
Abstract
Intercellular signaling controls all steps of odontogenesis. The purpose of this work was to immunolocalize in the developing mouse molar four molecules that play major roles during odontogenesis: BMP-2, −4, FGF-4, and WNT10b. BMP-2 and BMP-4 were detected in the epithelium and mesenchyme at the bud stage. Staining for BMP-2 markedly increased at the cap stage. The relative amount of BMP-4 strongly increased from E14 to E15. At E15, BMP-4 was detected in the internal part of the enamel knot where apoptosis was intense. In contrast to TGFβ1, BMP-2 and −4 did not show accumulation at the epithelial-mesenchymal junction where the odontoblast started differentiation. When odontoblasts became functional, BMP-2 and BMP-4 were detected at the apical and basal poles of preameloblasts. BMP-2, which induces ameloblast differentiation in vitro, may also be involved physiologically. The decrease in FGF-4 from E14 to E15 supports a possible role for the growth factor in the control of mesenchymal cell proliferation. The relative amount of FGF-4 was maximal at E17. The subsequent decrease at E19 showed correlation with the withdrawal of odontoblasts and ameloblasts from the cell cycle. WNT10b might also stimulate cell proliferation. At E14-15, WNT10b was present in the mesenchyme and epithelium except for the enamel knot, where the mitotic activity was very low. At E19 there was a decreasing gradient of staining from the cervical loop where cells divide to the tip of the cusp in the inner dental epithelium where cells become postmitotic. The target cells for FGF-4 and WNT10b appeared different.
Keywords
I
BMPs influence apoptosis, cell proliferation, and cell differentiation (von Bubnoff and Cho 2001). Several genes encoding BMPs (Bmp-2, −4, −5, −6, and −7) are expressed during tooth development (Aberg et al. 1997). BMP-2 and/or BMP-4 have been shown to mimic some of the signaling functions of the dental epithelium on mesenchyme during tooth initiation (Vainio et al. 1993; Bei and Maas 1998). Analysis of the expression patterns of different Bmps indicates that Bmp-2 and Bmp-4 show remarkable co-distribution. The corresponding proteins are involved in the mediation of epithelial-mesenchymal interactions. They are expressed in the EK and may play a role in the control of tooth morphogenesis (Aberg et al. 1997; Peterkova et al. 1998). BMP-2 and −4 have been shown to induce the differentiation of the odontoblast (Bègue-Kirn et al. 1994) and ameloblast (Coin et al. 1999a).
The FGF family in mammals is composed of at least 20 members (Ornitz and Itoh 2001). They have important regulatory functions in morphogenesis and organogenesis, as shown for limb, hair, lung, and feather. Several members of the FGF family (FGF-1, −2, −3, −4, −8, −9 and −10) have been implicated in dental morphogenesis (Kettunen and Thesleff 1998; Kettunen et al. 2000; Kratochwil et al. 2002).
Wnt genes encode a large family of secreted cysteine-rich proteins that play key roles in intercellular signaling during development (Parr and McMahon 1994; Christian 2000). WNTs have essential roles as regulators of cell proliferation, migration, differentiation, and in epithelial-mesenchymal interactions during morphogenesis. Wnt3, 4, 5a, 6, 7b, 10a, and 10b are differentially expressed in the molar from the lamina to the early bell stages (Dassule and McMahon 1998; Sarkar and Sharpe 1999).
Most data on the expression of these signaling molecules during odontogenesis was obtained by in situ hybridization. They showed temporal and spatial expression patterns consistent with paracrine signaling mechanisms (for review see Thesleff and Sharpe 1997). Because BMPs, FGFs and WNTs are diffusible molecules, the purpose of the present study was to immunolocalize BMP-2, −4, FGF-4, and WNT10b, proteins that play major roles during tooth development. The stages investigated in the first lower molar were from the bud to the late bell, during which odontoblast and ameloblast differentiations are initiated.
Materials and Methods
Immunohistochemistry
ICR day (13, 14, 15, 17, and 19) pregnant mice (vaginal plug day = day 0) were sacrificed in compliance with the recommendations of the European Economic Community (86/609/CEE) for the care and use of laboratory animals. The embryos were removed and the heads were frozen with 2-methyl butane in dry ice and stored at −20C until use. Frontal and serial (7-μm) frozen sections were fixed in cold acetone. After three washes in PBS, the slides were incubated in 3% H2O2 in methanol to quench endogenous peroxidase activity. Nonspecific binding sites were blocked with serum for 1 hr. This was followed by an overnight incubation at 4C in the presence of the primary antibody. Goat primary polyclonal antibodies directed against BMP-2 (1:250) (sc 6895), BMP-4 (1:150) (sc 6896), FGF-4 (1:400) (sc 1361), and WNT10b (1:250) (sc6546) were from Santa Cruz Biotechnology (Santa Cruz, CA). After rinsing with PBS, the sections were incubated for 30 min at room temperature in biotinylated anti-goat antibody (dilution 1:200 in PBS, 1.5% blocking serum). After rinsing in PBS, they were incubated with avidin coupled to horseradish peroxidase for 30 min, rinsed, reacted with the chromogenic substrate DAB, and counterstained with nuclear Fast Red. The ABC Staining System (sc 2023; Santa Cruz Biotechnology) was used. For negative controls the primary antibody was omitted.
Western Blotting
First lower molars from mouse embryos at E14, 15, 17, and 19 were homogenized in lysis buffer (1% NP-40, 20 mM Tris, pH 7.4, 150 mM NaCl, 10% glycerol, 1 mM sodium orthovanadate, 4 mg/ml NaF, 8.8 mg/ml sodium pyrophosphate decahydrate, 1 mM PMSF, 10 μg/ml aprotinin, and 20 μM leupeptin). After centrifugation at 16,000 × g for 10 min at 4C (Kwon et al. 1997), the protein concentration in the supernatant was determined by the Bradford method. Proteins (20 μg of each sample) were separated by SDS-PAGE (13% acrylamide) under reducing conditions and transferred to nitrocellulose for 3 hr at 36 V. Membranes were saturated with 1% BSA in Tris-buffered saline (TBS) pH 7.4, containing 0.1% Tween-20, incubated overnight with antibodies directed against BMP-2, BMP-4, FGF-4, and WNT10b (all from Santa Cruz; dilution 1:250) at 4C. After washing with TBS/Tween-20, the membranes were incubated for 1 hr at RT with a horseradish peroxidase-conjugated donkey anti-goat IgG (Sigma, St Louis, MO; dilution 1:40000), rinsed with TBS/Tween-20, and finally developed using the ECL Western Blotting Analysis System (Amersham Biosciences; Poole, UK).
For antigen quantitation (BMP-2, BMP-4, FGF-4, and WNT10b) during molar development, nitrocellulose membranes were then treated for 30 min at 60C with Tris 62.5 mM, pH 6.8, containing 2% SDS to detach antibodies. After two rinses with TBS, membranes were saturated as previously described and incubated for 2 hr with a primary antibody (Promega, Madison, WI; dilution 1:5000) which specifically recognized two isoforms (p44/ERK1 and p42/ERK2) of the Mitogen Activated Protein Kinase used as a reference (Seger and Krebs 1995). After washing, the membranes were incubated with an anti-rabbit IgG-horseradish peroxidase (Promega; dilution 1:5000) and developed as previously described. All protein bands were scanned and the density of each band was determined using the NIH Image 1.63 software (NIH; Bethesda, MD). Experiments were performed in triplicate. For each stage, the data were normalized against MAPK by calculating the protein:MAPK density ratio.
Results
For each antigen, the intensity of the staining observed in the molar was compared with reference tissues on the same section. These were the olfactory epithelium for BMP-2 and WNT10b (Figures 1B and 4B) and the eye for BMP-4 (Figure 2B).
BMP-2 and BMP-4
At the bud stage (E13), immunostaining for BMP-2 was more intense in the condensing mesenchyme than in the dental epithelium (Figures 1A and 1C). At this stage, BMP-4 was detected in both the epithelium and mesenchyme with a similar intensity (Figures 2A and 2D). The control was negative (Figure 2C). At the cap stage (E14 and E15), antibodies to BMP-2 mainly reacted with the mesenchyme (Figures 1D and 1E). BMP-4 was present in the epithelium and mesenchyme at E14 (Figure 2E) and the staining decreased in both tissues at ED15 (Figure 2F). At E14, the primary EK was negative for BMP-2 (Figure 1D). Furthermore, BMP-4 was detected in primary EK cells distant from the basement membrane at E15 (Figure 2F). From E15 to the bell stage (E17), BMP-2 was mainly detected in the mesenchyme (Figure 1F). During the same period, immunostaining for BMP-4 increased in the pulp (compare Figures 2F and 2G). The secondary EKs were negative with both antibodies (data not shown). Finally, at the late bell stage (E19), staining for BMP-2 (Figure 1G) and for BMP-4 (Figures 2H and 2K) decreased in the pulp, becoming more pronounced at the basal and apical poles of the inner dental epithelium (IDE) cells and in the stratum intermedium (SI) for BMP-2 (Figures 1I and 1J). The control slides were negative (Figure 1H).
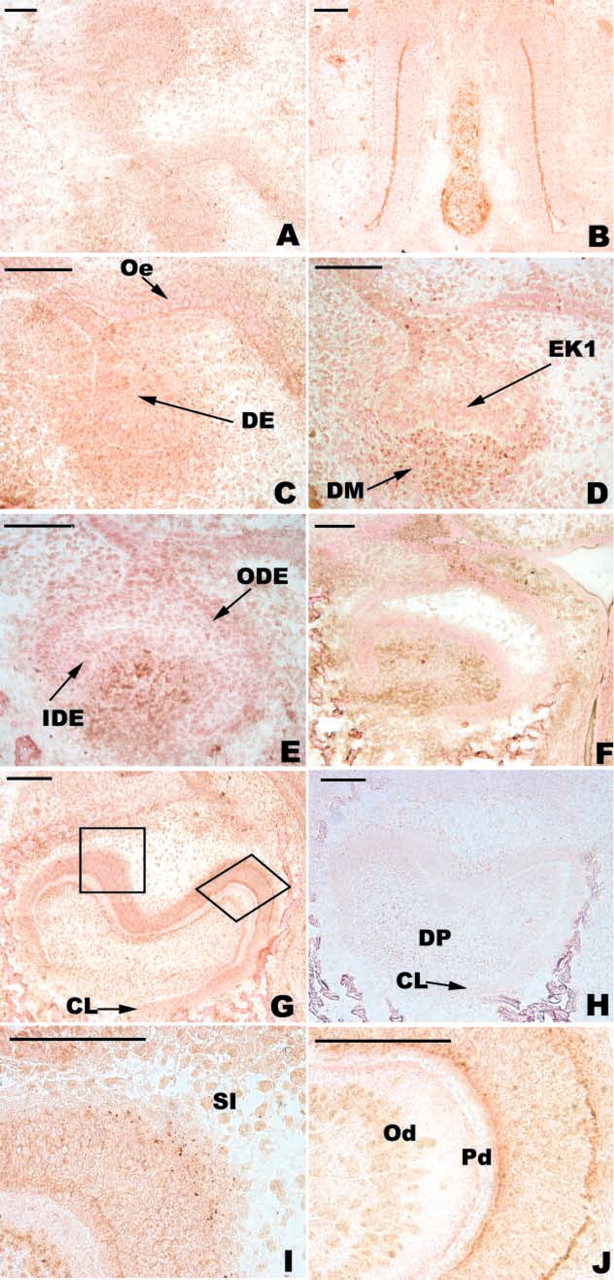
Immunolocalization of BMP-2 in the developing first lower mouse molar. At the bud stage (E13), BMP-2 was detected in the epithelium and mesenchyme (
Immunolocalization of BMP-4 in the developing first lower mouse molar. At the bud stage (E13), BMP-4 was detected in the epithelium and mesenchyme (
In the molar, only one protein band of 16 kD was detected with the antibody directed against BMP-2 (Figure 5). Compared to MAPK, which was used as a reference, the amount of BMP-2 was maximal at E14 and then decreased until E17 and remained almost constant at E19 (Table 1). Molars from E13 could not be assessed because of significant peridental mesenchyme contamination.
Two bands reactive with the anti-BMP4 antibody were detected in the molars at 21-22 kD and at 16 kD, but the signal was weak (Figure 5). The relative amount of BMP-4 increased from E14 to E19 (Table 1). BMP-4 was maximal at E19 (Table 1).
FGF-4
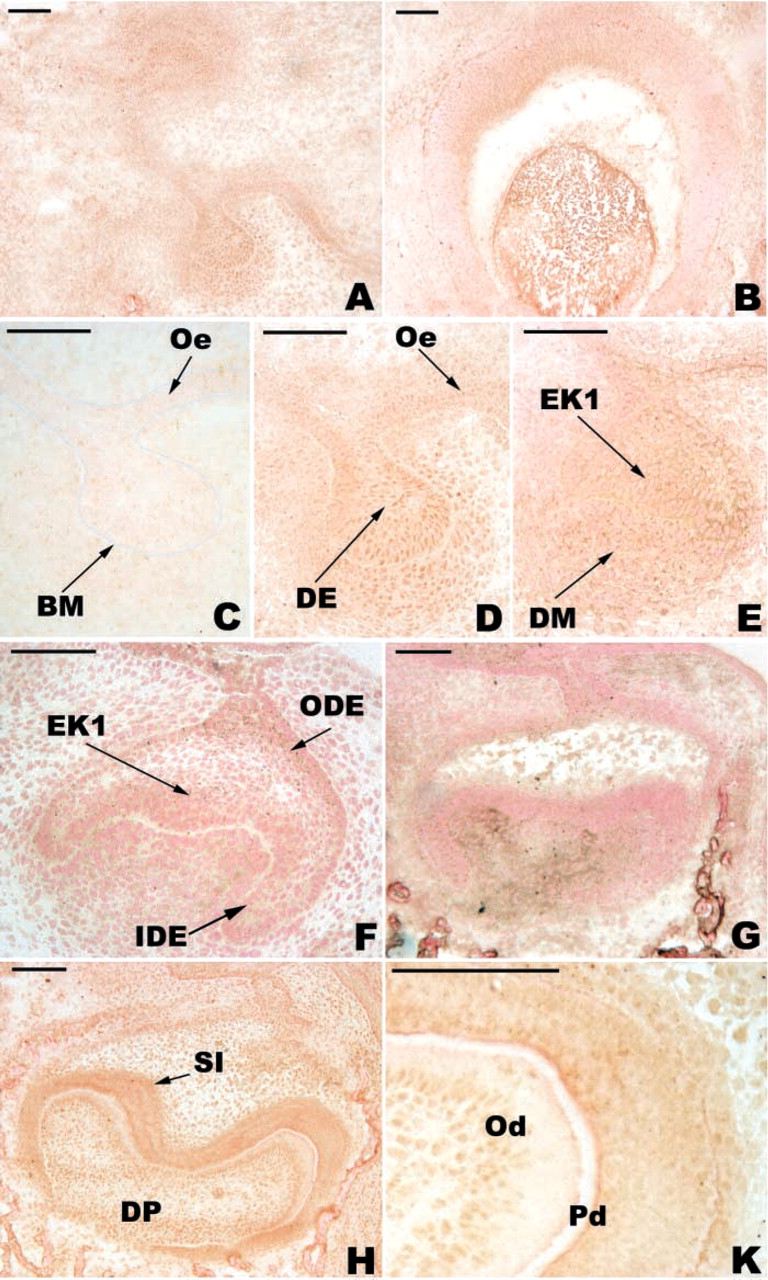
At E13, FGF-4 was localized in the epithelium and mesenchyme (Figure 3A). From the bud to the bell stage, FGF-4 was associated with the basement membrane (Figures 3A-3C) except for the basement membrane underlying cells of the primary EK at E14 and E15 (Figures 3B and 3C). At the cap stage (E14 and E15), the staining for FGF-4 was stronger in the enamel organ than in the mesenchyme (Figures 3B and 3C). At E17, the staining was very strong in the IDE and the SI (Figure 3D). In the mesenchyme, weak staining for FGF-4 was detected at the tips of cusps (Figure 3D). At the late bell stage (E19), the labeling significantly decreased (Figure 3E) except for the SI (Figure 3F).
Density ratio of antigens vs MAPK during molar development a
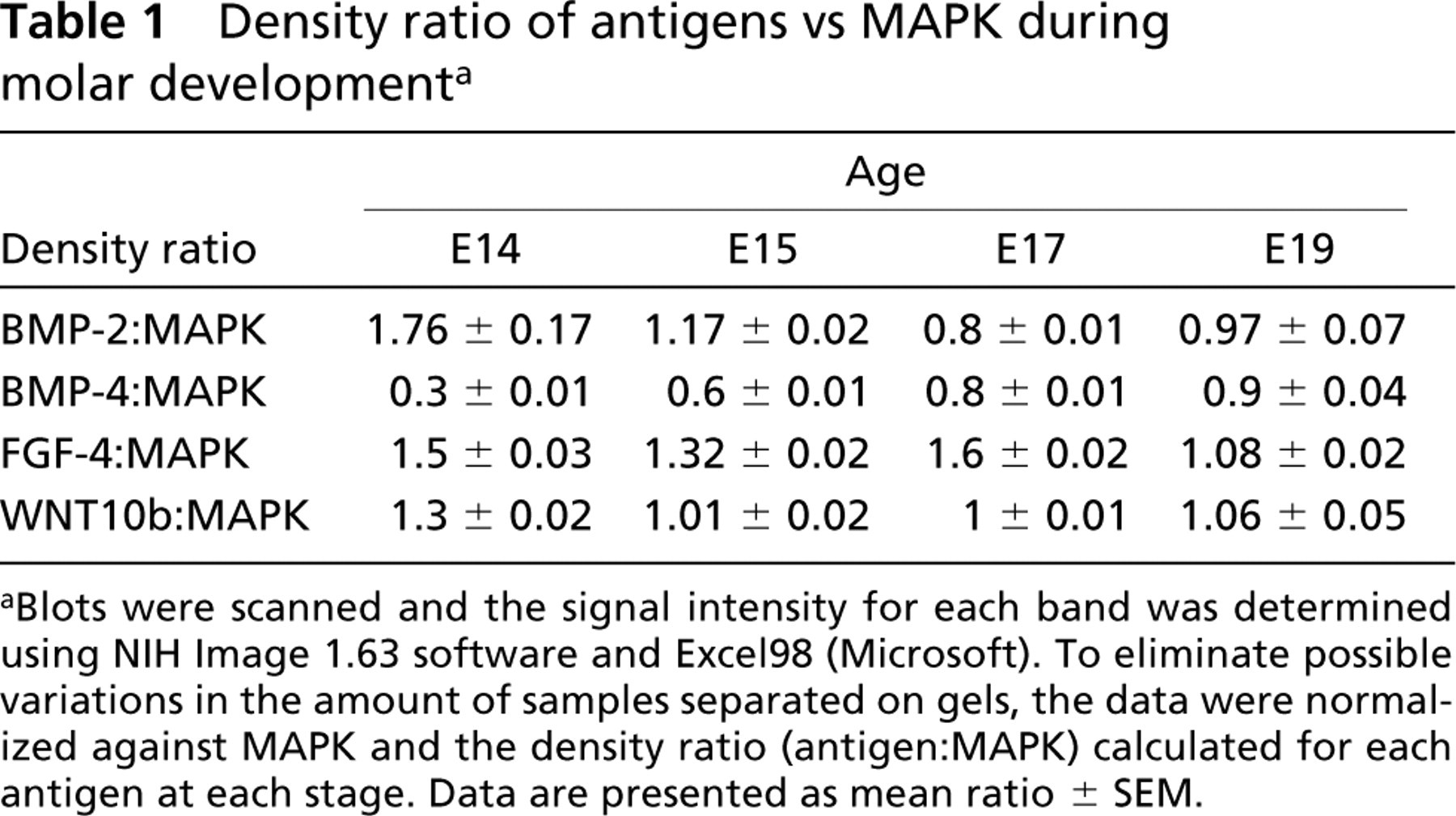
aBlots were scanned and the signal intensity for each band was determined using NIH Image 1.63 software and Excel98 (Microsoft). To eliminate possible variations in the amount of samples separated on gels, the data were normalized against MAPK and the density ratio (antigen:MAPK) calculated for each antigen at each stage. Data are presented as mean ratio ± SEM.
After immunoblotting, a single band was detected at 14-15 kD with anti-FGF-4 antibody (Figure 5). Expression of FGF-4 was maximal at E17 (Table 1).
WNT10b
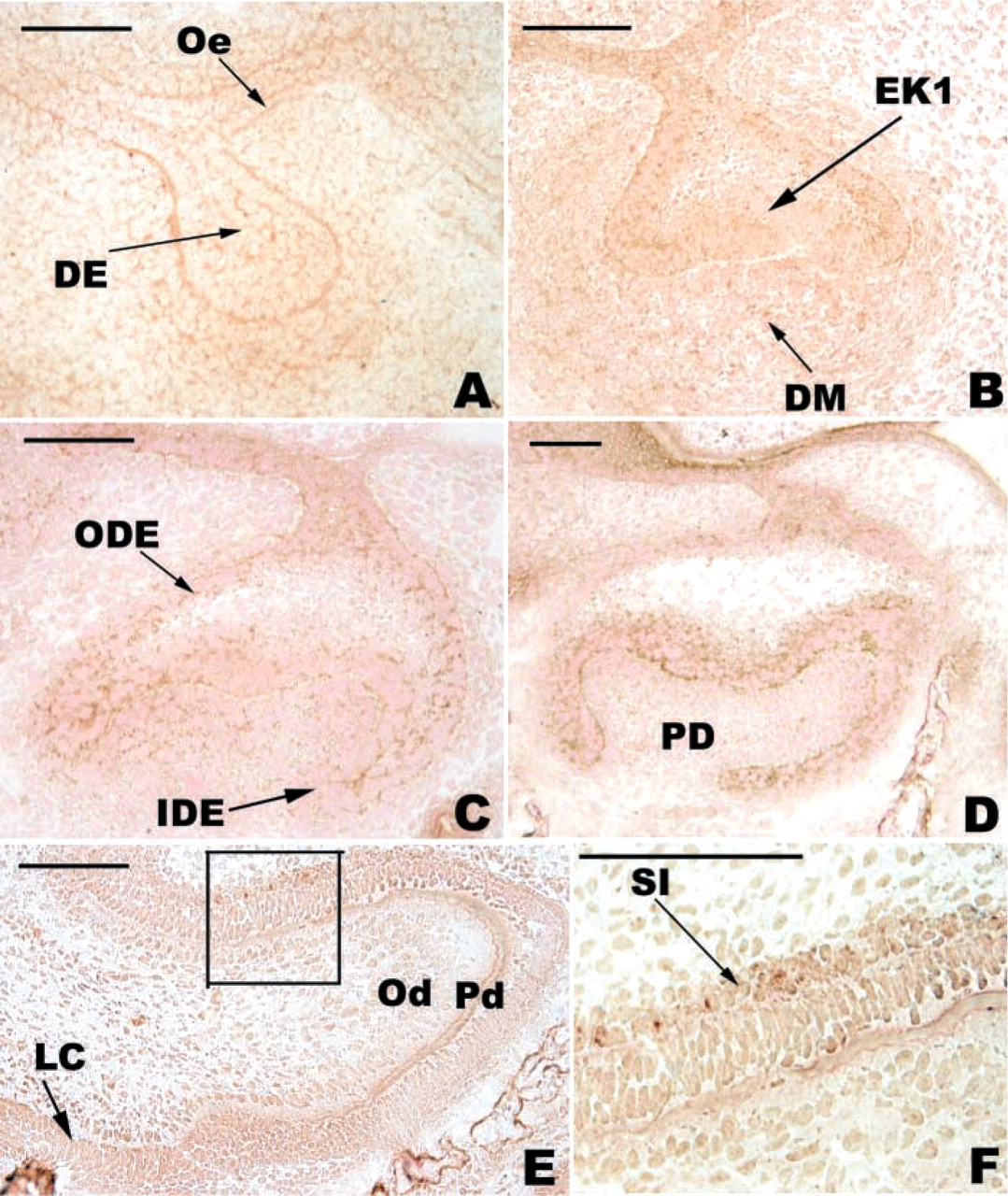
At E13, positive immunoreactivity with the anti-WNT10b antibody was observed in both the dental epithelium, mainly at the tip of the bud, and the condensing mesenchyme (Figure 4C). At the cap stage (E14-E15), the staining in the enamel organ was maintained but showed a buccal-lingual asymmetry: the staining was more intense on the buccal aspect (Figures 4A, 4D, and 4E). During this period, the cells of the primary EK remained negative (Figures 4D and 4E). From E13 to E15, the staining in the mesenchyme decreased (compare Figures 4C, 4D, and 4E). From E14 to E17, the staining of the cervical loop increased (Figures 4D-4F). At E17, the secondary EKs were negative (Figure 4F). At E19, WNT10b was detected in the preameloblasts and in SI cells (Figure 4G). At this stage, WNT10b staining disappeared from IDE cells at the tip of the main cusp except for the basal pole of these cells (Figure 4G).
The anti-WNT10b antibody reacted with a protein band of 28-29 kD (Figure 5). The relative amount of WNT10b was maximal at E14 (Table 1).
Discussion
Bud to Cap Transition
The bud to cap transition is characterized by formation of the cervical loop and histogenesis of the dental epithelium when the IDE and outer dental epithelium (ODE) are separated by the stellate reticulum. During this period the EK extends in the central part of the molar. In most cases, attempts to interfere with signaling molecules involved in odontogenesis have resulted in the arrest of tooth development at the bud-cap transition, as observed with mice deficient for several genes controlled by BMPs, FGFs, or WNTs (Peters and Balling 1999; Jernvall and Thesleff 2000).
The Bmp-2 gene is intensely expressed in the dental epithelium at the tip of the bud (Aberg et al. 1997). However, the corresponding protein was detected in the epithelium as well as in the mesenchyme, where the staining was more pronounced. This discrepancy between the in situ hybridization and immunostaining patterns is due to the ability of BMP-2 to diffuse. At the bud stage, however, the staining still remained very weak in the developing tooth compared to that observed in the cartilage or the olfactory epithelium. BMP-2 modulates chondrogenic differentiation (Zhang et al. 2002). BMP-2 is also involved in the morphogenesis of the olfactory system (Fishell 1999; Peretto et al. 2002). In the developing tooth, the immunostaining for BMP-2 markedly increased in the mesenchyme at the cap stage.
Immunolocalization of FGF-4 in the developing first lower mouse molar. At E13, the basement membrane (BM) was intensely stained (
At the bud stage, Bmp-4 was expressed in the condensing mesenchyme (Aberg et al. 1997). Similarly to BMP-2, BMP-4 was immunolocalized both in the epithelium and mesenchyme at the bud stage. According to Hall and Miyake (1995), BMP-4 might regulate mesenchymal condensation. At the cap stage, the Bmp-4 gene was strongly expressed in the dental pulp and in the distal part of the primary EK (Jernvall et al. 1998; Keranen et al. 1998). The corresponding protein was initially present in the dental epithelium and mesenchyme. BMP-4 from the mesenchyme may also regulate epithelial morphogenesis at the cap stage (Jernvall et al. 1998). At E15, the staining for BMP-4 decreased in the mesenchyme but persisted in the enamel organ. Strong staining for BMP-4 was observed in the inner part of the primary EK, where apoptosis is intense (Viriot et al. 1997; Lesot et al. 1999). BMP-2 and −4 are known to play a role in the regulation of apoptosis during development (Jernvall et al. 1998; Peterkova et al. 1998; Guha et al. 2002). The relative amounts of BMP-4 strongly increased from E14 to E15, which is in agreement with the immuno-histochemical findings and with the simultaneous increase in apoptosis within the primary EK. Furthermore, the differential effects of BMPs in the epithelium (apoptosis) and mesenchyme (condensation) suggest that different signaling pathways are stimulated in each tissue.
At E13, FGF-4 was detected in the epithelium and mesenchyme and strong staining was observed at the epithelial-mesenchymal junction. FGF-4 has a high affinity for heparan sulfate (Aviezer et al. 1999). Heparan sulfate, a component of the dental basement membrane, can modulate FGF-4 receptor recognition and regulate FGF-4 signaling activity (Aviezer et al. 1999; Allen et al. 2001). A specific CD44 splice variant binds both FGF-4 and FGF-8 and is involved in limb outgrowth (Sherman et al. 1998). Several heparan sulfate proteoglycans have been detected in the tooth, including syndecan isoforms (Bai et al. 1994) and the membrane-associated CD44 (Yu and Toole 1997). At E14, Fgf-4 was expressed only in the primary EK (Kettunen and Thesleff 1998). As a result of diffusion, the localization of the protein was again different from that of the corresponding transcripts observed by in situ hybridization. FGF-4 was present mainly in the dental epithelium and the staining was more intense in the ODE than in the IDE. FGF-4 stimulates cell proliferation in both the epithelium and mesenchyme and has been suggested to inhibit apoptosis in the dental mesenchyme (Vaahtokari et al. 1996). Only very limited apoptosis can be detected in the mesenchyme at E14 and E15 (Viriot et al. 1997). The relative amount of FGF-4 decreased from E14 to E15. During this period the mitotic index remained unchanged in the IDE-SI while it decreased in the mesenchyme from the central part of the tooth (Lesot et al. 1999). After E15, the weak staining for FGF-4 in the mesenchyme progressively disappeared when tooth development progressed.
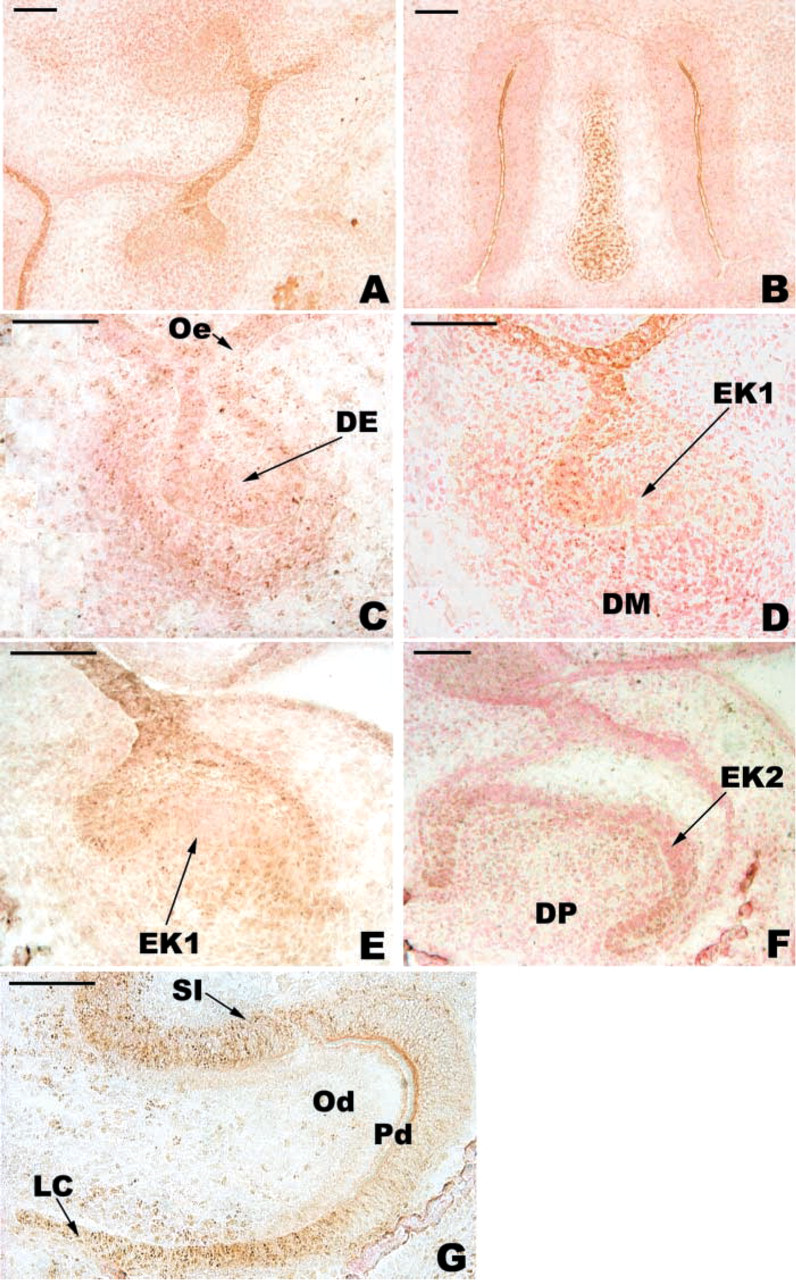
Immunolocalization of WNT10b in the developing first lower mouse molar. At E13, WNT10b was detected in the dental epithelium (DE) and in the condensing mesenchyme (
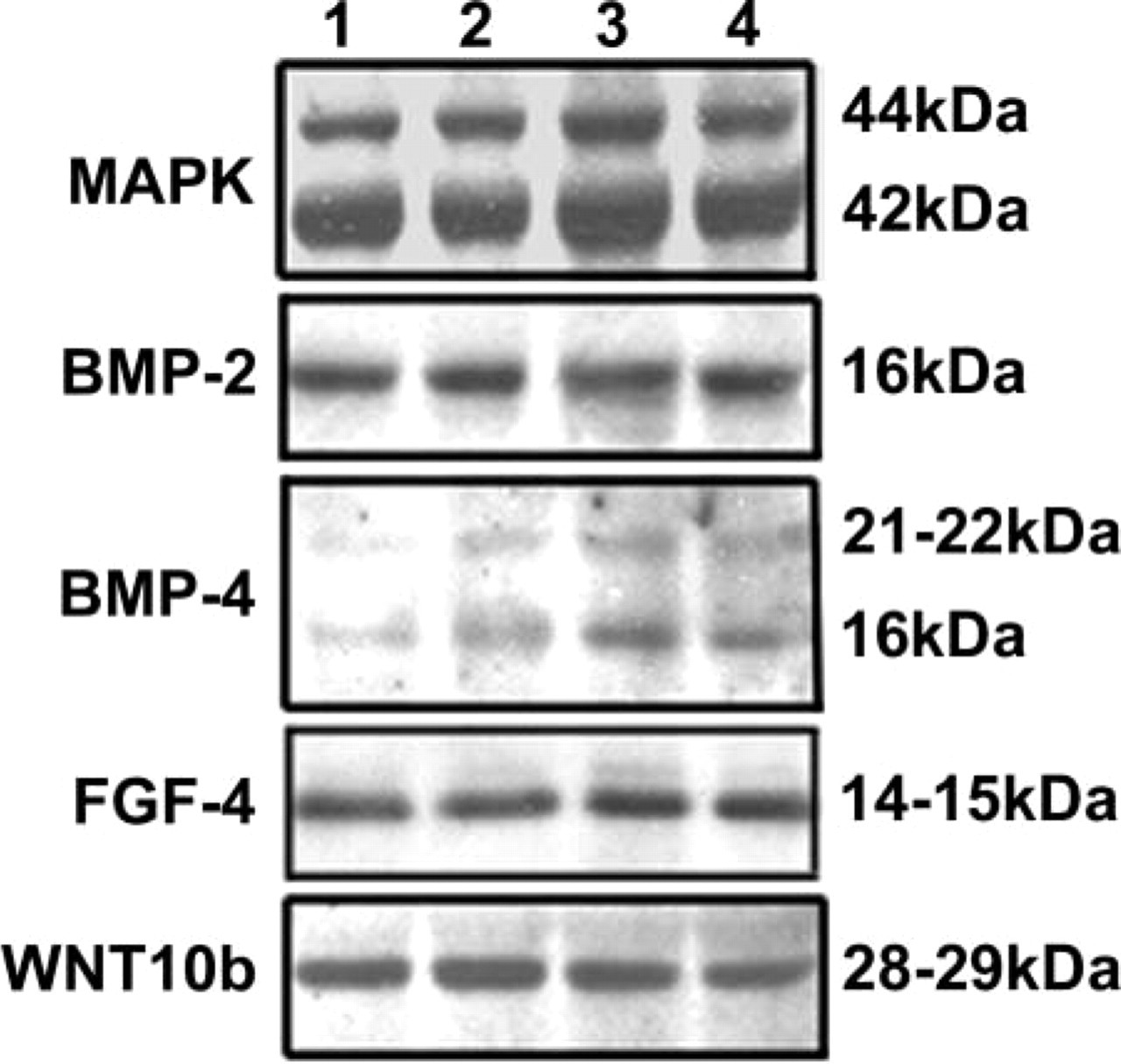
Western blot analysis of BMP-2, BMP-4, FGF-4, and WNT10b expression in the developing first lower mouse molar at E14 (Lane 1), E15 (Lane 2), E17 (Lane 3), and E19 (Lane 4). Twenty μg of proteins extracted from tooth germ at each stage was separated by 13% SDS-PAGE. MAPK was used as an internal reference protein.
At the bud stage, Wnt10b is expressed at the tip of the epithelial bud (Dassule and McMahon 1998), while WNT10b was detected in the epithelium and in the condensing mesenchyme. At the cap stage, Wnt10b was expressed in the primary EK (Dassule and McMahon 1998) and throughout the IDE (Sarkar and Sharpe 1999). The corresponding protein at E14 was present in the dental and peridental mesenchyme and also in the enamel organ except for the primary EK. The relative amount of WNT10b decreased from E14 to E15. At both the bud and cap stages, the locations of Wnt10b transcripts were different from that of the corresponding protein. Depending on the context, Wnts can act either as local inducers (Zecca et al. 1996) or as long-range morphogens to control tissue organization and growth (Zecca et al. 1996; Neumann and Cohen 1997). In the developing tooth, WNT10b showed significant diffusion and would therefore act as a long-range morphogen. Heparan sulfate has been proposed to be involved in the extracellular distribution of WNT (Baeg et al. 2001).
Cap to Bell Stage
During this transition from E15 to E17, the cervical loop elongates to surround the dental mesenchyme. The segregation of the non-dividing or slowly dividing IDE cells of the EK leads to the formation of the secondary EKs (Coin et al. 1999b; Lisi et al. 2003). The epithelial-mesenchymal junction changes its shape when cusps develop.
At E14, the expression of Bmp-2 became restricted to the primary EK (Keranen et al. 1998). Bmp-2 has been suggested to play a role in the determination of the secondary EKs (Aberg et al. 1997). Indeed, BMP-2 was mainly detected in the dental mesenchyme at E14. From E15 to E17, the staining for BMP-2 did not show any major change and remained positive in the mesenchyme only. The cap stage mesenchyme determines tooth morphogenesis (Kollar and Baird 1969), probably by controlling the pattern of segregation of the BrdU-negative cells from the primary EK (Schmitt et al. 1999). At E15-E17, Bmp-4 was strongly expressed in the pulp, including the preodontoblasts, when the cusps formed (Aberg et al. 1997). From E15 to E17, the corresponding protein showed an increased staining in the mesenchyme. The analysis of E13.5 molars cultured in the presence of antisense oligodeoxynucleotides suggested that BMP-4 was necessary for cusp formation at the late bud and cap stage (Tabata et al. 2002).
At E15, Fgf-4 was expressed only in the primary EK and later in the secondary EKs (Kettunen and Thesleff 1998; Kettunen et al. 1998). From E15 to E17, the immunostaining for FGF-4 strongly increased in the enamel organ. Conversely, there was a decrease in the staining of the mesenchyme except for the cusp tips. This localization was in accord with the possible role of FGF-4 in tooth growth and cusp formation (Jernvall and Thesleff 2000). During the cap to bell stage transition, the size of the molar significantly increased. The increase in the surface covered by the IDE resulted from both a histological reorganization of the IDE controlled by Shh (Lesot et al. 2002) and cell proliferation in the IDE, which could be stimulated by FGF-4.
The staining for WNT10b at E15 was strong in the cervical loop and negative in the primary EK, where the mitotic activity is very low. At E17, WNT10b was present in the mesenchyme and the enamel organ. From E15 to E17, the staining for WNT10b became negative in the ODE. The staining in the enamel organ decreased from E15 to E17 except for the cervical loop area. This staining pattern was maintained at E19, supporting the potential role for WNT10b in the growth of the cervical loop.
Late Bell Stage and Cytodifferentiation
At E18, the first odontoblasts differentiate at the main cusp tips and their differentiation progresses towards the basal region of each cusp with a specific time-space pattern, different from the pattern of cusp formation (Lisi et al. 2003). The terminal differentiation of ameloblasts follows 24 hr later.
Bmp-2 is expressed by odontoblasts and preameloblasts when predentin secretion has started (Bègue-Kirn et al. 1994; Aberg et al. 1997). Similarly, Bmp-4 is expressed by odontoblasts (Bègue-Kirn et al. 1994) and in the IDE, where it appears to persist longer than Bmp-2 (Aberg et al. 1997). Using in vitro functional approaches, it has been shown that TGFβ1 and −3, BMP-2, and BMP-4 can induce odontoblast differentiation (Bègue-Kirn et al. 1992; Ruch 1998; Unda et al. 2000). The relative amount of BMP-2 and, to a lesser extent, BMP-4 increased from E17 to E19. Immuno-staining showed that TGFβ1 accumulated at the epithelial-mesenchymal junction where odontoblasts differentiated (Cam et al. 1997; Kikuchi et al. 2001). However, immunostaining for BMP-2 and −4 did not suggest any storage of these proteins at the epithelial-mesenchymal junction in regions where odontoblasts had started to differentiate. This is in agreement with the fact that TGFβs could be extracted from dentin fractions, whereas there is no evidence for the presence of BMP-2 and −4 in dentin (Smith and Lesot 2001). However, BMP-2 and BMP-4 were detected at the basal pole of preameloblasts at the tip of the main cusp at E19, just before they differentiated. BMP-2 has been shown to induce ameloblast differentiation in vitro (Coin et al. 1999a). The molecule may therefore be involved in ameloblast differentiation in vivo.
From E17 to E19, the relative amount of FGF-4 decreased by 31% and the immunostaining for FGF-4 was very weak in all dental tissues at E19. The decrease in FGF-4 at E19 might therefore correspond to the withdrawal from the cell cycle for more and more odontoblasts and for the first ameloblasts.
At the late bell stage, there was weak staining for WNT10b in the mesenchyme except for the tip of the main cusp, which was negative. WNT10b was mainly detected in the IDE. There was an intense staining of preameloblasts and in the cervical loop area. The staining decreased towards the tip of the main cusp. Both FGF-4 and WNT10b appear to be involved in cell proliferation. However, the staining for WNT10b persisted for a longer time in the enamel organ. It can be expected that the target cells for FGF-4 and WNT10b are different. To test this hypothesis and extend this work, study of the receptors is in progress. Functional experiments will be necessary to analyze the signaling effects of these molecules after the cap stage, for which little information is available.
Footnotes
Acknowledgements
Amal Nadiri was financed by the IFRO.
We wish to thank Prof A.J. Smith for critical reading of the manuscript.