
Abstract
Synaptotagmin I (STG I) is a Ca2+ sensor and one of the synaptic vesicle proteins that mediate exocytosis. To determine the mechanism of release of large granules from mast cells, we studied by immunohistochemistry the presence of STG I in mast cells in normal human tissues simultaneously with the mast cell markers mast cell tryptase (tryptase) and c-kit. The tumor cells of systemic mast cell disease (SMCD) and a human mast cell leukemia cell line (HMC-1) were also examined. Human mast cells in normal tissues and the tumor cells of SMCD expressed STG I as well as mast cell tryptase (tryptase) and c-kit. STG I mRNA and its products in HMC-1 were examined by RT-PCR analysis and immunocytochemistry, respectively. STG I expression in HMC-1 cells was compared with that in cells stimulated and non-stimulated by phorbol 12-myristate 13-acetate and also with that in NB-1 and PC12 cells, known to express STG I. STG I mRNA was detected in both non-stimulated and stimulated HMC-1 cells and in NB-1 and PC12 cells. STG I immunoreactivity was weaker than NB-1 or PC12 immunoreactivity. However, it increased in the stimulated HMC-1 cells. Mast cells expressed STG I in various states. STG I may mediate exocytosis of large granules in mast cells.
Keywords
M
It was recently reported that mast cells express rab3a and rab3d proteins and interfere with a rate-limiting step in IgE receptor-stimulated exocytosis (Oberhauser et al. 1994; Smith et al. 1997). Rab3s are synaptic proteins postulated to regulate exocytotic vesicle traffic in neurons and many secretory cells (Sudhof 1995). Furthermore, Baram et al. (1998) demonstrated that rat peritoneal mast cells and rat basophilic leukemia cells express STG I by Western blotting and immunocytochemistry. STG-I localized almost exclusively to their secretory granules. This encouraged us to examine whether STG I mediates human mast cell exocytosis.
We conducted an immunohistochemical study of STG I in mast cells of various normal and neoplastic human tissues and in systemic mast cell disease (SMCD). SMCD is a rare disease characterized by a generalized abnormal infiltration of mast cells, most notably involving the bone marrow, lymph node, liver, spleen, and other parenchymal organs, with or without cutaneous involvement. There are four forms of SMCD: (a) an indolent form limited to the skin, with a favorable prognosis; (b) a form with associated hematological disorders; (c) an aggressive form with a poor prognosis, which exhibits visceral and bone involvement that may or may not be associated with skin lesions; and (d) mast cell leukemia (Warnke et al. 1995). C-kit is also a useful marker in the differential diagnosis of SMCD involving the bone marrow (Natkunam and Rouse 2000). Furthermore, we verified the expression of STG I mRNA and its products in HMC-1 cells that were established from the peripheral blood of a patient with mast cell leukemia by Butterfield et al. (1990) with or without stimulation by phorbol 12-myristate 13-acetate (PMA).
Materials and Methods
Histochemistry and Immunohistochemistry
Ten normal human tissues, including stomach, colon, pancreas, adrenal medulla, skin, salivary gland, and uterine cervix obtained by surgery, were fixed in 10% buffered formalin and embedded in paraffin. To avoid nonspecific staining, the histochemistry and immunohistochemistry were based on Shiltz et al. (1993). Mast cells in these tissues were identified by immunohistochemistry of tryptase and c-kit. In a case of SMCD, Dr. Maruyama (Hirakata, Japan) kindly supplied paraffin-embedded tissues of the kidney and stomach with lymph node. This case was of Type c, with an elevated plasma histamine level of 3.59 ng/ml (normal <0.8 ng/ml). An autopsy had revealed mast cell infiltration mainly in the spleen, liver, kidney, lymph nodes, and bone marrow. This case has been previously reported by Maruyama et al. (1998).
A human mast cell line (HMC-1) was cultured in Dulbecco's medium (IMDM) with 10% heat-inactivated fetal bovine serum. A rat pheochromocytoma cell line (PC12) and a human neuroblastoma cell line (NB-1), both known to contain STG-I (Shoji–Kasai et al. 1992), were used for positive controls. PC12 and NB-1 cells were cultured in RPMI 1640. These cells were stimulated with PMA 10 ng/mL for 2, 5, and 8 hr and the expression of STG I was compared with that of non-stimulated cells by immunocytochemistry and RT-PCR analysis.
Immunohistochemical Staining
Mast cells contain heparin, which is a proteoglycan. Proteoglycans contain many sulfate and carboxyl groups that impart a highly negative charge to molecules such as avidin and immunoglobulins. Furthermore, nonspecific labeling of mast cells results from an ionic interaction between the (Fab′)2 segments of antibodies and the heparin constituent of the mast cell secretory granules (Schiltz et al. 1993). Thus, histochemistry and immunohistochemistry in mast cells have been carefully performed.
We used the Histofine SAB-PO(M) kit (Nichirei, Tokyo), and Simple Stain Max (M) kit (Nichirei), which is an amino acid polymer labeled by peroxidase and rabbit anti-mouse IgG Fab' for immunohistochemistry. Sections were dewaxed in xylene, washed in ethanol, and incubated in a 0.06% hydrogen peroxide–methanol solution to block endogenous peroxidase activity. For the SAB-PO kit, but not the Simple Stain kit, nonspecific staining was blocked with 10% normal rabbit serum. Then the sections were incubated with the primary antibodies at the appropriate dilution for 18 hr at 4C. After a wash in PBS, the sections were incubated in biotin-labeled rabbit anti-mouse IgG + IgA + IgM antibody (10 μg/ml) for 30 min at room temperature (RT). After another wash in PBS, the sections were incubated in peroxidase-labeled streptavidin for 30 min at RT.
For the Simple Stain kit, the procedure was the same as for the SAB-PO kit until the first antibodies were applied. The sections were incubated in the solutions containing amino acid polymer labeled by peroxidase and rabbit anti-mouse IgG for 60 min. After a wash with PBS, the sections were washed and colorized with diaminobenzidine solution (Wako Chemicals; Tokyo, Japan). Methyl green or hematoxylin was used for counterstaining of nuclei.
To avoid nonspecific binding of the antibodies to the heparin contained in mast cells, the specificity of immunostaining was checked. PBS for washing and dilution of the antibodies was acidified to pH 6.0. Preadsorption of the STG I antibody with 1000 U/ml heparin (Sigma; St Louis, MO) for 60 min at RT before use was carried out to avoid nonspecific binding of immunoglobulins and heparin. The specificity of the immunohistochemistry was confirmed with negative controls: absence of primary or secondary antibody and avidin-labeled peroxidase. Normal non-immune mouse serum was also used instead of the primary antibody. The tissues of rat brain, human brain, and pheochromocytoma that contain abundant synaptic vesicle proteins were fixed in 10% buffered formalin and simultaneously stained as positive controls. The specificity of the antibody for STG I has been confirmed by Western blotting using rat brain.
The sources and characteristics of the antibodies used are as follows: mast cell tryptase mouse, monoclonal, 1:3000, Chemicon (Temecula CA), against human lung mast cell tryptase; c-kit rabbit, polyclonal, 1 μg/ml, IBL (Fujioka, Japan), against the C-terminus of synthesized c-kit peptide (k963); synaptotagmin I mouse, monoclonal, 1:100, Wako Chemicals (Osaka, Japan), against rat brain synaptosome.
Cultured HMC-1 cells were autosmeared by a cytocentrifuge at 1000 rpm for 3 min and fixed with Bouin's solution for 1 hr. The smears were frozen and thawed. After that, immunocytochemical staining was performed the same as was done in the tissues. The cells cytocentrifuged and fixed with 10% buffered formalin easily peeled off from the glass and were not suitable for immunostaining. Therefore, the smears were fixed with Bouin's solution.
RT-PCR
Trizol, Superscript II reverse transcriptase, and Taq DNA polymerase were obtained from GIBCO BRL (Gaithersburg, MD). Total RNA was isolated from PMA-treated or nontreated cells by a modified acid-guanidine thiocyanate-phenol-chloroform method using Trizol. RNA was quantified by spectrophotometry at 260 nm. Total RNA (5 μg) was reverse-transcribed with 200 U of Superscript II reverse transcriptase in the presence of 0.5 μg of oligo-dT anchor primer and 0.5 μM of each dNTP. The mixture was incubated at 37C for 60 min. One tenth of the total reverse-transcribed single-stranded cDNA was then subjected to PCR with 25 pmol each of the forward and reverse primers, 0.2 mM of each dNTP, and 1 U of Taq DNA polymerase in a final volume of 20 μl in PCR buffer. The sense primer for rat or human STG I was 5′-GTGAGTGCCAGTCATCCTGAG-3′ or 5′-GTGAGCGAGAGTCACCATGAG-3′, and the antisense primer for rat and human STG I was 5′-CCTTCATGGTCTTCCCTAAGTC-3′. The predicted size of the amplified product was 340 base pairs (Xi et al. 1999). To confirm the integrity of the total RNA samples and RT activity, the β-actin sequence was detected by PCR on the same RT reactions with sense primer, 5′-TTCTACAATGAGCTGCGTGTGG-3′, and antisense primer, 5′-ATACCCAGGAAGGAAGGCTGGAAG-3′, for rat and human β-actin. Amplification was performed for 30 cycles with each cycle consisting of denaturation for 45 sec at 94C, 30-sec annealing at 55C, and 1-min extension at 72C. An additional 10 min at 72C was added at the end of the 30 cycles. The PCR amplification products were resolved on a 1.5% Agarose–Tris acetate–EDTA gel and visualized with ethidium bromide. DNA molecular weight markers were included on each gel to confirm PCR product sizes.
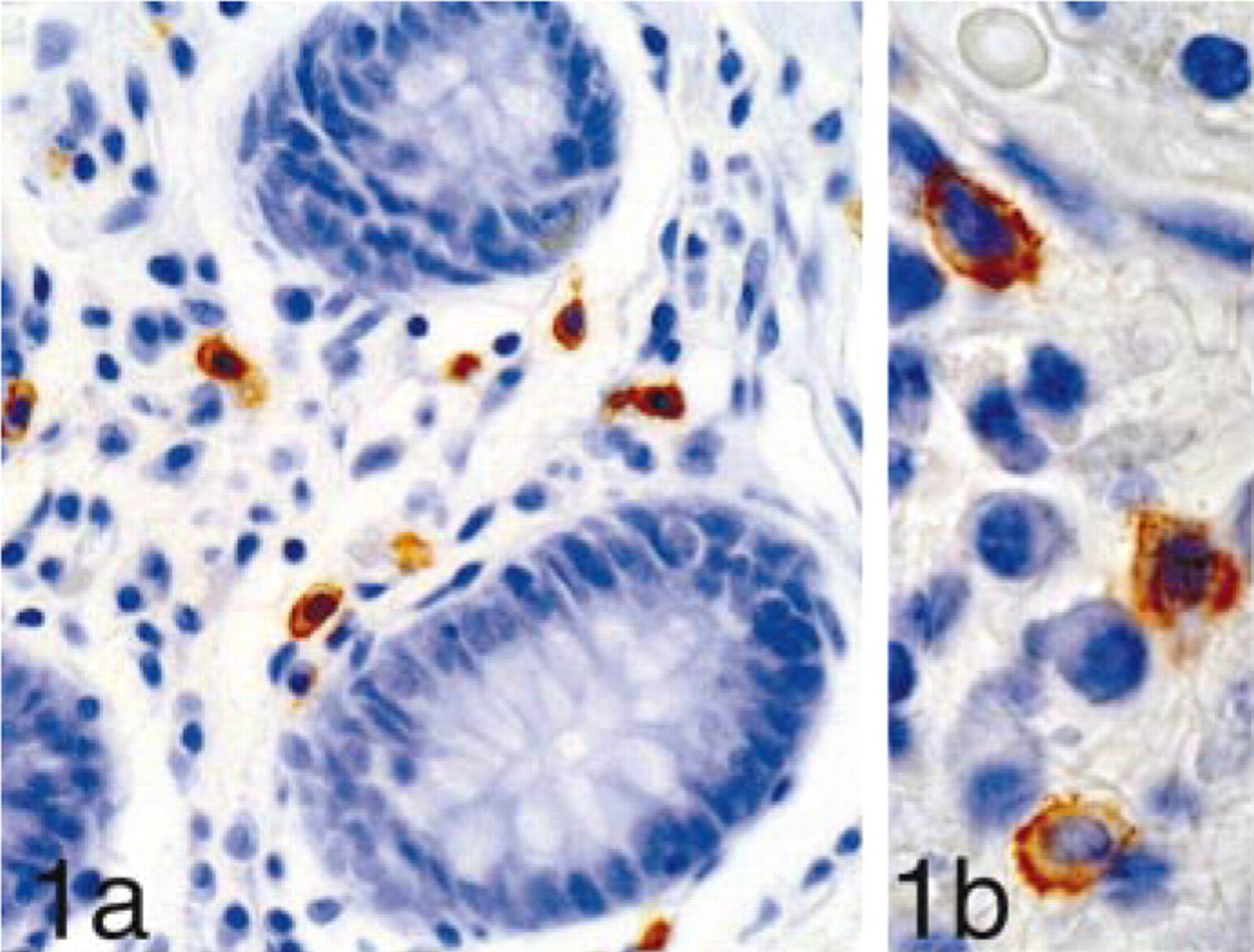
(
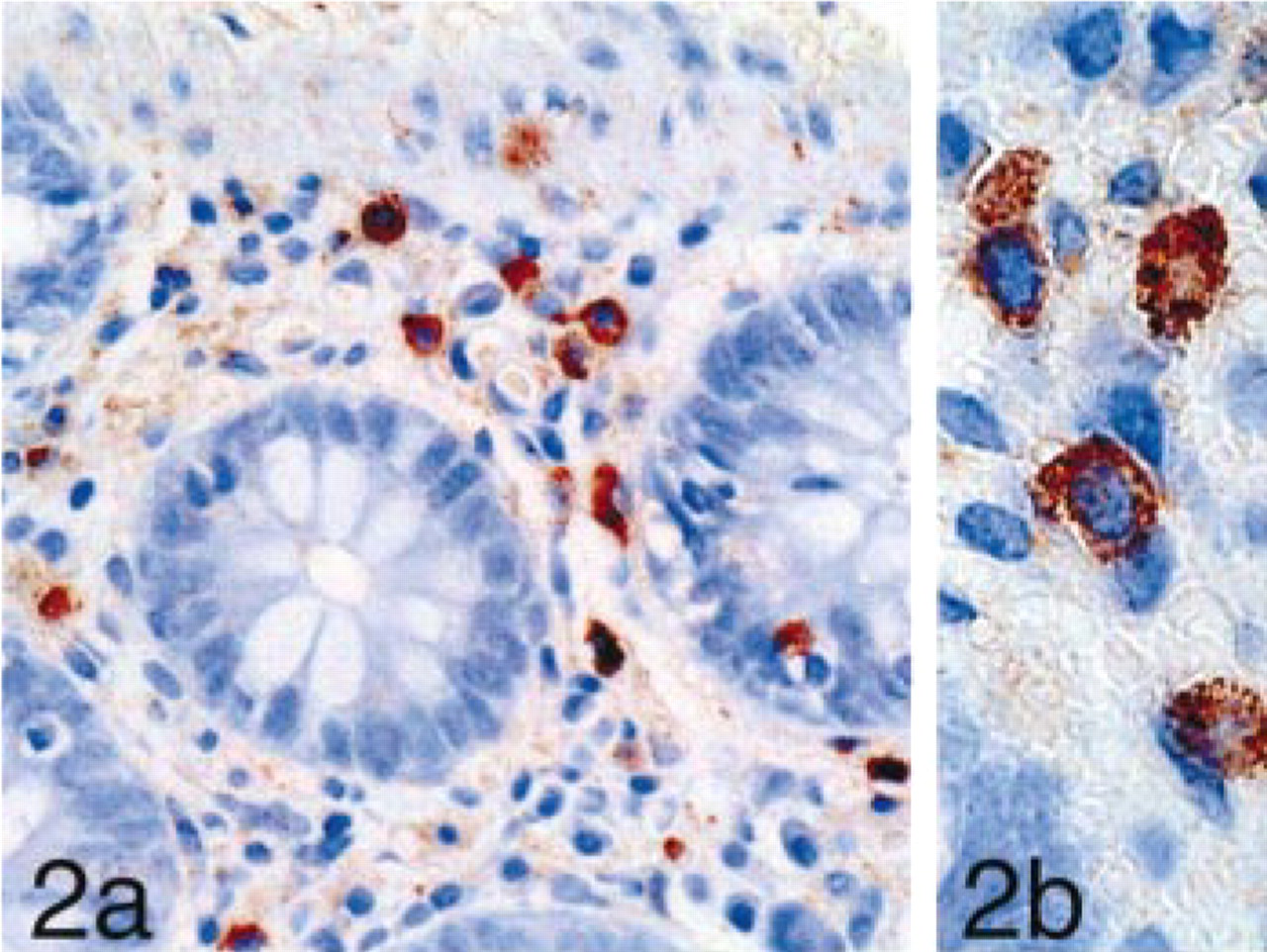
(
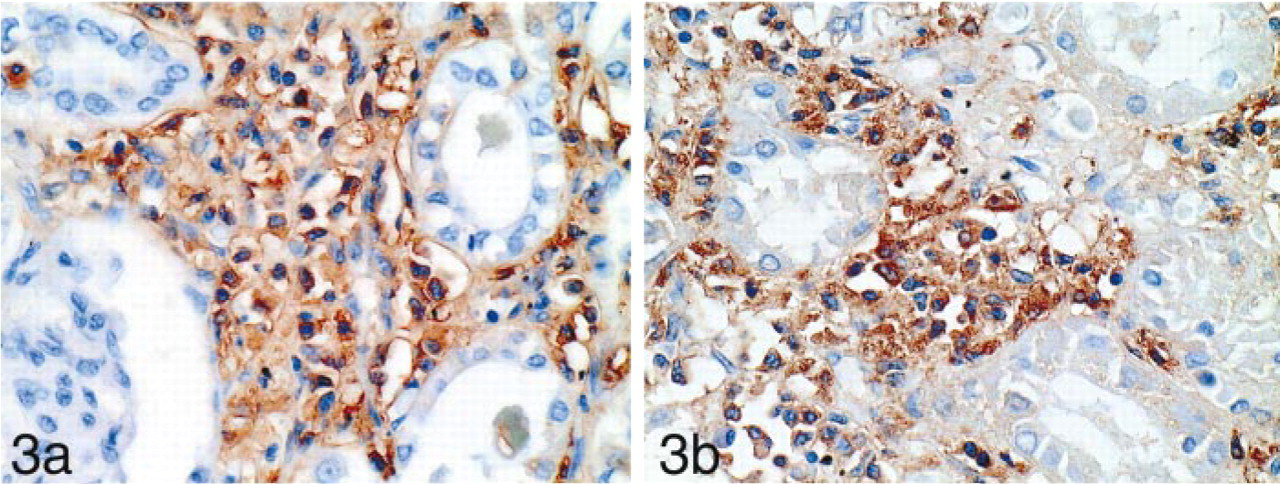
The infiltrating tumor cells of systemic mast cell disease in the stroma of the kidney are immunoreactive to the antibodies for human mast cell tryptase (
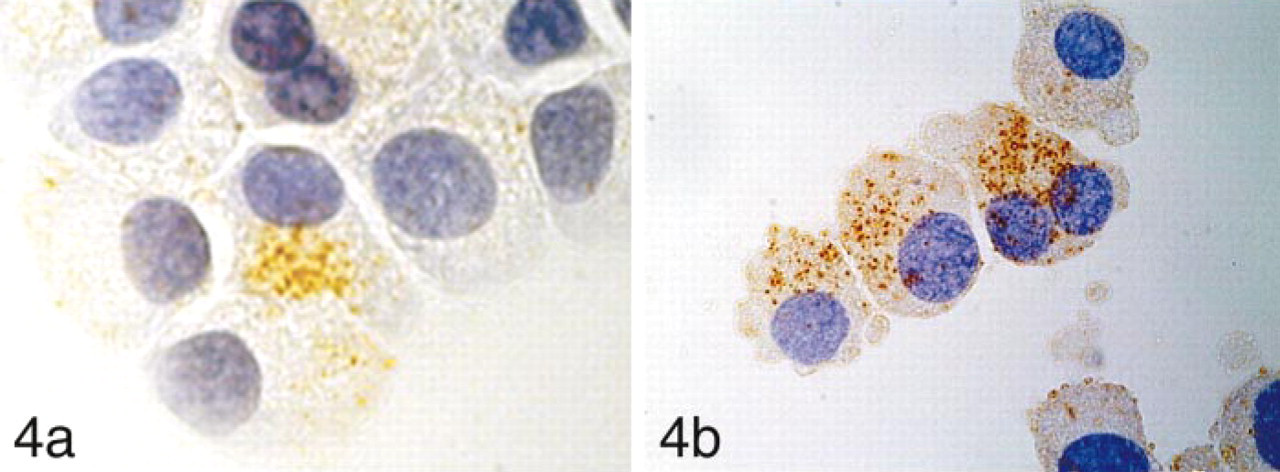
(
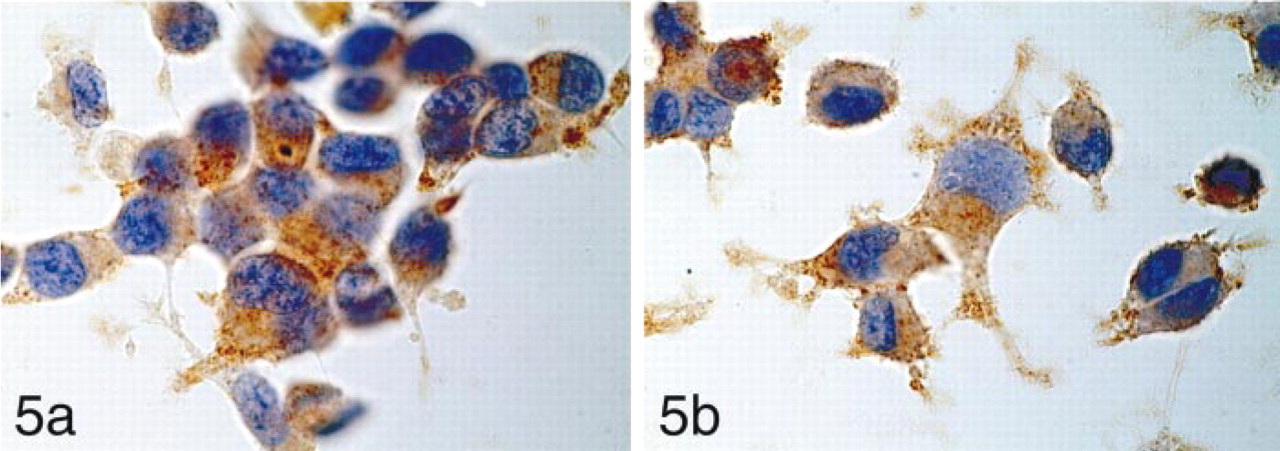
Non-stimulated NB-1 cells (
Results
Detection of Mast Cells in Normal Tissues
We detected many mast cells in all the tissues examined, showing cytoplasmic staining by mast cell tryptase and cell membrane staining by c-kit (Figures 1a and 1b).
STG I in Mast Cells of Normal Tissues
Both the SAB-PO kit and the Simple Stain kit showed specific immunoreactivity compared to the control studies. STG I immunoreactivity was observed as rough granules in mast cells locating in the mucosal tissues and connective tissues of all the specimens examined (Figures 2a and 2b).
Negative Controls
Non-immune serum, omission of the primary and the secondary antibody, and avidin complex produced no specific immunostaining.
Positive Controls for STG I
The tissues of rat brain, human brain, and pheochromocytoma stained simultaneously showed strong immunoreactivity in the cytoplasm.
STG I in Mast Cells of SMCD
Infiltrating abnormal mast cells in the stomach, lymph node, and kidney of the patient with SMCD showed immunoreactivity for tryptase (Figure 3a), c-kit, and STG I (Figure 3b).
STG I in HMC-1, PC12, and NB-1 Cells
STG I was detected as a rough granular pattern in the HMC-1 cells. The majority of non-stimulated HMC-1 cells had fewer granules immunoreactive for STG I (Figure 4a). In contrast, HMC-1 cells stimulated with PMA contained many granules immunoreactive for STG I (Figure 4b). The number of STG I-positive cytoplasmic granules increased after PMA stimulation, reaching a maximum at 5 hr after stimulation. The cells stimulated for 8 hr showed fragmented nuclei compatible with apoptotic death. NB-1 cells (Figure 5a) and PC12 cells (Figure 5b) demonstrated strong immunoreactivity in the entire cytoplasm regardless of PMA stimulation. The results are summarized in Table 1.
Synaptotagmin I expression in HMC-1 stimulated and non-stimulated NB1 and PC12 cells a

a+, weakly stained (under 1/10 intensity of NB1); ++, moderately stained (about 1/12 intensity of NB1); + + +, extensively stained (same intensity of NB1).
RT-PCR
Expression of STG I mRNA in Cultured Cell Lines. RT-PCR analysis revealed STG I mRNA expression in HMC-1 cells before and after PMA treatment as well as in PC12 cells and NB-1 cells. Both non-stimulated and stimulated HMC-1 cells expressed mRNA showing the same sized PCR product as that from NB1 and PC12 cells. (Figure 6).
Discussion
Because mast cells are well known for their nonspecific binding with immunoglobulin, we have carefully carried out immunohistochemical studies using heparin preadsorbed antibody for STG I and two different detection kits for immunohistochemistry: streptavidin–biotin complex, and an amino acid polymer-linked with IgG and peroxidase. We confirmed that both kits gave the same immunoreactivity.
STG I was detected in human mast cells in the mucosal and connective tissues of various normal tissues examined. Mast cells were immunoreactive for STG I tryptase, and c-kit. The tumor cells of SMCD also demonstrated immunoreactivity for STG I as well as tryptase and c-kit.
STG I immunoreactivity increased in HMC-1 cells stimulated by PMA. HMC-1 is an immature mast cell line and c-kit receptors are constitutively activated in these cells (Butterfield et al. 1989). In the stimulated HMC-1 cells, the rough granular immunoreactivity to STG I antibody increased remarkably. PMA is believed to induce cell maturation and secretion of the large granules via protein kinase C. It has been reported that PMA increased the expression of histidine decarboxylase in mast cells (Maeda et al. 1998).
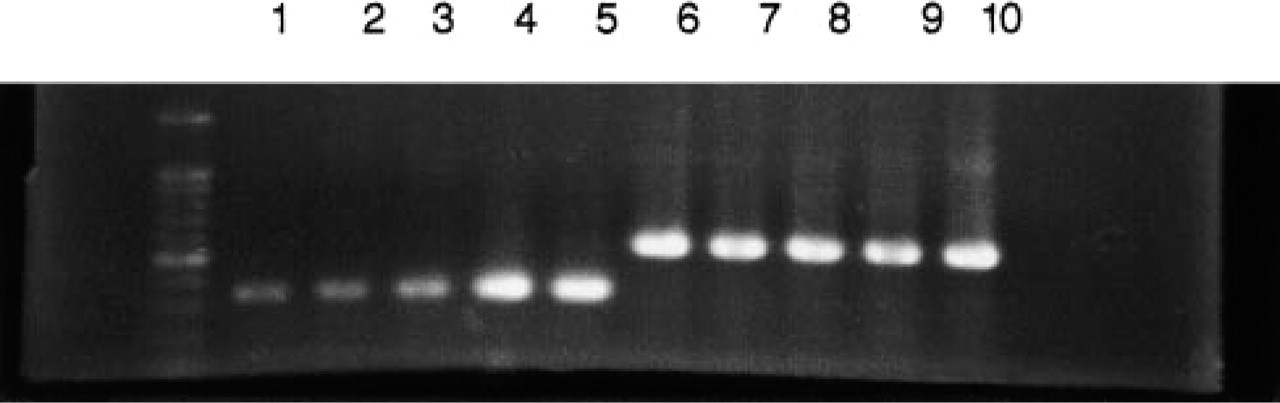
RT-PCR products of STG I and β-actin. From left to right: DNA size marker, HMC-1 non-stimulated (Lane 1), HMC-1 treated with PMA 10 ng/ml and incubated for 2 hr (Lane 2) and 5 hr (Lane 3) after treatment, NB1 (Lane 4) and PC12 (Lane 5) in order. STG I transcripts are observed in all lanes. Lanes 6–10 are β-actin in HMC-1, NB1, and PC12, the same order as above.
STG is a calcium sensor on the surface of synaptic vesicles (Brose et al. 1992). The microinjection of monoclonal and polyclonal antibodies raised against STG decreased the K+/Ca2+-mediated dopamine β-hydroxylase surface staining, which supports the hypothesis that STG is important in regulation of exocytosis in neurons (Elferink et al. 1993). STG I has been detected not only in synaptic vesicles but also in anterior and intermediate pituitary and in adrenal medullary cells (Marqueze et al. 1995). Baram et al. (1998) found that STG I mediated rat mast cell exocytosis using immunocytochemistry and Western blotting analysis. These authors also reported that STG I was localized almost exclusively to the secretory granules of the mast cells. Furthermore, the present study demonstrated the presence of mRNA of STG I in HMC-1 by RT-PCR. We therefore propose that STG I plays an important role in exocytosis in large granules of mast cells, especially mature cell types.
In conclusion, the present study confirms that STG I mRNA and STG I protein are present in human mast cells, the tumor cells of SMCD, and HMC-1 cells, especially cells stimulated by PMA. STG I may play important roles in mast cell exocytosis.
Footnotes
Acknowledgements
We thank Dr J.H. Butterfield (Mayo Medical School, Mayo Clinic, and Mayo Foundation; Rochester, MN) for supplying the HMC-1 cells and Dr H. Maruyama (Hoshigaoka Koseinenkin Hospital; Hirakata, Japan) for supplying tissues from a patient with SMCD.