer not containing Mg++ ions (Neri et al. 1997a, c). In this connection, it should be remembered that when nuclei were kept at 0C in the absence of Mg++ we never detected changes in the spatial distribution of p160, p125, and 126-kD constituents of nuclear bodies (Neri et al. 1997c) or in the splicing component and the RNP constituent (unpublished experiments). On the other hand, heat treatment without Mg++ does not alter the distribution of p160 antigen (Neri et al. 1997c) but changes the immunofluorescence patterns of the other polypeptides, p125 and 126-kD constituent of nuclear bodies (Neri et al. 1997c) and of the splicing component and the RNP constituent (unpublished experiments). In addition, we have also reported that for some proteins, such as topoisomerase II β, Mg++ can prevent antigen redistribution induced by 37C heat exposure but is ineffective if the temperature is raised to 42C (Neri et al. 1997b). All these results show that the behavior of nuclear antigens to various combinations of Mg++ ions and temperature is extremely variable and must be carefully evaluated each time to optimize the isolation protocol. We feel that such an issue should not be neglected in future studies in which the nuclear matrix will be prepared. Conceivably, because of the artifacts that are induced in nuclear structure by Mg++ ions and heat stabilization, a re-evaluation of the functions that are found associated with the nuclear matrix after similar treatments appears desirable (Martelli et al. 1996). The method of isolating the nucleokeleton proposed by Jackson and Cook (1985, 1986a, b), which entails encapsulation of cells in agarose beads, permeabilization in physiological buffers, and digestion of chromatin with restriction enzymes with subsequent electroelution of DNA fragments, is believed by many investigators to be at present the most gentle procedure devised, because it does not require exposure of structures to either Mg++ or heat, factors that undoubtedly cannot preserve the original nuclear structure. However, it should be considered that electroelution of DNA fragments after nuclease digestion may produce oxidative stress and macromolecular crosslinking.
The different sensitivity to Mg++ or heat displayed by different antigens renders the elaboration of a common protocol for isolating the nuclear matrix more challenging. Clearly, morphological controls at every step of the procedure are necessary to rule out the occurrence of artifacts that could also affect the functional properties of the individual polypeptides.
Footnotes
Acknowledgements
Supported by Italian CNR grant PF ACRO and 94.–00413.CT12, by Fondi AIRC (Associazione Italiana Ricerca sul Cancro) 1995 and 1996, and by Italian MURST 60% grants to Università di Trieste and Ferrara.
We thank Giovanna Baldini for the illustrations. We are grateful to Drs R. van Driel and L. de Jong (E.C. Slater Institute for Biochemical Research, Amsterdam, The Netherlands), for supplying the antibodies.