
Abstract
Experiments were performed to investigate whether adrenal neuronal nitric oxide synthase (nNOS) mRNA and protein expression are responsive to alterations in body volume. Using an RT-PCR technique, the relative quantities of nNOS mRNA as well as the tyrosine hydroxylase and phenylethanolamine N-methyltransferase mRNA in the adrenals of water-deprived rats significantly increased from 12 hr to 4 days. In situ hybridization and immunohistochemical study showed that water deprivation activated nNOS mRNA and protein expression in the adrenal medulla. Four days after water deprivation, nNOS protein expression determined by Western blot significantly increased in the adrenal gland. Our results are the first to demonstrate that nNOS syntheses in the adrenal medulla are markedly increased in water-deprived rats. This study also indicates that the upregulation of nNOS synthesis of the adrenal medulla is associated with the activation of adrenal medullary function in the face of volume depletion.
N
Materials and Methods
Animal Experiments
Male Wistar rats, 8–10 weeks of age and weighing between 290 and 360 g, purchased from the Experimental Animal Center, National Science Council, Taiwan, were housed individually in metabolic cages for 3 days before the start of the experiments. The rats were divided into two groups: water deprivation and normal control. Complete water deprivation was done on normal rats for 0.5, 1, 2, or 4 days (n=5, 5, 5, and 10, respectively). Body weight- and age-matched rats for 0.5, 1, 2, or 4 days (n = 5, 5, 5, and 10, respectively) were given tap water to drink and were included as normal controls. All rats were fed normal rat laboratory chow. Body weight was recorded at the beginning and at the end of the experiments. Twenty-four-hour urine samples were collected continuously during the entire study period for catecholamine determination. The rats were sacrificed at the end of the experiment by decapitation to collect blood and tissues. Truncal blood was collected in prechilled tubes containing EDTA (1 mg/ml) and aprotinin (500 kIU/ml), and plasma was stored at −20C until assay. The right adrenal of each animal was frozen and stored at −70C for RNA extraction and Western blot. The left adrenal was bisected and fixed in 4% paraformaldehyde for more than 10 hr, then paraffin-embedded, following standard procedures. Animal studies were approved by the Animal Care and Treatment Committee of our institution.
Plasma and Urine Samples
Atrial natriuretic peptide (ANP) in plasma samples was determined using a specific RIA kit (Peninsula Laboratories, Inc.; Belmont, CA) after extraction as in our previous reports (Lee et al. 1993,1996). Plasma renin activity was measured by enzymatic incubation of plasma at 37C and pH 6.4. The angiotensin I generated during the incubation step was quantified using an RIA kit (DuPont-NEN; Boston, MA). The concentrations of plasma sodium were determined in an automatic analyzer (Nova Biochemical; Newton, MA). Plasma aldosterone was measured by radioimmunoassay with the use of a commercial kit (Diagnostic Products Corporation; Los Angeles, CA). Urinary catecholamine levels were determined by means of high-performance liquid chromatography (HPLC, BioRad, Model 1330 pump, Model 1340 electrochemical detector; Hercules, CA) with electrochemical detection.
Isolation of Total RNA and Reverse Transcription
Total RNA was isolated from the right adrenal gland by a commercially available kit (Trizol reagent, Gibco; Burlington, ON, Canada). Two micrograms of total RNA from the right adrenals were reverse transcribed as in our previous reports (Lee et al. 1993,1996).
Multiplex PCR with “Primer Dropping”
To minimize problems inherent in the quantification of mRNA with RT-PCR, primer-dropping multiplex PCR with an internal standard was employed (Wong et al. 1994). Sequences for rat nNOS, TH, phenylethanolamine N-methyl-transferase (PNMT), StAR, and β-actin were obtained from GenBank and used to design the primer pairs (Table 1). Images were captured using a digital SPOT RT camera (Diagnostic Instruments Inc.; Sterling Heights, MI) and SPOT RT software (Diagnostic Instruments Inc.). The amount of mRNA product for nNOS, TH, PNMT, and StAR was analyzed by ONE-Dscan 1.33 softwave (Scanalytics, Inc.; Fairfax, VA) and was expressed as the ratio to β-actin mRNA product.
In Situ Hybridization
In situ hybridization was carried out as reported previously (Lai et al. 2000,2002) with the use of digoxigenin-labeled antisense or sense nNOS cRNA probes. Digoxigenin-labeled RNA hybrids were detected by an enzyme-linked immunoassay kit (Roche Molecular Biochemical; Mannheim, Germany). After immersion in 1.5% blocking solution, the slides were exposed to an anti-digoxigenin alkaline phosphatase conjugate diluted at 1:1000 for 30 min. The hybrids were visualized as purple/black precipitates by subsequent alkaline phosphatase-chloro-3-indolyl phosphate and nitroblue tetrazolium.
Immunohistochemistry
Immunohistochemistry was carried out with the use of a rabbit polyclonal antiserum against nNOS (Calbiochem-Novabiochem Corporation; San Diego, CA) as the primary antiserum (Lee et al. 1993; Shin et al. 2000). This was followed by sequential 10-min incubations with biotinylated link antibody and peroxidase-labeled streptavidin [Dako, LSAB 2 kit (Dako Corp.; Carpinteria, CA)]. Staining was completed after incubation with DAB substrate-chromogen solution (Vector Laboratories, Inc.; Burlingame, CA), then counterstained with hematoxylin and mounted in mounting medium.
Western Blot Analysis
Western blot analysis of adrenal glands from five representative normal control and five representative water-deprived rats on the fourth day after water deprivation was performed as described by Roczniak et al. (1998) with the use of a rabbit polyclonal antiserum against nNOS or monoclonal anti-β-actin monoclonal antibody (Sigma; St Louis, MO). This was followed by incubation with horseradish peroxidase-conjugated goat anti-rabbit IgG (Jackson ImmunoResearch; West Grove, PA). Specific antibody-antigen complex was detected with the use of an enhanced chemiluminescence Western blot detection system (NEN Life Science Products; Boston, MA). The amount of nNOS protein was quantified by Photo-Print Plus software (ETS Vilber-Lourmat; Marne LuVallee Cedex, France) and was expressed as the ratio to β-actin protein.
PCR primer name, sequence, and product size

Numbers indicate the location of the nucleotide.
Data for normal control and water-deprived rats on day 4 of the study
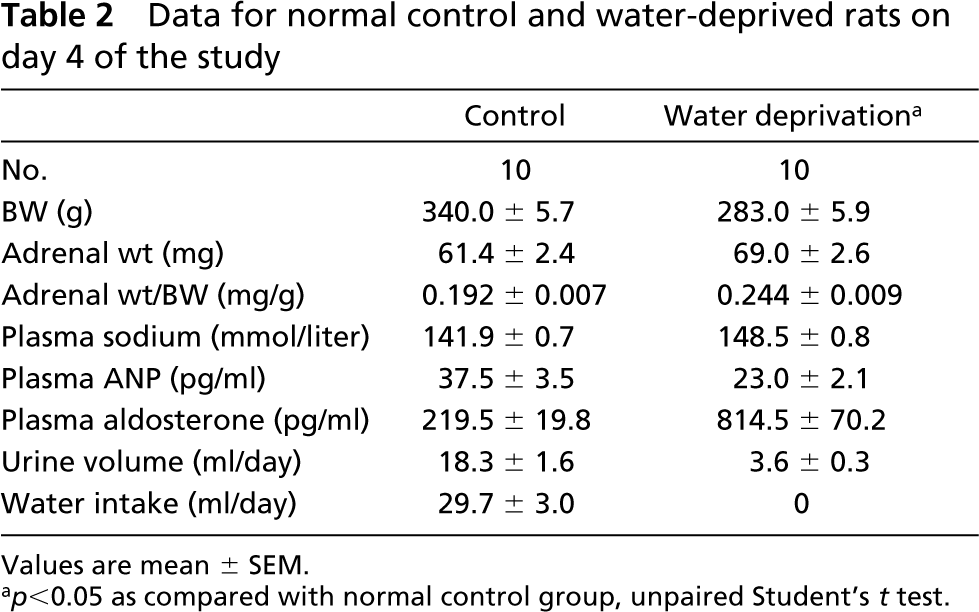
p<0.05 as compared with normal control group, unpaired Student's t test.
Statistical Analysis
The data are expressed as the mean ± SEM. To test the difference between the normal control and water-deprived rats, unpaired Student's t-test was performed. Urinary epinephrine and norepinephrine data obtained before water deprivation were compared with experimental data for the same rats after the water deprivation by ANOVA and Dunnett t-tests for multiple comparisons where appropriate. Statistical significance was accepted at a value of p<0.05.
Results
The laboratory data of 10 normal control and 10 water-deprived rat groups on the fourth study day are shown in Table 2. The mean amount of daily water intake in the normal control group was 29.7 ± 3.0 ml, whereas complete water deprivation was performed in the water-deprived rats. The mean body weight of water-deprived rats was significantly lower than that of the normal control group. The mean adrenal weight value of water-deprived rats was significantly increased as compared to that of normal control animals. The mean plasma sodium levels and plasma aldosterone concentrations also significantly increased, whereas plasma ANP concentrations were lower in the water-deprived group after 4 days of water deprivation. The mean urine volume value was significantly lower in the water-deprived group than in the normal control group. Mean 24-hr urinary epinephrine and norepinephrine excretion rates for rats before and after water deprivation are shown in Table 3. After water deprivation, the 24-hr epinephrine and norepinephrine excretion rates from days 1 to 4 were significantly higher. The plasma osmolality and plasma renin activity of normal and water-deprived rats on days 0.5, 1, 2, and 4 are shown in Figure 1. In water-deprived groups, the plasma osmolality and plasma renin activity from days 1 to 4 were significantly higher.
Water Deprivation Increases Adrenal nNOS, TH, and PNMT mRNA
To determine the relative changes in tissue nNOS, TH, PNMT and StAR mRNA expression, the yields of nNOS, TH, PNMT, and StAR cDNA PCR products were normalized to the amount of β-actin cDNA amplified from the same tube. Figure 2A shows the results of multiplex RT-PCR amplification of nNOS and β-actin in the adrenals from three representative normal control rats and five representative water-deprived rats on day 4. The relative ratios of the cDNA product for nNOS with those for β-actin in the adrenals are shown in the lower panel of Figure 2A. Significantly increased nNOS mRNA levels were observed in the adrenals of water-deprived rats from 12 hr to 4 days. Figure 2B shows the results of the PCR amplification of TH and β-actin in the adrenals from the five representative normal control rats and the five representative water-deprived rats on day 1. The relative ratios of the cDNA product for TH with those for β-actin in the adrenals are shown in the lower panel of Figure 2B. Significantly increased TH mRNA levels were observed in the adrenals of water-deprived rats from 12 hr to 4 days. Figure 2C shows the results of the PCR amplification of PNMT and β-actin in the adrenals from five representative normal control rats and five representative water-deprived rats on day 0.5. The relative ratios of the cDNA product for PNMT with those for β-actin in the adrenals are shown in the lower panel of Figure 2C. Significantly increased PNMT mRNA levels were observed in the adrenals of water-deprived rats from 12 hr to 4 days. Figure 2D shows the results of the PCR amplification of StAR and β-actin in the adrenals from the five representative normal control rats and the five representative water-deprived rats on day 0.5. The relative ratios of the cDNA product for StAR with those for β-actin in the adrenals are shown in the lower panel of Figure 2D. No significant difference in StAR mRNA levels was observed in the adrenals of water-deprived rats from 12 hr to 4 days.
Mean 24-hr urinary epinephrine and norepinephrine excretion rates for rats (n = 5) before and after water deprivation
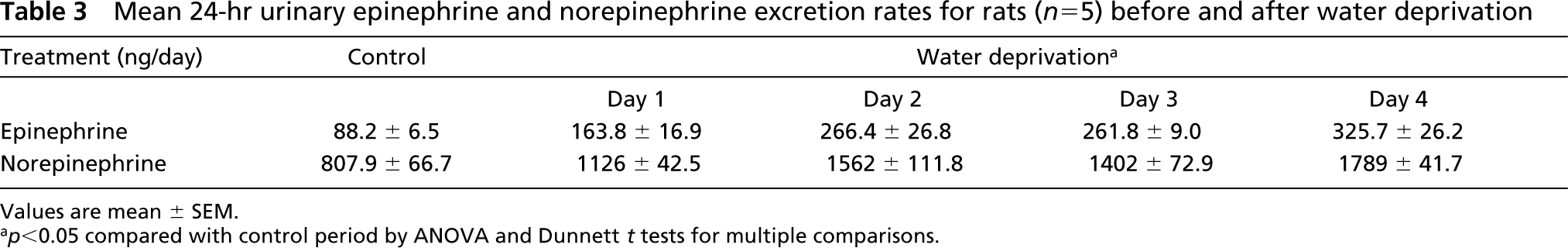
Values are mean ± SEM.
<0.05 compared with control period by ANOVA and Dunnett t tests for multiple comparisons.
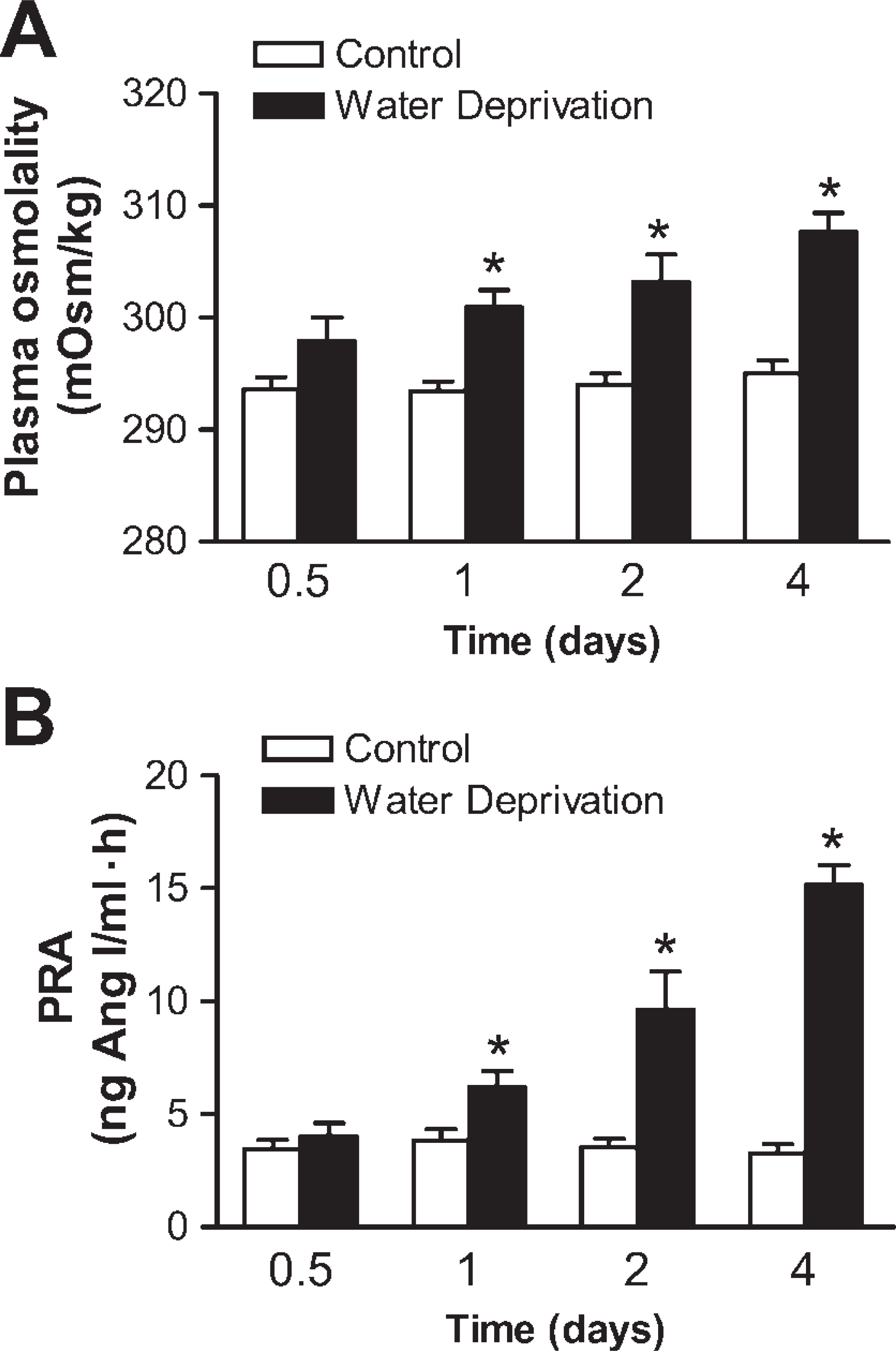
Plasma osmolality and plasma renin activity between normal control and water-deprived groups on days 0.5, 1, 2, and 4. Values (mean ± SEM) from five normal control and five water-deprived rats on days 0.5, 1, and 2, and from 10 normal control and 10 water-deprived rats on day 4. ∗p<0.05 (compared with normal control group, by Student's t-test).
Water Deprivation Enhances Adrenal nNOS mRNA Distribution
We determined directly how nNOS mRNA was distributed through in situ hybridization of adrenal sections from normal control (Figures 3A-3C) and water-deprived (Figures 3E-3G) rats on day 4 using a specific cRNA probe for nNOS. In normal control rats, the signal of adrenal nNOS mRNA was detected mainly in the medulla and also in the zona glomerulosa. In water-deprived rats on day 4, the density of nNOS mRNA hybridization increased in the medulla relative to that in normal controls. Positive staining was found in the cytoplasm and nucleus of parenchymal cells (Figures 3C and 3G). We detected no signal using a sense cRNA probe (Figures 3D and 3H).
Water Deprivation Enhances Adrenal nNOS Immunoreactive Distribution
Immunohistochemical localization of the nNOS in the adrenal gland was determined in both groups (Figure 4). The zonal distribution of adrenal nNOS immunostaining was well in accordance with that of adrenal nNOS mRNA by in situ hybridization. In normal control rats, nNOS staining was detected in the adrenal medulla and slightly in the zona glomerulosa (Figures 4A-4C). After 4 days of water deprivation, the adrenal nNOS immunostaining was enhanced in the medulla (Figures 4E-4G). Positive staining was restricted to the cytoplasm of parenchymal cells as fine granules. No immunoreactive labeling was observed when the sections were incubated with preabsorbed antibodies (Figures 4D and 4H).
Water Deprivation Increases Adrenal nNOS
The effect of water deprivation on the expression of nNOS protein in the adrenal gland was determined using Western blot. Four days after water deprivation, the nNOS protein expression was significantly higher in the adrenal gland (Figure 5A). The relative ratios of the protein for nNOS with those for β-actin in the adrenals are also shown in Figure 5B.
Discussion
The present study demonstrates that nNOS mRNA and protein expression was strongly activated in the adrenal medulla of water-deprived rats in association with the activation of adrenal medullary function. This conclusion is based on the following findings: (a) nNOS mRNA and nNOS levels were significantly enhanced in the adrenal glands of water-deprived rats in conjunction with the activation of the intra-adrenal TH mRNA and PNMT mRNA after water deprivation from study day 0.5 to day 4; (b) the intensity of staining for nNOS mRNA in the cells of adrenal medulla was markedly increased in water-deprived rats; (c) the immunoreactive staining for nNOS in adrenal medulla was also enhanced; and (d) plasma renin activity, angiotensin II, aldosterone, and urine 24-hr epinephrine and norepinephrine excretion rate were significantly higher in water-deprived rats.
Kishimoto et al. (1996) could only observe negligible levels of both nNOS mRNA and protein in the adrenal cortex of normal rats. However, Cymeryng et al. (2002) reported that nNOS mRNA and protein in the adrenal cortex were detectable in basal conditions. This preliminary study demonstrates that intensity of nNOS mRNA was predominantly hybridized in adrenal medulla and zona glomerulosa by use of a sensitive in situ hybridization technique, but not in zona fasiculata or reticularis in normal control rats. By using paraffin sections with microwave treatment instead of frozen sections without microwave heating, nNOS immunoreactive staining was detectable in adrenal medulla and zona glomerulosa, but not in zona fasiculata or reticularis in normal rats. Apparently, the distribution of nNOS immunoreactivity was well paralleled with that of adrenal nNOS mRNA. This similar distribution of nNOS immunoreactivity and nNOS mRNA reflects the possibility that nNOS substance is translated from nNOS mRNA itself in these adrenal cortical and medullary cells.
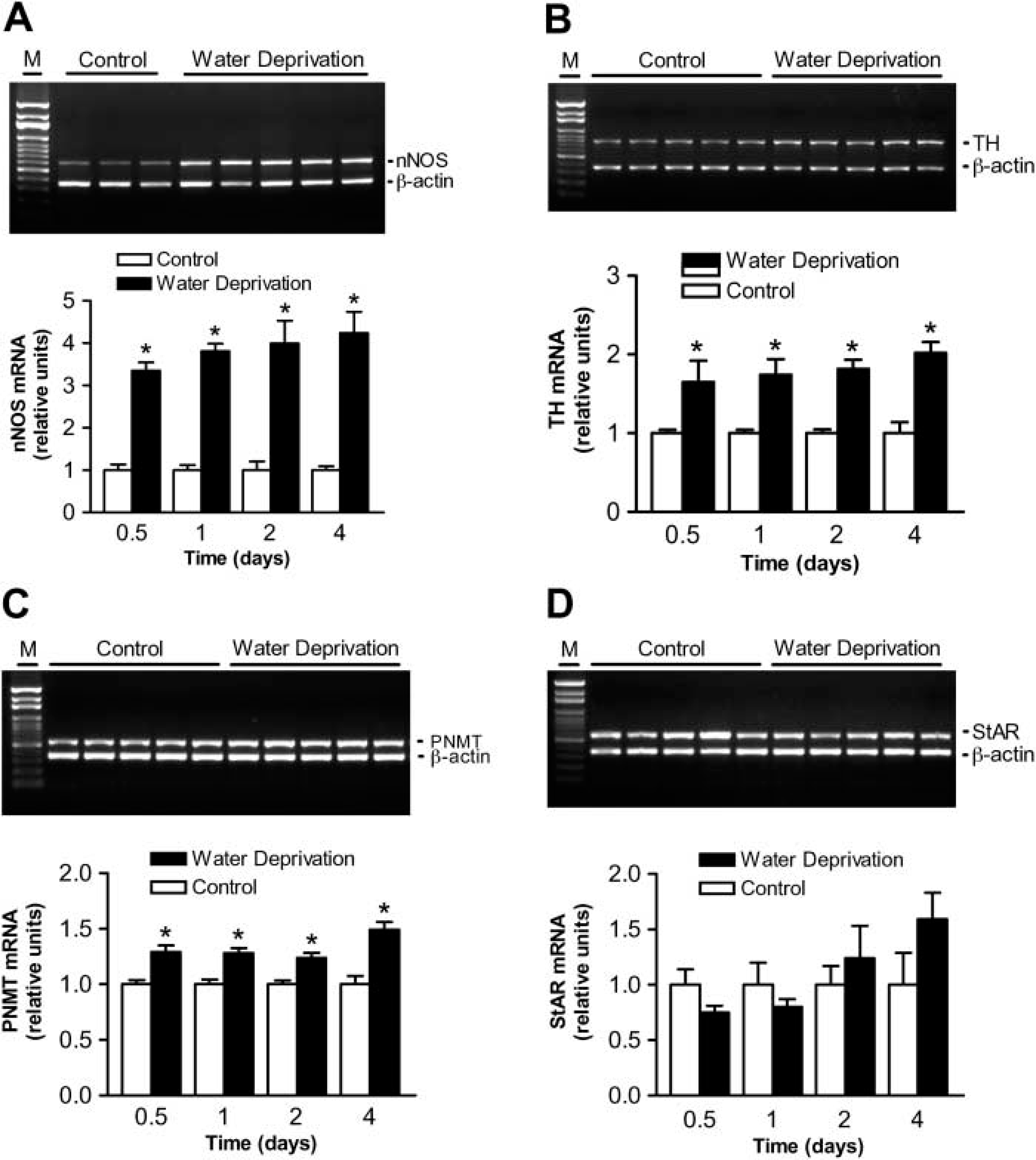
Upregulation of adrenal nNOS, TH, and PNMT mRNA expression but no significant change of StAR mRNA expression after water deprivation. (
Although the adrenal glands were found to be able to synthesize nNOS, very few studies have investigated the change of adrenal-derived nNOS in pathophysiological conditions. In 1995, Iwai et al. showed that nNOS mRNA expression of the decapsular portion of adrenal glands in spontaneously hypertensive rats (SHR) significantly decreased compared to that in Wistar-Kyoto rats. The administration of angiotensin II antagonist, hydralazine, or reserpine enhanced nNOS mRNA expression in both SHR and Wistar-Kyoto rats (Iwai et al. 1995). Therefore, these authors concluded that the reduced expression of nNOS in adrenal glands of SHR possibly resulted from hypertension. In contrast to findings in SHR, immobilization stress by wrapping with flexible wire gauze markedly increased the expression of nNOS in adrenal cortex of rats (Kishimoto et al. 1996; Tsuchiya et al. 1996). Water deprivation decreases the extracellular and intracellular volume and leads to hyperosmolality and hypernatrenemia in adult rats. Therefore, it is a good experimental model for volume depletion. In the current study, water deprivation markedly increased nNOS mRNA and nNOS levels detected by RT-PCR and Western blot analysis, respectively, in adrenal glands. Moreover, by using sensitive in situ hybridization and immunohistochemistry, we further found that the intensity of nNOS mRNA signal and nNOS immunoreactive staining was strongly increased in adrenal medullary cells in water-deprived rats. In contrast to adrenal medulla, the intensity of nNOS mRNA and immunoreactivity exhibited no apparent alteration in the adrenal cortex of water-deprived rats as compared with normal control rats. Additionally, water deprivation also significantly enhanced TH and PNMT mRNA expression in the adrenal glands and increased the 24-hr urinary epinephrine and norepinephrine excretion rates. These findings show that adrenal medullary function is strongly activated by water deprivation. To our knowledge, this study first indicates that nNOS synthesis in the adrenal medulla is strongly activated and that this activation is in conjunction with the increase of adrenal medullary function in the face of severe volume depletion.
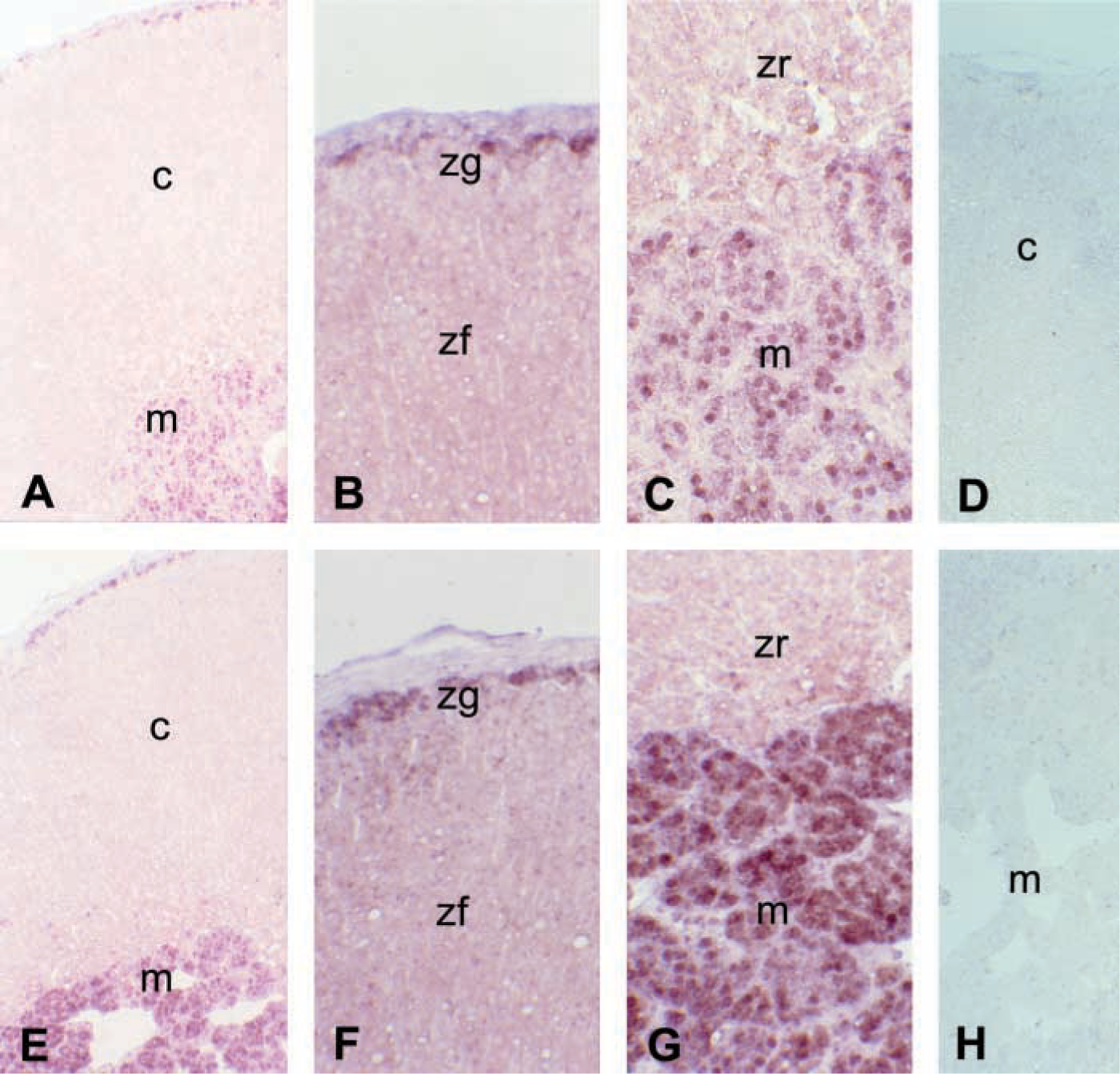
Representative photomicrographs show in situ hybridization of nNOS mRNA in the adrenal gland of 4-day control (
Water deprivation results in changes in the hormone secretions regulating the fluid and electrolyte balance. These changes maintain normal tissue perfusion in central nervous and cardiovascular systems in volume depletion (Yared et al. 1985; Gardiner and Bennett 1985; Chen et al. 1988; Thornton and Proppe 1988; Chatelain et al. 2003). In this current study, water deprivation increases plasma renin activity, angiotensin II, and aldosterone but decreases the plasma concentrations of ANP. Moreover, we also showed that water deprivation significantly increased the 24-hr urinary epinephrine and norepinephrine excretion rates and significantly enhanced adrenal TH and PNMT mRNA from days 1 to 4. These results are consistent with other studies (Gardiner and Bennett 1985; Brooks et al. 1997; Chatelain et al. 2003). However, Blair et al. (1997) found no significant change in plasma epinephrine or norepinephrine concentrations in male Sprague-Dawley rats after 2 days of water deprivation. The inconsistency for catecholamine concentration may be caused by different methodological measurement, species, and periods of water deprivation. Considered together, most of above investigations indicate that volume depletion concomitantly activated the renin-angiotensin-aldosterone system, sympathetic activity, and adrenomedullary function.
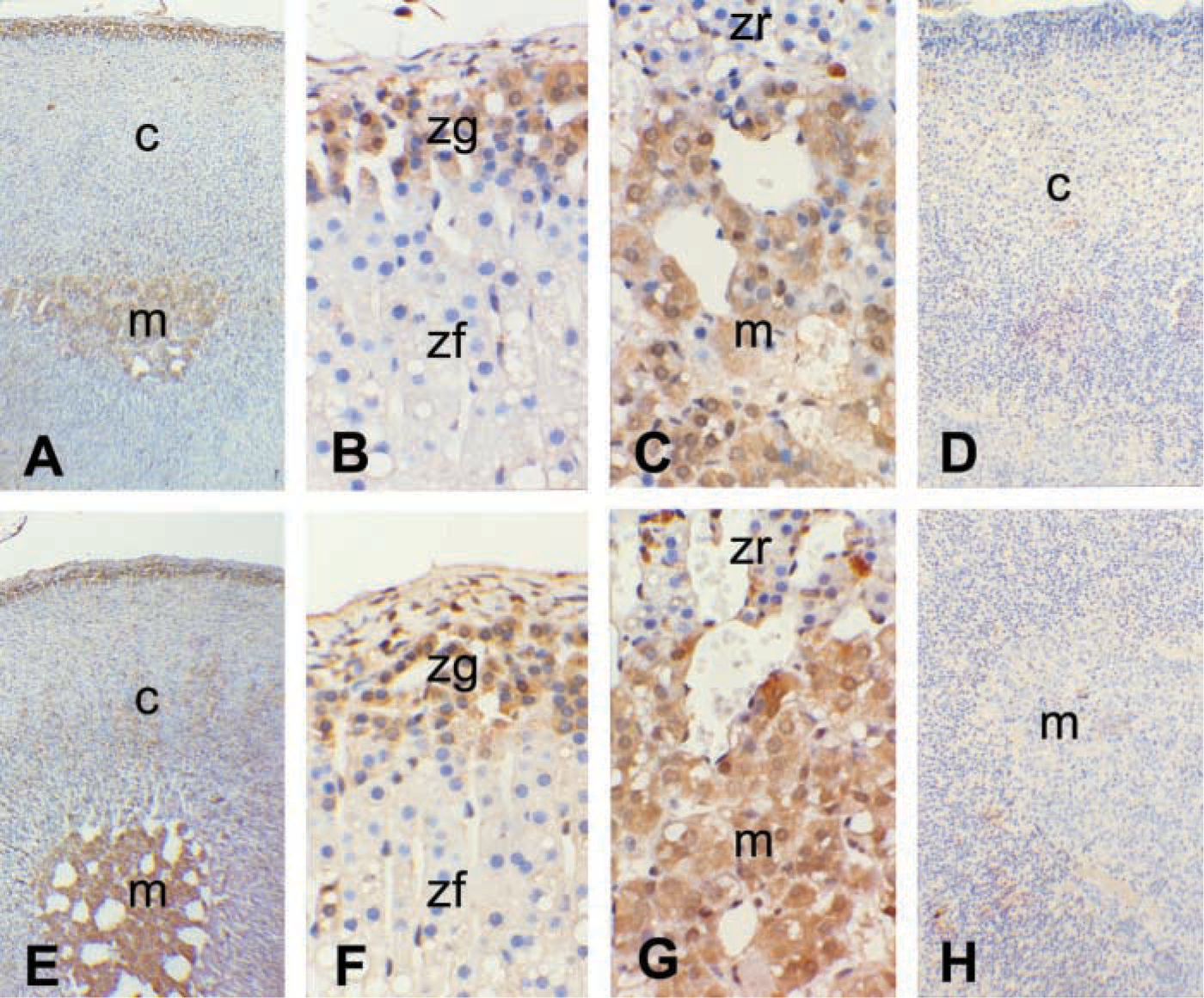
Representative photomicrographs show immunostaining of nNOS in the adrenal gland of 4-day control (
Several investigations have demonstrated that NO, sodium nitroprusside, and 8-bromo-cyclic guanosine monophosphate (GMP) produce a marked inhibition of acetylcholine- and KCl-stimulated catecholamine secretion in bovine chromaffin cells (Oset-Gasque et al. 1994; Torres et al. 1994; Kumai et al. 1999). Conversely, NO was reported to have a stimulatory effect on the basal secretion of catecholamine evoked by low doses of nicotine (Oset-Gasque et al. 1994). Kim et al. (2003) indicate that NO may lead to long-term upregulation of catecholamine synthesis via induction of TH, dopamine, β-hydroxylase, and PNMT gene expression in bovine chromaffin cells. Recently, Carabelli et al. (2002) showed that NO regulates the rate of catecholamine release by inhibiting neuroendocrine Ca(V)1 L-channel gating via cGMP-dependent protein kinase in cell-attached patches of bovine chromaffin cells. A direct inhibitory effect of NO on aldosterone and corticosterone production has also been described in adrenal glomerulosa (Salemi et al. 2001) and fasciculata (Cymeryng et al. 1998,1999,2002) cells. In addition to the modulation of NO on catecholamine and aldosterone production, endogenous NO is found to be capable of regulating adrenal blood flow (Breslow et al. 1993; Cameron and Hinson 1993). Therefore, further study is indicated to learn whether NO may affect adrenomedullary function in volume depletion.
In summary, our results show that the levels of nNOS mRNA and protein in the adrenal medulla of water-deprived rats were markedly enhanced in conjunction with these increases of intraadrenal TH mRNA and PNMT mRNA from 12 hr to 4 days, of 24-hr urinary epinephrine and norepinephrine excretion rates, and of serum aldosterone levels. The findings of this study indicate that the upregulation of nNOS synthesis in the adrenal medulla may be involved in the alteration of adrenomedullary function in the face of volume depletion.
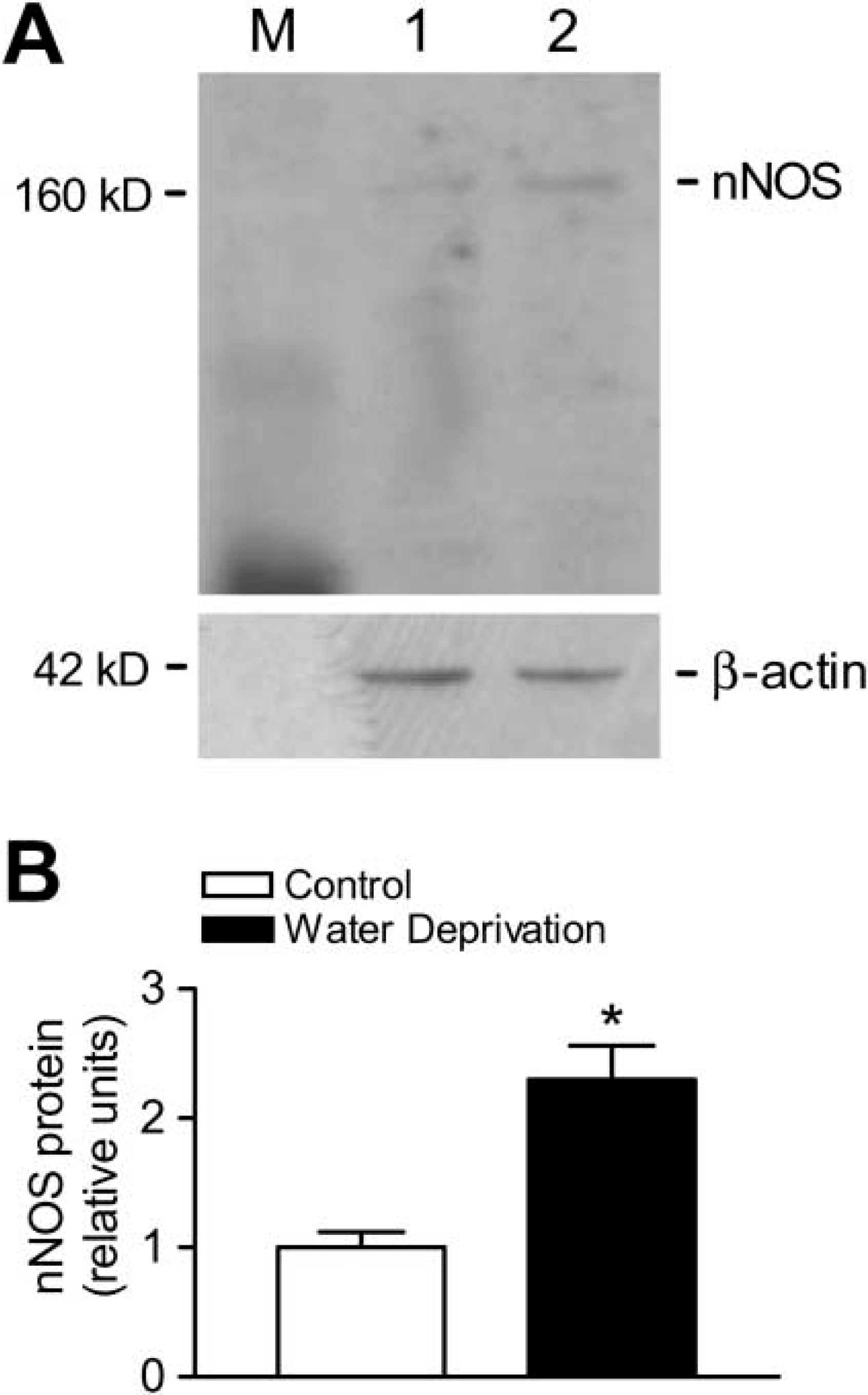
Expression of nNOS protein is upregulated by water deprivation. (
Footnotes
Acknowledgements
This work was supported by grants (NSC 90-2314-B-037-047 and NSC 91-2314-B-037-233) from the National Science Council, Taiwan; and grant RE86M018C from National Health Research Institutes, Taiwan (FJL).
We are indebted to Dr Chiang-Shin Liu, Dr Kun-Bow Tsai, Ms Chia-Lin Chen, Ms Yu-Hsiu Tung, Ms Pei-Chun Tsai, Mr Ming-Shih Li, Ms Yu-Hsiu Lee, Mr Chu-Ho Hung, and Ms Ching-Wen Wu for their excellent assistance.