
Abstract
A local renin-angiotensin system (RAS), including specific angiotensin II receptor subtypes, is present in the rat ovary. Immunohistochemistry using a polyclonal antibody and mRNA in situ hybridization were performed on perfusion-fixed, paraffin-embedded ovaries obtained from untreated sexually mature, normally cycling rats. Immunofluorescent staining revealed an exclusive and distinct labeling of follicular granulosa cells showing a plaque-like expression pattern at the cell borders, being detectable in different stages of atretic degeneration. On adjacent sections the expression of the respective mRNA could be shown in granulosa cells of the same follicle. The AT2 receptor may be implicated in the ovarian atretic process by influencing follicular cell-cell communication.
Keywords
I
In the present experiments, ovaries of adult female Sprague-Dawley rats, irrespective of the stage of the estrous cycle, were examined. Female rats were kept under standard laboratory conditions as described earlier (Obermüller et al. 1998). Animals were subjected to perfusion-fixation through the distal abdominal aorta using 2% paraformaldehyde dissolved in PBS, pH 7.4. Ovaries were removed and embedded in paraffin for subsequent procedures. Both ovaries from five animals were investigated. From each ovary, 10-15 sections were cut for analysis.
For IHC, sections 2-4-μm thick were deparaffinized and washed in bidistilled water. Next, sections were equilibrated in 10 mM citric acid, pH 6.0, for 10 min, microwaved at 500 W for 5 min, and again at 250 W for 10 min. Slides were allowed to cool to room temperature (RT), rinsed in PBS, and incubated in blocking solution (2% bovine serum albumin in PBS) for 30 min. The sections were then incubated with a rabbit polyclonal anti-AT2 antibody (H-143; Santa Cruz Biotechnology, Santa Cruz, CA). This antibody is directed against amino acids 221-363 from the carboxy terminus of human AT2 and cross-reacts with the rodent AT2 receptor, i.e., from K-ras-transformed rat kidney cells. The amino acid sequence of the human AT2 receptor is overall 92.6% identical to the rat AT2 receptor sequence (Tsuzuki et al. 1994). In the carboxy terminal region chosen for antibody generation, the homology is exactly 97.2%.
The antibody was applied in blocking solution at a dilution of 1:400 for 2 hr at RT and then overnight at 4C. Slides were then rinsed three times in PBS and incubated with a Cy3 (fluorochrome)-coupled secondary antibody produced in goat (Dianova; Hamburg, Germany) for 1 hr at RT at a dilution of 1:600. After washing three times for 10 min in PBS, slides were mounted in PBS-buffered glycerol. Control incubations on adjacent sections were carried out using 4-10% normal rabbit serum or blocking solution in place of the primary antibody.
Ovarian sections adjacent to those used for IHC were also subjected to non-radioactive mRNA ISH using a rat AT2 cDNA for riboprobe synthesis. The method for non-radioactive ISH has been described earlier for rat ovarian cryostat sections (Obermüller et al. 1998) and was adapted for paraffin sections used in this study. In brief, after deparaffination and postfixation in 4% paraformaldehyde at RT, slides were treated with proteinase K (Roche; Roche Diagnostics, Mannheim, Germany) at a concentration of 8 μg/ml in PBS for 30 min at 37C. Slides were then postfixed again in 4% paraformaldehyde for 1 min and washed in PBS before the prehybrization procedure continued. The hybridization and post-hybridization washing steps and the immunological detection using an antidigoxigenin alkaline phosphatase-coupled antibody were carried out as described (Obermüller et al. 1998).
A distinct and specific immunostaining for the AT2 receptor could be found in follicular granulosa cells of a vast majority of antral follicles (~85% of over 100 antral follicles analyzed), rarely in preantral follicles, but not in cells of corpora lutea (Figures 1A-1C). At higher magnification, AT2 receptor immunoreactivity showed a punctate to plaque-shaped expression pattern, which can be attributed to the cell surface of the granulosa cells (Figure 1D). Even in advanced stages of atresia when the follicles show reduced cell numbers and considerable damage to granulosa cells, immunoreactivity could still be observed (Figure 1E). The IHC expression of AT2 was found exclusively in granulosa cells and not in underlying theca cells (Figure 1E).
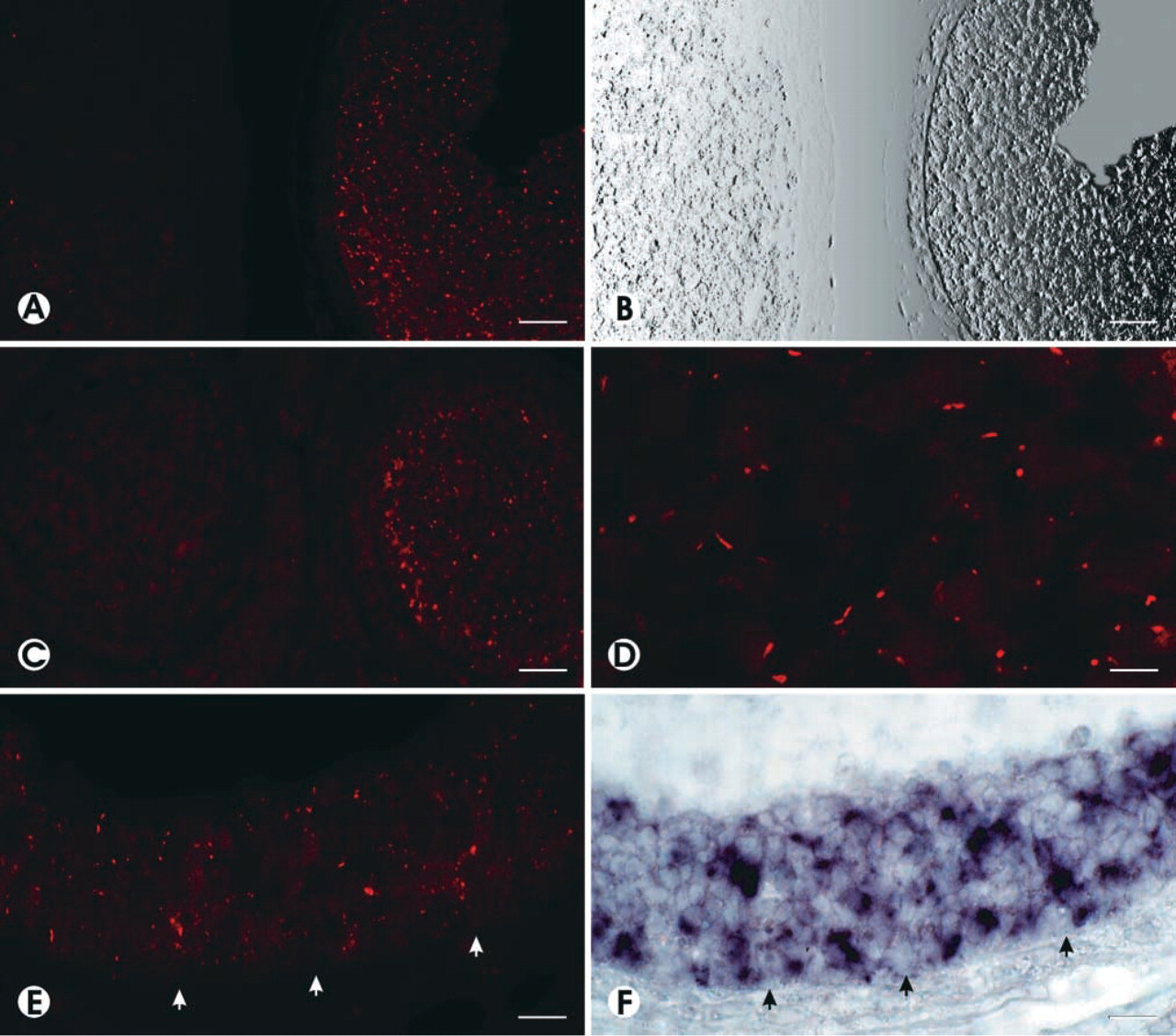
IHC for AT2 receptor protein (immunofluorescence in
The IHC detection of the AT2 protein in follicular granulosa cells was paralleled by demonstration of robust AT2 mRNA expression by ISH on adjacent sections of the same follicle (compare Figures 1E and 1F). The strong and exclusive follicular expression of AT2 mRNA detected by ISH on paraffin sections also corroborates the findings obtained previously on cryostat sections (Obermüller et al. 1998).
Our data are in good agreement with the results of the earlier experiments demonstrating follicular AT2 receptors in atretic follicles by autoradiographic binding studies (Husain et al. 1987; Daud et al. 1988; Pucell et al. 1991). Those studies found that either an entire follicle (meaning the majority or all follicular cells in a follicle) was positive for the AT2 receptor in terms of grain density or that a follicle was negative. This observation is paralleled by the present IHC results in which positive antral follicles showed staining in the majority of the cells of a given follicle. It is not clear at which stage of follicular atresia the AT2 receptor is already present. However, our IHC findings indicate its expression even in the early stages of atresia.
Functional studies have shown that the AT2 receptor mediates antiproliferative effects and apoptosis in different cells (Stoll et al. 1995, Yamada et al. 1996). In addition, its involvement in the follicular atretic process is suggested but has not yet been defined. Because atresia is considered as an all-or-none event, one can hypothesize that intercellular communication might be involved in the coordination of atresia and apoptosis. The distinct expression of the AT2 receptor on the cell surface with a plaque-like appearance (resembling intercellular plaques) may hint at an involvement in cell-cell communication between follicular granulosa cells. However, this assumption remains speculative and awaits future investigation.
Several studies show that there are marked species differences in the intraovarian AT2 receptor expression. Interestingly, in the bovine ovary receptor auto-radiography showed intense AT2 receptor binding in the theca cell layers, especially in the theca externa surrounding the majority of antral follicles. Moreover, the AT2 receptor is also present in tissue encompassing the corpora lutea. However, in contrast to the situation in the rat, no AT2 binding is seen in follicular granulosa cells (Schauser et al. 2001). The role of the AT2 receptor in theca cells in this species remains puzzling. It is possible that it participates in the regulation of folliculogenesis and also in the development of the corpora lutea.
In the postpubertal porcine ovary, the AT1 receptor and also the AT2 receptor could be detected by IHC in zones of neovascularization (Shuttleworth et al. 2002). Although here granulosa cells of healthy antral follicles exclusively exhibited staining for AT1 receptor, the authors also report on a subset of presumably atretic follicles with AT2 receptor-expressing granulosa cells, the latter finding resembling in part the situation in the rat (Shuttleworth et al. 2002). Although in the mamalian ovary the AT2 receptor may be implicated in differentiation and atresia in some cell types of at least some species, other regulatory roles in steroidogenesis or vascular growth should also be considered as possible.
In summary, the present IHC experiments localize the AT2 receptor to the cell surface of rat ovarian follicular granulosa cells and corroborate previous studies identifying AT2 receptor expression by several other techniques in this particular cell type.
Footnotes
Acknowledgements
We thank Ms U. Horr for preparation of the figures.