
Abstract
Somatostatin acts on specific membrane receptors (sst1–5) to inhibit exocrine and endocrine functions. The aim was to investigate the distribution of sst1–5 in pancreatic islet cells in normal mice and rats. Pancreatic samples from five adult C57BL/6 mice and Sprague-Dawley rats were stained with antibodies against sst1–5 and insulin, glucagon, somatostatin, or pancreatic polypeptide (PP). A quantitative analysis of the co-localization was performed. All ssts were expressed in the pancreatic islets and co-localized on islet cells to various extents. A majority of the β-cells expressed sst1–2 and sst5 in mouse islets, while ≤50% in the rat expressed sst1–5. The expression of sst1–5 on α-cells did not differ much among species, with sst2 and sst5 being highly expressed. About 70% of the 8-cells expressed sst1–4 in the rat pancreas, whereas 50% of the islet cells expressed sst1–5 in the mouse. Furthermore, 60% of the PP-cells expressed sst1–5 in the mouse, while the rat islets had lower values. Co-expression with the four major islet hormones varies among species and sst subtypes. These similarities and differences are interesting and need further evaluation to elucidate their physiological role in islets.
Keywords
S
The physiological activities of somatostatin are mediated by high-affinity interactions with a family of G-protein-coupled receptors. Thus far, five different somatostatin receptor (sst1–5) subtypes have been cloned (Yamada et al. 1992a,b,1993). They are distinguished by their pharmacological specificity for different analogues, their tissue distribution, their regulation, and their intracellular signaling pathways (Florio and Schettini 1996; Patel 1999). Inhibition of hormone secretion is considered to be mediated mainly through sst2 and sst5 (Chen et al. 1994) and growth-inhibiting signals by sst1, sst2, and sst5 (Buscail et al. 1994,1995), whereas sst2 and sst3 are believed to be involved in somatostatin-induced apoptosis (Sharma et al. 1996; Teijeiro et al. 2002). The sst4 is the least studied receptor, and therefore not much is known about the putative function of this receptor.
The cloning of sst genes made subtype-specific probes available, which allowed the investigation of sst expression at the mRNA level. The tissue distribution pattern of sst was studied in the central nervous system and peripheral tissues by RT-PCR in rat (Raulf et al. 1994). The study showed expression of all five ssts in the brain and pituitary, whereas other investigated organs, e.g., lungs, liver, and pancreas, varied in their tissue sst expression. The mRNAs for all five ssts were also detected in mouse brain (Fehlmann et al. 2000) and in the retina (Cristiani et al. 2002), whereas only sst3–5 were expressed in the kidney (Bates et al. 2003). However, to elucidate the more exact cellular distribution of sst, immunohistochemical (IHC) investigations are needed.
Recently developed antibodies selective for sst subtypes have been raised to investigate the expression of sst in various tissues (Raulf et al. 1994; Hunyady et al. 1997; Kumar et al. 1999; Portela-Gomes et al. 2000; Kimura et al. 2001; Cristiani et al. 2002; Bates et al. 2003). The pancreas is an important target for the action of somatostatin, and the nature of the sst expressed in islet cells of the pancreas is therefore of particular interest. In the pancreas, somatostatin is produced in islet δ-cells and acts directly on the islet cells to inhibit secretion of insulin, glucagon, somatostatin, and pancreatic polypeptide (PP) (Patel 1999). Thus far, only expression of sst2 and sst5 has been demonstrated in the pancreatic islets of rat (Hunyady et al. 1997; Mitra et al. 1999; Kimura et al. 2001). To our knowledge, the expression and distribution of all five ssts in pancreatic islets of mice and rats have not been reported. Furthermore, we here show the co-expression of all five ssts with the four major islet cell types, β-, α-, δ-, and PP-cells, in mice and rats. Knowledge about the tissue- and cell-specific expression of ssts is of major importance in understanding their physiological function. Therefore, the aim of this study was to investigate the co-localization of sst1–5 in the pancreatic islet cells of normal mice and rats.
Materials and Methods
Animals
For this study, five adult male C57BL/6 mice (B&K Universal; Sollentuna, Sweden) and five adult male Sprague-Dawley rats (Biomedical Center; Uppsala, Sweden) were used. The animals had free access to water and pelleted food and were housed in a room with a 12-hr light/dark cycle. All experiments were approved by the local ethics committee at Uppsala University (Uppsala, Sweden).
Tissue Preparation
All animals were weighed and their blood glucose concentration was measured (Medisense; Waltham, MA). They were sacrificed by cervical dislocation and, according to good laboratory practice, the rats were anesthetized (Mebumal 60 mg/kg) before they were subjected to this procedure.
The abdomen was immediately opened by a transverse incision, followed by resection of the pancreas, which was fixed in 10% buffered formalin (Merck; Darmstadt, Germany) overnight. The tissues were then stored in 70% ethanol until they were embedded in paraffin. Sections 5 μm thick were cut and attached to Polysine glass slides (Menzel-Gläser; Braunschweig, Germany).
Development of Antibodies Against Different ssts
The production and specificity of subtype-specific somatostatin receptor antibodies have been described earlier (Portela-Gomes et al. 2000). In short, five amino acid sequences deduced from human sst1–5 (Yamada et al. 1993), were synthesized by a solid-phase system using Fmoc chemistry (Model 430A; Applied Biosystems, Foster City, CA). The peptides were purified by reverse-phase chromatography and analyzed by plasma desorption mass spectrometry (PDMS; Bioion 20, Bioion Nordic, Uppsala, Sweden). The amino acid sequences were selected to be specific for the respective sst and the homology was less than 48% to any other known protein sequence in the protein database SWISS-PROT (release 37.0, Dec. 1998), except for the respective sequences of ssts from other species. The selected sequences were amino acids 363–380 for sst1, amino acids 330–343 with an additional tyrosine residue at the N-terminal (Tyr0) for sst2, amino acids 366–381 with an additional tyrosine residue at the N-terminal (Tyr0) for sst3, amino acids 332–345 with an additional tyrosine residue at the N-terminal (Tyr0) for sst4, and amino acids 327–341 with an additional tyrosine residue at the N-terminal (Tyr0) for sst5. All peptides had amidated C-terminals.
The chosen sequence of peptides used to generate a selective antibody against sst1 was identical to both the mouse and the rat receptor sequence. The peptide used for sst2 differed by one amino acid compared to the mouse and two compared to the rat. These amino acids are located in the middle of the peptide and were not expected to interfere with the crossreactivity of the antibody. The peptide used to generate the sst3 differed by four amino acids from humans compared to the rat and mouse sequences. These amino acids are also located to a part of the peptide that was not expected to interfere with the crossreactivity of the antibody. In the sst4 peptide sequence there is an insertion of nine amino acids in the middle of the peptide sequence in both rats and mice. However, the C-terminal is identical with the human receptor sequence, and the sst4 antibody was therefore expected to crossreact with rat and mouse tissues. The peptide sequence for sst5 antibody production differed by one amino acid in both the mouse and rat compared to the human sequence. In this case, a lysine residue has been replaced by an arginine residue and should therefore not interfere with the crossreactivity of the antibody.
Before immunization the peptides were coupled to a carrier protein. Two mg peptide and 20 mg bovine serum albumin (BSA; ICN Biomedicals, Costa Mesa, CA) or keyhole limpet hemocyanogen were dissolved in a 50 mmol/liter sodium phosphate buffer at pH 7.4 containing 150 mmol/liter NaCl. Coupling was then induced by addition of 90 μl glutaraldehyde. The resulting complexes were injected into New Zealand White rabbits, using the intradermal injection technique to produce polyclonal antibodies (Vaitukaitis et al. 1971).
Morphological Examinations
Single Staining with sst1–5 Antibodies. The formalin-embedded preparations from pancreas specimens were stained for all of the five ssts. All solutions were diluted and sections were washed in Tris-buffered saline (TBS, pH 7.6) and incubated at room temperature unless otherwise stated. For staining of ssts, the sections were pretreated by boiling the tissues in a microwave oven for 10 min in 50 mM Tris-HCl (pH 8.0). Nonspecific binding of secondary antibody was blocked by incubation for 30 min with normal goat serum (Vector Laboratories; Burlingame, CA). Incubations of the primary antibodies against sst1, sst2, sst3, sst4, and sst5, respectively, were performed overnight at 4C in a humidified chamber at titers of 1:300, 1:600, 1:500, 1:500, and 1:750, respectively, each diluted in TBS with 1% BSA. Unbound antibody was removed by washing thrice for 5 min. The secondary antibody, biotinylated goat F(ab')2 anti-rabbit IgG (1:200; Southern Biotechnology, Birmingham, AL), was applied to the tissues for 45 min. Unbound antibody was removed by washing thrice for 5 min. The immune reaction was amplified by an avidin-biotin complex coupled to alkaline phosphatase (Vectastain ABC-AP; Vector Laboratories) and visualized with Vector Red (Vector Laboratories) as substrate. Finally, the tissues were counterstained with Mayer's hematoxylin (Histolab; Göteborg, Sweden) and permanently mounted with Mountex (Histolab) and coverslips.
Double Immunofluorescence Staining of sst1–5 and Islet Cells. The formalin-embedded preparations from pancreas specimens were simultaneously stained for all of the five ssts together with insulin, glucagon, somatostatin, or PP. All solutions were diluted and sections were washed in TBS and incubated at room temperature unless otherwise stated. The sections were pretreated by boiling the tissues in a microwave oven for 10 min in 50 mM Tris-HCl (pH 8.0). Nonspecific binding of the secondary antibody was blocked by incubation for 30 min with normal donkey serum (1:5; Jackson ImmunoResearch Laboratories, West Grove, PA). Each of the rabbit anti-sst1–5 antibodies was mixed with chicken anti-insulin (1:750; Immunsystem, Uppsala, Sweden), chicken anti-glucagon [1:400; a kind gift from Associate Professor Anders Larsson; raised against human glucagon (Novo Nordisk; Bagsvaerd, Denmark)], sheep anti-somatostatin (1:25; Guildhay, Guildford, UK) or sheep anti-PP antibodies (1:25; SeroTech, Oxford, UK), and the sections were incubated overnight at 4C in a humidified chamber. All primary antibodies were diluted in TBS with 1% BSA. Unbound antibody was removed by washing thrice for 5 min. To perform double immunofluorescence and visualize the distribution of sst1–5 and islet hormones, sections were incubated with a cocktail consisting of the secondary antibodies Cy3-conjugated donkey anti-rabbit IgG (1:100; Jackson ImmunoResearch) + Cy2-conjugated donkey anti-sheep IgG (1:100; Jackson ImmunoResearch) or Cy2-conjugated donkey anti-chicken IgG (1:100; Jackson ImmunoResearch) for 45 min. Unbound antibody was removed by washing thrice for 5 min. The tissues were then mounted with mounting medium containing 50% TBS + 50% glycerol (Calbiochem; San Diego, CA) and coverslips.
Staining Specificity
To investigate the specificity of sst antibodies, a preincubation for 24 hr at 4C of primary antiserum with the relevant antigen (10 nmol/ml diluted antibody solution, respectively) was performed before application to the sections. The sst antigens were the synthesized peptides used for immunization.
Light Microscopic Evaluation
The intensity and distribution of the sst stainings were examined and scored semiquantitatively: + + +, strong positive staining; ++, clear positive staining; +, weak positive staining; −, not detected. Moreover, the location of the positive staining of ssts was scored (center/periphery) referring to whether the sst was located in the center of the islet or only in the periphery. The pancreatic sections and the different rankings were evaluated with the examiner being unaware of the origin of the sections and the identity of the sst antibody used. Sections from each pancreas were stained on three different occasions with each sst antibody and 8–10 islets were analyzed per animal. Because the islets in a given section varied in size, they were randomly selected to include both large and small islets.
Evaluation of Immunofluorescence
The sections were examined in a Leica Leitz DMR fluorescence microscope (Leica Microsystems; Wetzlar, Germany) equipped with filters of 492 nm to 510 nm for Cy2 (green) and 550 nm to 570 nm for Cy3 (red).
Pictures from a Zeiss Axiocam camera (Carl Zeiss; Oberkochen, Germany) of each pancreatic islet, using both filters, were merged together with Adobe PhotoShop 7.0 software (Adobe; San Jose, CA), in which a yellow color indicated co-expression of sst subtype with any of the four islet hormones tested in this study. The results are expressed as a percentage of sst-positive cells in relation to the total number of the respective islet cell type in a specific pancreatic islet.
Statistical Analysis
Statistical analysis was performed by comparing groups of data with Student's t-test, where p<0.05 was regarded as significant. The results are presented as means ± SEM (n = 5). The computer program used was SigmaStat 2.0.
Results
Animals
The mice (n=5) allocated to this study weighed ∼30 g and had a blood glucose concentration of 8.8 ± 0.1 mmol/liter, whereas the rats (n=5) weighed ∼300 g and their blood glucose concentration was 5.9 ± 0.6 mmol/liter.
Distribution of ssts in the Islets of Langerhans
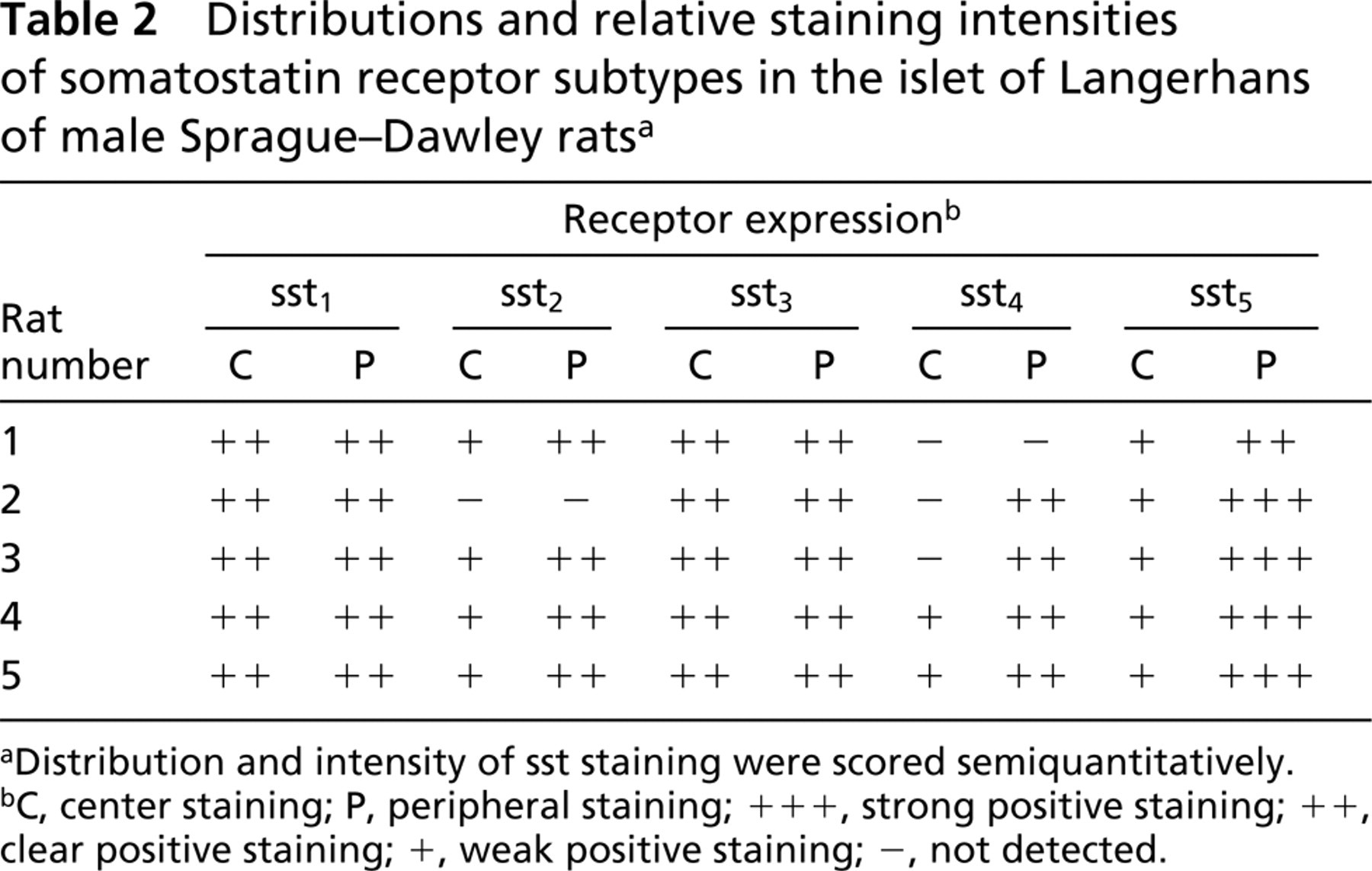
According to the area of the islet of Langerhans, different distributions and relative staining intensities of sst subtypes were observed. These findings are summarized in Table 1 for mice and in Table 2 for rats. Figure 1 shows representative expression of the five ssts in the pancreatic islets of a mouse (Figures 1A-1E) and a rat (Figures 1F-1J).
In both mice and rats we found a homogenous staining for sst1 of the entire islet profiles, but the staining was more intense in mice. The staining intensity was essentially similar in the islet center and periphery.
Distributions and relative staining intensities of somatostatin receptor subtypes in the islet of Langerhans of male C57BL/6 mice a

aDistribution and intensity of sst staining were scored semiquantitatively.
bC, center staining; P, peripheral staining; + + +, strong positive staining; ++, clear positive staining; +, weak positive staining; −, not detected.
The staining for sst2 was more pronounced in the periphery of the islets compared to the center in both mice and rats. One rat was negative for sst2, and in some small islets there was no difference in staining intensity between the central and the peripheral part of the islet. Moreover, acinar cells were frequently stained for sst2 in both mice and rats.
The pancreatic islets of all rats were positive for sst3, whereas in two mice we could not detect any staining for sst3. Furthermore, in all sst3-positive islets we observed a homogenous localization of sst3 in the entire islet.
In two mice we did not detect any staining for sst4 in the islets of Langerhans, and in the other three mice the entire islet had weak positive staining for sst4. However, in rats we observed a different staining pattern, with intense staining of the periphery of the islets and faint or no staining of the central parts of the islets. One rat was negative for sst4.
In mice we found more intense staining for sst5 compared to rats. The staining appearance was similar in all animals, with more pronounced staining in the periphery of the islets. In some small islets the entire islet had homogenous staining for sst5.
Distributions and relative staining intensities of somatostatin receptor subtypes in the islet of Langerhans of male Sprague-Dawley rats a
aDistribution and intensity of sst staining were scored semiquantitatively.
bC, center staining; P, peripheral staining; + + +, strong positive staining; ++, clear positive staining; +, weak positive staining; −, not detected.
Pancreatic islets of a C57BL/6 mouse (
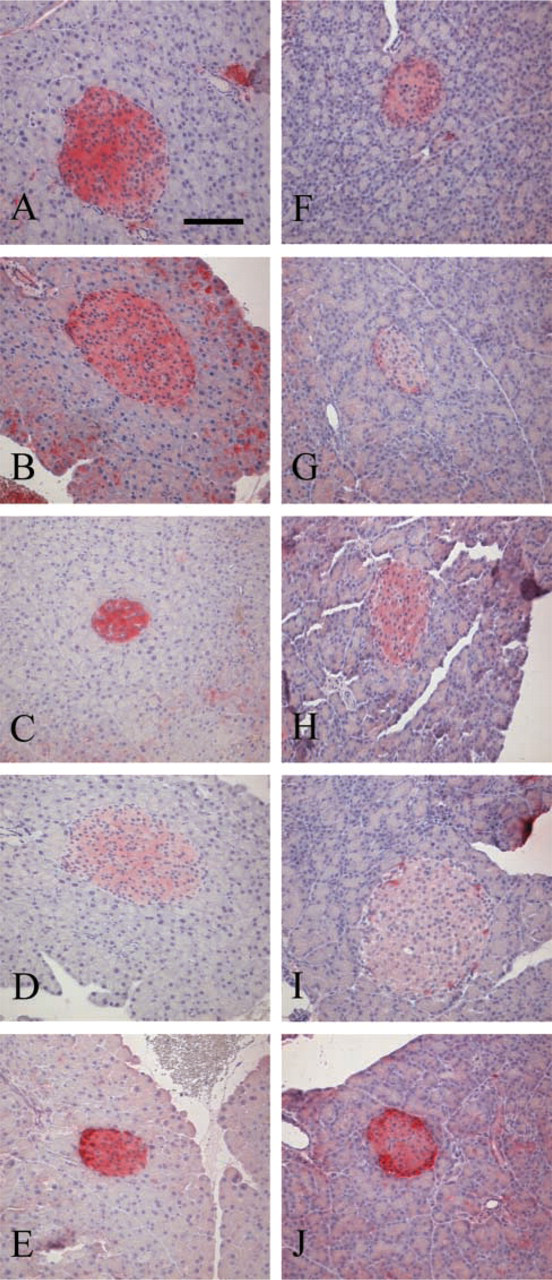
Figure 2 shows the control staining from a representative rat pancreas. The left column shows the pancreatic sections normally stained with all five sst antibodies (Figures 2A-2E). When we preincubated the sst-specific antibodies with the peptides used for immunization, the staining for all receptors in both mice and rats was completely blocked (Figures 2F-2J).
Pancreatic islets of a Sprague-Dawley rat stained with sst1–5-selective antibodies (
Immunofluorescence staining for sst1–5 and Islet Cells
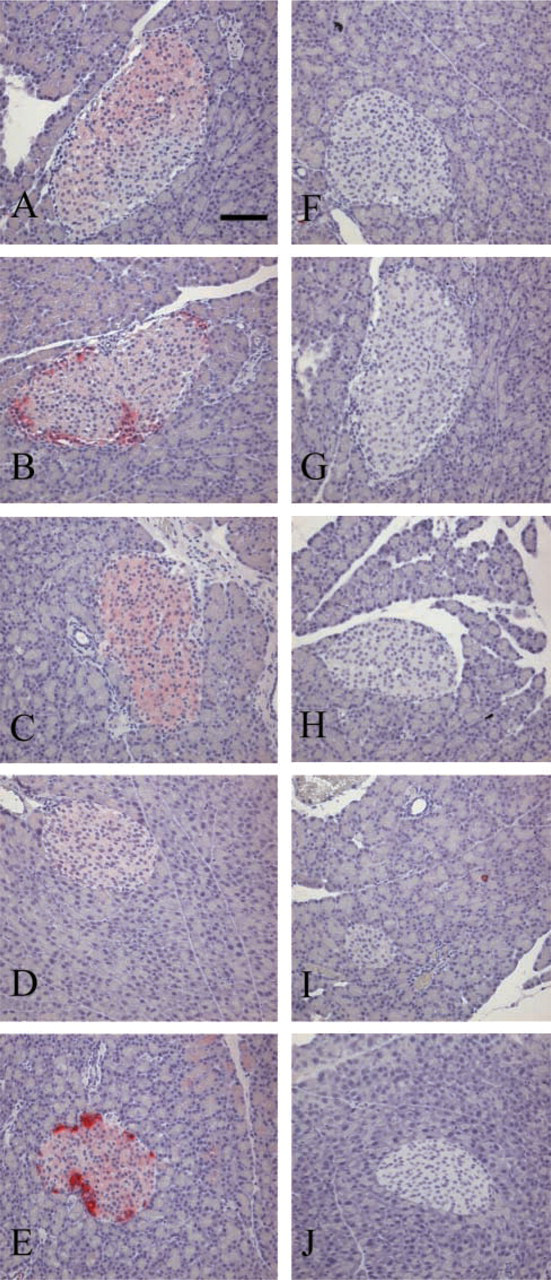
β-Cells and sst1–5. Figures 3A-3O show Cy3-conjugated immunofluorescence distribution of sst1–5 (red, Figures 3A-3E) in a representative islet sample of a mouse. β-Cells were identified in the same islet by Cy2-conjugated immunofluorescence with insulin antibody (green, Figures 3F-3J). Overlapping ssts and β-cells revealed co-expression of sst1–5 with insulin (yellow, Figures 3K-3O). Figure 5 shows the quantitative analysis of the pancreatic islet with insulin and sst1–5, in which the results from the mouse are represented by black bars and the rat by white bars. It shows that sst1, sst2, and sst5 were co-expressed in a majority of all insulin-positive cells in the pancreatic islets of the mouse. However, in the pancreatic islets of the rat, sst1, sst2, and sst5 were expressed only in about half of all β-cells. The fraction co-localization of sst3- and insulin-positive cells did not significantly differ between the species. Moreover, sst4 was expressed on β-cells to almost the same degree in both mice and rats.
α-Cells and sst1–5. Figures 4A-4E show representative mouse islet sections processed for co-expression of sst1–5 with glucagon, in which the co-localization yields a yellow color. As expected, the α-cells were fewer than the β-cells and the cells were distributed mostly in the periphery of the islet. Quantitative analysis of sst1 and sst5 in mouse α-cells shown in Figure 6 demonstrated lower values compared to rat α-cells. The co-localization of the other ssts was similar in mouse and rat islets, with sst2 and sst5 being strongly co-expressed to about 70%, while sst3 and sst4 were only expressed in close to 20% of the α-cells in both species. A slight significant difference was found for sst5 between the species.
8-Cells and sst1–5. Figures 4F-4J show the results of sst1–5 co-expressed with endogenous somatostatin in representative islet sections of a rat. The somatostatin cells were found mostly around the mantle zone of the islet, and to a lesser degree than both the β- and the α-cells. As shown in Figure 7, the extent of co-localization of sst1 on somatostatin-positive cells in mouse and rat islets was almost the same. Comparing the two species, however, we found that the rat pancreas had a somewhat stronger co-localization of sst3 and sst4 than the mouse. As for sst1, the co-localization of sst2 and sst5 on 8-cells was similar in both mouse and rat islets.
PP-Cells and sst1–5. Figure 4 shows the co-expression of sst1–5 with PP-cells in a representative pancreatic islet of a rat (Figures 4K-4O). PP-Cells were the least abundant islet cell type in this study, and were identified as a sparse population of cells distributed in the mantle zone of the islet. Figure 8 summarizes the quantitative analysis of the sst together with PP-cells in mouse and rat. All five ssts were expressed in almost the same numbers and in a majority of PP-positive cells in mouse islets, whereas the co-localization of sst in the rat was only expressed in about 15% for all five subtypes.
Pancreatic islets of a C57BL/6 mouse stained with sst1–5-selective antibodies (red cells,
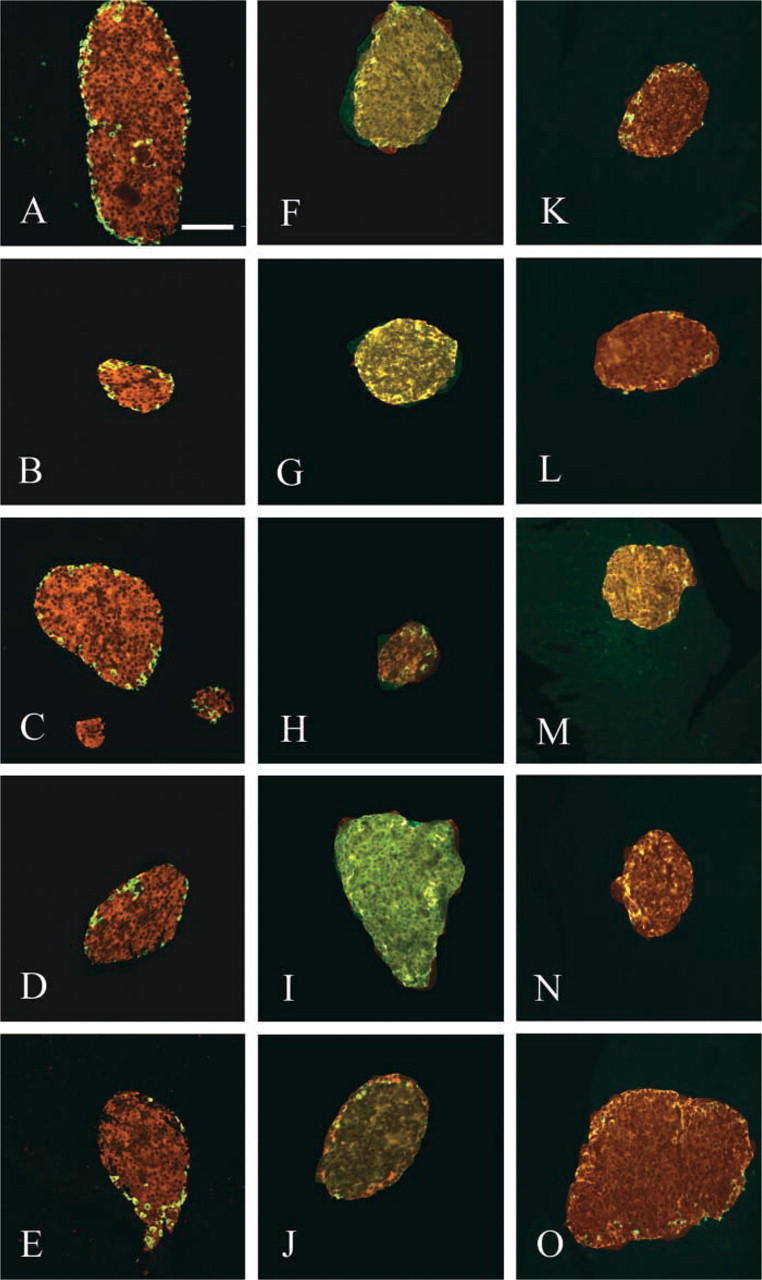
Pancreatic islets of a C57BL/6 mouse co-expressing sst1–5 and glucagon-secreting cells (yellow cells,
Discussion
IHC studies of sst subtype expression in various tissues of mouse and rat have recently been performed (Hunyady et al. 1997; Fehlmann et al. 2000; Kimura et al. 2001; Bates et al. 2003). Another study has demonstrated the co-localization of sst5 and insulin in rat pancreatic islets (Mitra et al. 1999). However, to our knowledge the present study is the first description of the staining pattern of all five somatostatin receptor subtypes in the pancreatic islets of rodents. We also report the co-localization of sst1–5 with the four major islet hormones; insulin, glucagon, somatostatin, and pancreatic polypeptide cells, in the pancreas of mice and rats. For this purpose we used polyclonal antibodies that recently have been proved specific for human sst receptor subtypes (Portela-Gomes et al. 2000). The human sst1–5 peptide sequences against which these antibodies were raised are similar to the corresponding sequences in mouse and rat. The peptide sequence used for sst1 is identical in humans, rats, and mice, whereas the sequences used for sst2–5 differ in only a few amino acids located in parts of the peptide that are considered not to be interfering with the crossreactivity of the developed antibodies for the species used in this study. It was therefore likely that these antibodies were suitable also for mouse and rat tissue. To verify this, the specificity of the human sst antibodies on mouse and rat tissue was investigated by preincubating the sst subtype-specific antibodies with the peptides used for immunization. We were thereby able to completely block the staining for all receptors in both mice and rats.
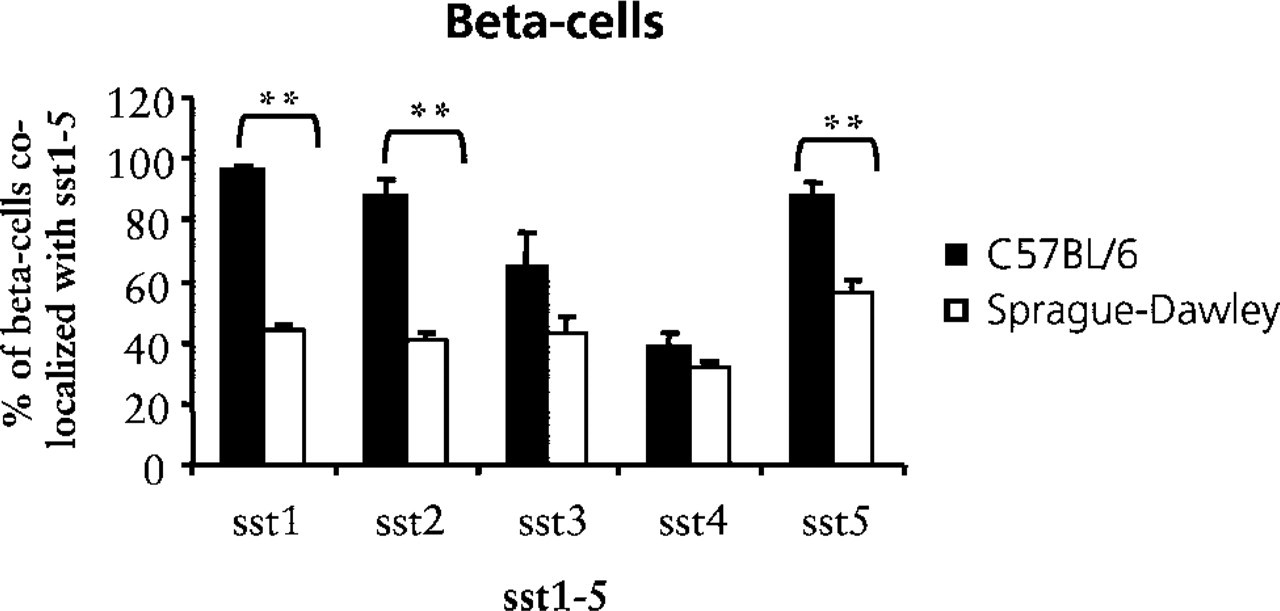
Quantitative analysis of the expression of sst1–5 in β-cells in pancreatic islets of mice (black bars) and rats (white bars). Bars represent the mean ± SEM (n = 5) of percentage cells positive for a given sst subtype. For each animal, three sections were analyzed and in total 20–30 islet profiles per animal were measured. 5p<0.01.
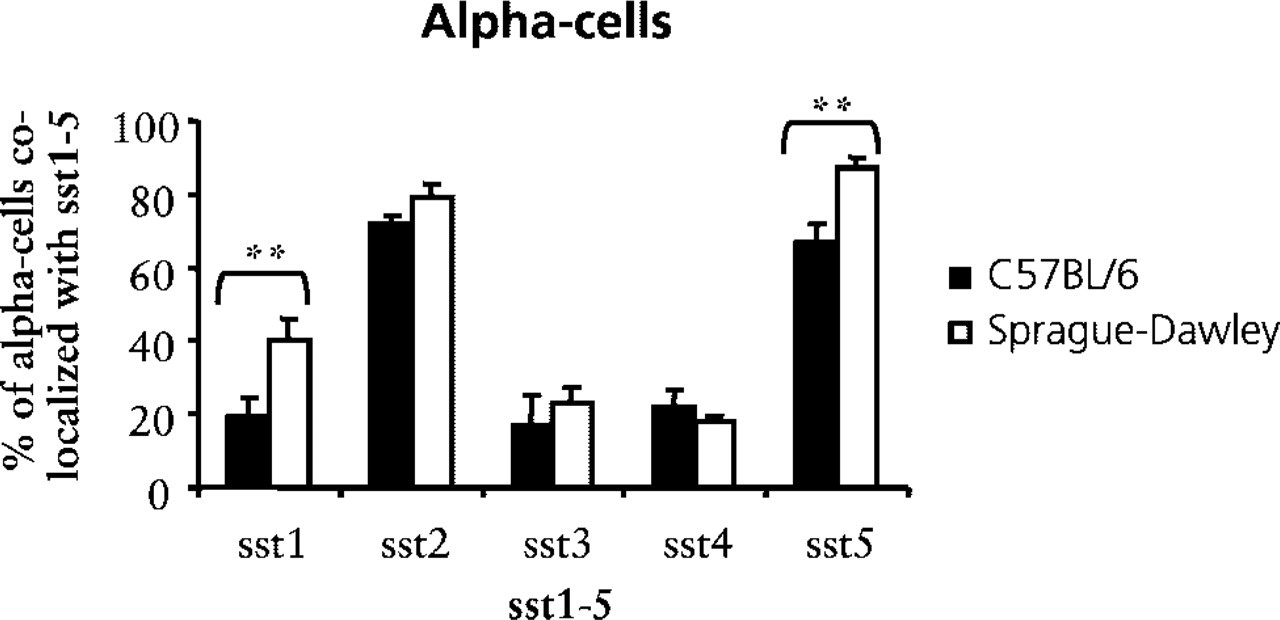
Quantitative analysis of the expression of sst1–5 in α-cells in pancreatic islets of mice (black bars) and rats (white bars). Bars represent the mean ± SEM (n = 5) of percentage cells positive for a given sst subtype; see Figure 5. 6p<0.01.
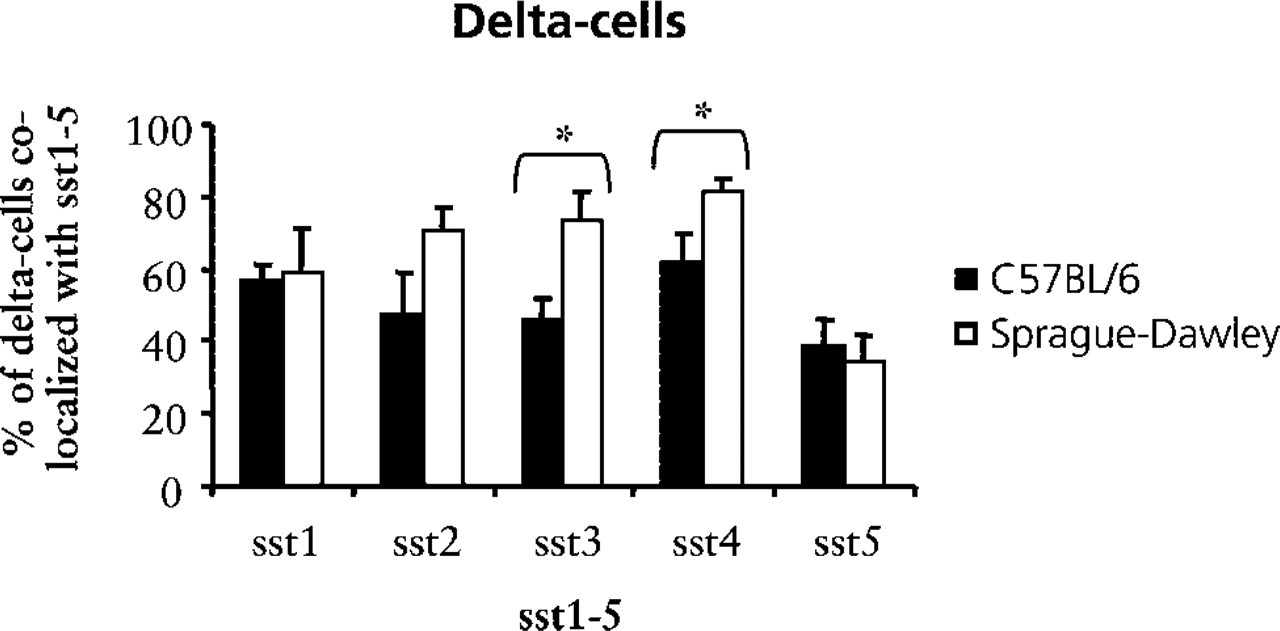
Quantitative analysis of the expression of sst1–5 in 8-cells in pancreatic islets of mice (black bars) and rats (white bars). Bars represent the mean ± SEM (n = 5) of percentage cells positive for a given sst subtype; see Figure 5. ∗p<0.05.
Recent studies have reported that sst subtypes are expressed in human pancreatic islets (Kumar et al. 1999; Portela-Gomes et al. 2000). In line with theses reports, we also observed that all five receptor subtypes were present in the rodent islets but that the staining pattern differed for sst1–5 among individual animals of the same species and between mice and rats. We found that the entire pancreatic islet of mice and rats stained positive for sst1. Moreover, we showed a distinct staining pattern for sst2 and sst5 in mice and rats, with weak central islet staining and intense staining of peripheral islet cells. An immune reactivity for sst2 in acinar cells was also found in both mice and rats, which is supported by previous studies (Hunyady et al. 1997; Kimura et al. 2001). Expression was not detected for any of the other sst subtypes in the exocrine tissue of either mice or rats.
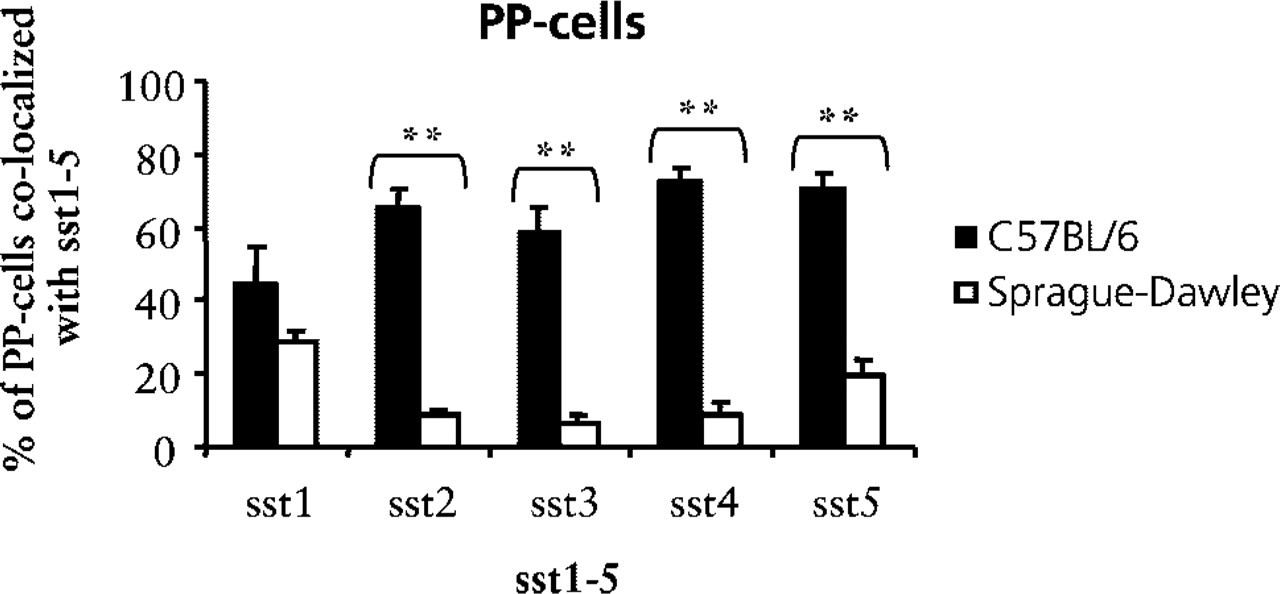
Quantitative analysis of the expression of sst1–5 in PP-cells in pancreatic islets of mice (black bars) and rats (white bars). Bars represent the mean ± SEM (n = 5) percent of cells positive for a given sst subtype; see Figure 5. ∗∗p<0.01.
All rat islet cells stained positive for sst3, and in some mice the staining pattern was similar, whereas other mice did not express sst3 in their islets. The function of sst3 in pancreatic islets is presently unknown and needs further investigation.
By using RT-PCR analysis, Raulf et al. (1994) demonstrated the presence of all sst subtypes in whole rat pancreas preparations, except for sst4. In mice, we here report weak positive staining for sst4 in most islet cells in some animals and no staining in other animals. Interestingly, in this study we detected all sst subtypes in rat islets and the staining pattern for sst4 was different than that of mice, with more intense staining in the islet mantel zone. The sst4 is the receptor subtype that has been studied the least, and the function of sst4 in islet cells is essentially unknown.
In comparing the amount of co-localization of ssts with the various islet cell types, we found intriguing differences between the two species. A high percentage of β-cells expressed sst1, sst2, and sst5 in mouse islets. In contrast, less than half of the β-cells expressed those receptors in the rat. Several reports have demonstrated that inhibition of insulin secretion is mediated via sst5 in rat and mouse (Strowski et al. 2000,2003). Because it is now known that sst1 and sst5 heterodimerize in a tissue on stimulation, this may be an explanation for why sst1 and sst5 are expressed in the mouse islet to almost the same degree (Rocheville et al. 2000). An interesting possibility in this context is that receptor-positive and receptor-negative cells may represent two different functional populations of β-cells. Therefore, the former would be responsive to somatostatin-induced inhibition of insulin secretion, whereas the latter cells would not be affected by the hormone. However, it should be noted that a negative staining does not exclude the possibility of expression of the receptor below the detection level. Another issue to consider is whether the expression of sst receptors per cell is changing dynamically, (e.g., in relation to the metabolic state). This issue may be elucidated by in vitro experiments using isolated pancreatic islets.
Mouse PP-cells had a much higher degree of sst2–5-positive cells in islets compared to the rat. On the other hand, in the somatostatin-positive cells, about 75% of the cells co-expressed sst3–4 in the rat islet and only about 55% in the mouse. We found that the sst4 was the receptor subtype showing the most extensive expression in 8-cells in mouse and rat pancreatic islets. The significance of this finding is unclear because this receptor is the least studied and not much is known about its function in pancreas. The role of the highly expressed sst3 in 8-cells in rat pancreas is also unknown. Furthermore, only about 35% of the 8-positive cells did express sst5. It can be suggested that sst2 and/or sst1, rather than sst5, are involved in the inhibition of somatostatin in pancreas.
The most “conserved” islet cell type was the α-cell, with a fairly similar co-expression pattern in the two species for all five ssts. The receptors sst2 and sst5 were expressed in a majority of α-cells in mouse and rat islets. It has been reported from several studies that sst2 is the dominant inhibitor of glucagon secretion in mouse and rat (Rossowski and Coy 1994; Strowski et al. 2000), but the results from this study may suggest that the sst2 and sst5 act together to contribute to the inhibitory effect. Using sst2- and sst5-specific analogues in an in vitro study of glucagon secretion may elucidate this possibility.
By using double immunofluorescence staining Mitra et al. (1999) reported that sst5 was exclusively expressed in β-cells in the pancreatic islets of Sprague-Dawley rats and was not detected in other islet cells. We could not confirm this observation because we found also sst5 expression in a substantial fraction of the other hormone-expressing islet cells in Sprague-Dawley rats. Double immunofluorescence staining studies similar to the present study have been performed on normal human pancreatic tissue using sst antibodies (Kumar et al. 1999; Portela-Gomes et al. 2000). There appears to be a difference among species (Yamada et al. 1992a,b; Raulf et al. 1994). However, there are considerable overlapping expression patterns between human and rodent pancreatic islet cells. Moreover, when the present results from mouse and rat pancreas are compared with the human pancreas studies, it appears that the co-expression of sst2 on α-cells is in good agreement in humans and rodents (Kumar et al. 1999; Portela-Gomes et al. 2000).
In summary, the present investigation demonstrates that all sst subtypes are expressed in islets of Langer-hans of mice and rats. We report staining patterns that are in line with and partly different from those previously reported for human islets. In addition, some variations in expression patterns for sst subtypes between mice and rats were seen, which may reflect a difference in response to somatostatin in islet cells of the two species. In addition, the co-expressions of sst1–5 with the four major islet cell types varied among individual animals. These similarities and differences are interesting and need further evaluation to understand the functional role of the somatostatin receptors in islet cell physiology.
Footnotes
Acknowledgements
Supported by grants from the Swedish Research Council (72X-8273), the Swedish Diabetes Association, the Family Ernfors Fund, the Juvenile Diabetes Research Foundation, the Wallenberg Fund, the Novo Nordisk Fund, and the Swedish Society for Medical Research.
The technical expertise of Eva Törnelius and Ing-Britt Hallgren is gratefully acknowledged. The chicken anti-glucagon antibody was a kind gift from Assoc Prof Anders Larsson (Department of Medical Sciences, University Hospital; Uppsala, Sweden).