
Abstract
Leptin, the ob protein, regulates food intake and satiety and can be found in the anterior pituitary. Leptin antigens and mRNA were studied in the anterior pituitary (AP) cells of male and female rats to learn more about its regulation. Leptin antigens were found in over 40% of cells in diestrous or proestrous female rats and in male rats. Lower percentages of AP cells were seen in the estrous population (21 ± 7%). During peak expression of antigens, co-expression of leptin and growth hormone (GH) was found in 27 ± 4% of AP cells. Affinity cytochemistry studies detected 24 ± 3% of AP cells with leptin proteins and growth hormone releasing hormone (GHRH) receptors. These data suggested that somatotropes were a significant source of leptin. To test regulatory factors, estrous and diestrous AP populations were treated with estrogen (100 pM) and/or GHRH (2 nM) to learn if either would increase leptin expression in GH cells. To rule out the possibility that the immunoreactive leptin was bound to receptors in somatotropes, leptin mRNA was also detected by non-radioactive in situ hybridization in this group of cells. In estrous female rats, 39 ± 0.9% of AP cells expressed leptin mRNA, indicating that the potential for leptin production was greater than predicted from the immunolabeling. Estrogen and GHRH together (but not alone) increased percentages of cells with leptin protein (41 ± 9%) or mRNA (57 ± 5%). Estrogen and GHRH also increased the percentages of AP cells that co-express leptin mRNA and GH antigens from 20 ± 2% of AP cells to 37 ± 5%. Although the significance of leptin in GH cells is not understood, it is clearly increased after stimulation with GHRH and estrogen. Because GH cells also have leptin receptors, this AP leptin may be an autocrine or paracrine regulator of pituitary cell function.
I
Leptin levels are a natural signal for the hypothalamus to decrease appetite and regulate fuel homeostasis (Rowland et al. 1996; Vasselli 2001), which is how it got its name. [In Greek, “leptos” means thin (Imagawa et al. 1998).] However, leptin has recently been recognized as a multifunctional paracrine regulator in a number of organs, including the anterior pituitary (AP) (Yu et al. 1997; Jin et al. 1999, 2000; Morash et al. 1999; Lin et al. 2000; Vidal et al. 2000; Korbonits et al. 2001; Lloyd et al. 2001; Popovic et al. 2001; Pralong and Gaillard 2001; Sone and Osamura 2001; Sone et al. 2001).
A critical level of fat deposition is vital in most species for normal reproduction. It is widely believed that leptin (as a product of adipocytes) is an important signal to the body that there are sufficient fat stores to support reproduction. Recent reviews indicate that leptin is one of a number of permissive factors that allow the onset of puberty (Urbanski 2001; Mann and Plant 2002). Animals without leptin receptors do not go through puberty (Popovic et al. 2001). Animals without leptin reproduce poorly (Isozaki et al. 1999), and mice that overexpress leptin go through accelerated puberty (Yura et al. 2000).
One way that leptin might influence reproduction is via the regulation of AP gonadotropes, which have leptin receptors (Iqbal et al. 2000). Leptin stimulates gonadotropin release from pituitary gonadotropes, either by direct actions on AP cells or by stimulating gonadotropin-releasing hormone (GnRH) (Yu et al. 1997; Yura et al. 2000; Barb et al. 2001; Caprio et al. 2001; Ogura et al. 2001; Spicer 2001; Borowiec et al. 2002). Leptin receptors have also been found on corticotropes and somatotropes, suggesting that it may affect these types as well (Iqbal et al. 2000; Sone et al. 2001). Both inhibitory and stimulatory effects have been reported on somatotropes (Simard et al. 1986; Shimon et al. 1998; Tannenbaum et al. 1998; Vuagnat et al. 1998; Pombo et al. 1999; Isozaki et al. 1999; Barb et al. 2001; Chen et al. 2001; Roh et al. 2001; Marzullo et al. 2002).
Leptin has been considered a local regulatory factor because some AP cells contain leptin antigens. However, studies of rodents have reported different results as to the cell type. One set of studies of rats and mice found leptin only in TSH cells (Jin et al. 2000). Another group suggested that leptin was in gonadotropes or thyrotropes but not in somatotropes (Sone et al. 2001). Iqbal et al. (2000) reported leptin receptors in 70% of somatotropes in ovine pituitaries. Sone et al. (2001) showed the leptin receptor in 97% of somatotropes (but not the other cell types). They suggested that leptin might be produced by gonadotropes for paracrine regulation of somatotropes.
In humans, leptin proteins are found in 70% of corticotropes, 21% of somatotropes, and 30% of gonadotropes, prolactin cells, or thyrotropes (Popovic et al. 2001). In human pituitary adenomas, leptin mRNA was detected in subsets of all tumors but not in normal pituitaries (Korbonits et al. 2001).
This study was motivated by several factors. First, it is unclear if the pituitary leptin represents a significant source of the circulating hormone, because the cells are normally bathed in serum leptin from fat cells. However, it could be significant locally if it is tightly regulated by neuroendocrine mechanisms. Therefore, we were curious about potential regulators and changes in expression of AP leptin. Second, the lack of agreement about leptin production sites in the rodent AP could reflect species or tissue preparation differences. Third, leptin proteins could be in a cellular site as a result of binding to leptin receptors. To test these possibilities, we developed both immunolabeling and in situ hybridization (ISH) protocols that would detect co-expression of leptin in cells with somatotropic phenotypes. Once we identified leptin in somatotropes, we began tests of differential and regulated expression of pituitary leptin. GH cells were identified by their expression of GH antigens, mRNA, and GHRH receptors (Childs et al. 1999, 2000). These studies show evidence for changes in leptin expression with the estrous cycle. These studies also indicate that somatotropes have the potential to provide a source of leptin that is regulated by factors that stimulate GH cells.
Materials and Methods
Animals
Female Sprague-Dawley rats (weight 200–250 g) obtained from Harlan Sprague Dawley (Indianapolis, IN) were used throughout this study. Animals were housed three to four per cage with a 12-hr light-dark cycle maintained (lights on at 0600) at a constant room temperature of 68C. A standard pellet chow diet (Harlan Teklad-Rodent diet #8640; Madison, WI) and water were available ad libitum. Animals were allowed to become acclimated for approximately 2 weeks before vaginal smears were started. Pituitaries were taken during the morning from different stages of the cycle after the rats had completed at least two estrous cycles until we had collected at least five rats/stage. Rats of similar weight were taken when they had reached estrus or diestrus. They were anesthetized with IP injection of sodium pentobarbital (25 mg/kg or 0.05 ml/250 g rat) and then sacrificed by guillotine. The animal care protocol was approved, annually, by the Animal Use and Care Protocol Committee.
Dispersion of Pituitary Cells
Pituitaries from male rats and from female rats in each stage of the cycle (at 1000 hr) were rapidly removed and dispersed into single suspensions by the following protocol. After gently teasing away and discarding the neurointermediate lobe (by lifting it at the cleft), the remaining APs were chopped into small pieces and transferred to a 15-ml tube that contained DME and then centrifuged for 1 min at 400 rpm. The media were removed and the AP cells resuspended in DME containing trypsin (15 mg/4 ml) and DNase (10 μl of stock 1 mg/4 ml). The cell suspension was shaken gently on a flat shaker for 20 min and then centrifuged at 900 rpm for 5 min. The cells were placed in 4 ml of dispersion solution containing trypsin inhibitor (100 μl of stock 5 mg/ml DME; Sigma, St Louis, MO) and DNase (100 μl of stock 1 mg/ml; Sigma). A 3-ml syringe with an 18G needle was used to provide shearing force that would gently break up the pellets and disperse the cells. Cells were then resuspended in DME medium containing additives (ITS liquid media supplement 100X; 1 ml/100 ml of DME; Sigma). The cells were plated by pipeting 20 μl of cell suspension into each of the polylysine-coated coverslips and were allowed to adhere for 1 hr (providing 40,000–50,000 cells/well). Additional DME plus additives were added for an incubation of 15–48 hr in a CO2 incubator at 37C.
Stimulation of Pituitary Cells
The increase in endogenous AP leptin expression during diestrus suggested that it might be regulated, perhaps by estrogen. Furthermore, the fact that some leptin was found in GH cells suggested that GHRH may also regulate expression. Therefore, additional cultures of estrous rat pituitary cells were collected and half were stimulated overnight with 100 pM estrogen (Sigma; water soluble). Estrus was chosen because of its relatively low percentage of leptin-bearing cells. If either GHRH and/or estrogen were going to increase leptin expression, we reasoned that estrus might be an important nadir point. The experiment was therefore designed to learn if GHRH and/or estrogen would increase leptin expression, bringing it to levels seen during diestrus or proestrus. The following day, half of the cultures received 2 nM GHRH for 3 hr. Cells were fixed and double labeled for GH and leptin. Additional cultures pretreated with estrogen only were stimulated for 10 min with biotinylated GHRH and then dual labeled for the Biotinylated GHRH and leptin (Childs et al. 1999). Media were collected and frozen for leptin RIA, which was run using a commercial kit and kit instructions (RIA was run in the laboratory of Dr. Phil Kern, Dept. of Internal Medicine, UAMS).
Immunocytochemistry
Before immunocytochemical staining, the cells were fixed with 2.5% glutaraldehyde diluted in 0.1 M of phosphate buffer for 30 min at room temperature (RT). Fixation was followed by washes with 0.1 M phosphate buffer containing 4.5% sucrose and glycine 4× for 15 min.
Single Labeling for Leptin. One-day monolayer cells were pretreated with a 3% hydrogen peroxide solution for 5 min. After bleaching, the cells were washed with a 0.05 M Tris buffer three times. The cells were blocked with 0.05 M Tris diluent containing 1% BSA + 5% normal goat serum (NGS). The cells were then incubated in the primary antibody to rat/mouse leptin (Sigma) diluted 1:10,000 (in the blocking solution) for 30 min at 37C and washed with 0.05 M Tris buffer three times (note: the anti-leptin from NIH Hormone distribution office gave comparable results). Cells were then treated with a 1:80 dilution of biotinylated goat anti-rabbit IgG (Bio-Ganti; Vector Laboratories, Burlingame, CA). The Bio-Ganti was diluted in Tris diluent buffer solution and cells were incubated in it for 30 min at RT. Each of the wells were washed three times after application of Bio-Ganti. The cells were then exposed to 1:200 streptavidin (DAKO; Carpinteria, CA) for 30 min at RT. Then the cells were washed twice with Tris buffer and twice with sodium acetate. The peroxidase was detected with nickel-intensified diaminobenzidine (Ni-DAB) solution. One tablet of DAB (Sigma) was dissolved in 30 ml of acetate buffer (pH 6.0) that contained 0.45 g nickel ammonium sulfate. It was used within 1 hr. Just before use, 20 μl of 30% H2O2 was added and the solution was filtered immediately through a Whatman Grade I filter paper and applied to the cells for 6 min. Cells were washed three times with acetate buffer, dehydrated, and mounted on a slide.
Dual Labeling for GH and Leptin Antigens. After the labeling for leptin was established, the investigation focused on its expression by GH cells. For dual labeling, GH was first detected as previously described with anti-rat GH (NIH Hormone Distribution Program; 1:30,000 dilution). Nickel-intensified DAB was the reaction product (blue-gray to black). The coverslips were then labeled for leptin with techniques described above. However, the reaction product was DAB alone, which was orange/amber. Thus, we labeled for GH using a 1:30,000 dilution of the anti-GH and a 1:10,000 dilution of anti-leptin and Ni-DAB. Specificity tests for the GH have been described previously (Childs et al. 1999).
Validation of the Labeling Protocol. Method controls involved the omission of anti-leptin or the second antibody (Bio-Ganti). No labeling was seen in either method control; these were repeated multiple times as the labeling protocol was developed. Sensitivity tests used anti-rat leptin (Sigma) titrated from 1:1000 to 1:30,000. The labeling was gradually reduced with the concentration of anti-leptin and eliminated entirely at concentrations greater than 1:25,000. Optimal labeling that allowed detection of cells with different storage levels (a range of intensities) was seen after incubation in 1:10,000 for 2 hr at 37C. Therefore, this was used as the working dilution. Finally, to test specificity of the labeling for leptin-bearing cells, absorption tests with leptin were carried out, testing a final concentration of 1–10 μg/ml of recombinant rat leptin (L-5037; Sigma) in the 1:10,000 leptin antiserum. Initial tests showed that unabsorbed diluted anti-leptin lost its immunoreactivity in an overnight incubation, even at 4C. After tests showed that unabsorbed, diluted anti-leptin retained its reactivity for up to 4 hr, parallel tubes of absorbed and unabsorbed anti-leptin were incubated for 4 hr at 37C. Under these conditions, absorption with 10 μg/ml of leptin reduced labeling significantly in all cells.
Affinity Cytochemistry Detection of GHRH-receptive Cells
Cells were grown in defined medium overnight and stimulated the next day with 1 nM biotinylated GHRH. This analogue is biologically active and as potent as the native GHRH. Specificity tests for the affinity cytochemical detection method have been described previously (Childs et al. 1999). Stimulation for 10 min allows maximal labeling of all target cells. After this period, the cells were fixed and prepared for the detection of the biotinylated analogues with avidin-biotin-peroxidase complex (Childs et al. 1999). Control cultures containing vehicle only detected no labeling for Bio-GHRH.
In Situ Hybridization
The protocol used for ISH for leptin mRNA has been described in previous publications, including two recent techniques papers (Childs 1996, 1999; Childs et al. 2000). The probe for leptin was a 48-base pair (bp) oligonucleotide synthesized with 10 optimally spaced biotins (GreenStar; GeneDetect.com)/molecule. It is complementary to nucleotides 342–389 located within the coding sequence of the gene (accession number D49653). The biotinylated sense oligonucleotide sequence was also produced by GeneDetect. com and used as a control probe. Optimal labeling was detected by analysis of the intensity and number of labeled cells. Optimal labeling resulted in maximal numbers of cells without creating background. It was achieved on glutaraldehyde-fixed cells with 200 ng/ml of leptin probe. The same amount of sense leptin probe produced no labeling.
The pituitary cells were fixed in 2% glutaraldehyde as described previously, diluted in 0.1 M phosphate buffer plus 4.5% sucrose. They were then stored for use in the ISH protocol for no longer than 10 days. Prehybridization steps and the prehybridization buffer were as described previously (Childs 1996, 1999). The cells were then incubated in 200 ng/ml biotinylated leptin probe overnight (12–24 hr) at 41.2C. The biotinylated probe was then detected by a monoclonal antibody to biotin that was amplified by biotinylated anti-mouse IgG (Vector; rat absorbed) and streptavidin-peroxidase. The peroxidase was detected by Ni-DAB. This produced a sensitive blue-black reaction in patches or linear patterns in the cell. In parallel experiments, additional groups of cells were labeled for the leptin mRNA after which the population was prepared for dual labeling for GH antigens. The immunocytochemical technique was identical to that described above. It produced an orange/amber reaction product that usually filled the entire cytoplasm.
Statistical Analysis
Fields that were single labeled were counted to determine the percentages of leptin-containing cells. The first 200 cells encountered in a random scan of the field on one coverslip were analyzed. There were three coverslips/data point and three replicates. Leptin-bearing cells were dark gray-black-lighter gray and the unlabeled cells were clear and colorless. The leptin mRNA was detected in gray-black patches or linear patterns. Each experiment collected estrous or diestrous cells from at least two rats, but the experiments were repeated so that at least five rats were used. The experiment included three coverslips/group (×200 cells/coverslip). The values from each experiment were averaged. Each experiment was replicated three times and the final n represents the number average of the three replicates. One-way ANOVA was run to determine if there were any differences between experimental groups. If there were differences of p≤0.05, then Fisher's least significant differences test was run to identify means that were different.
Results
Immunolabeling for Leptin
Figure 1 compares percentages of pituitary cells with leptin antigens in groups of five to seven male rats or female rats in different stages of the estrous cycle. Male rats have 48% leptin-bearing cells. In diestrous female rats, 39 ± 8% of pituitary cells expressed leptin compared with 21 ± 7% of pituitary cells from estrous female rats (all values ± SEM unless otherwise noted). As animals approach proestrus, the leptin-bearing cells represented 46% of pituitary cells. These percentages are not different from values in the male, or those in the diestrous females. Figure 2 illustrates a field from a proestrous female rat immunolabeled for leptin with 1:10,000 anti-leptin. Figure 2 also shows the results of the specificity tests in which 10 μg/ml leptin neutralized labeling with the anti-leptin.
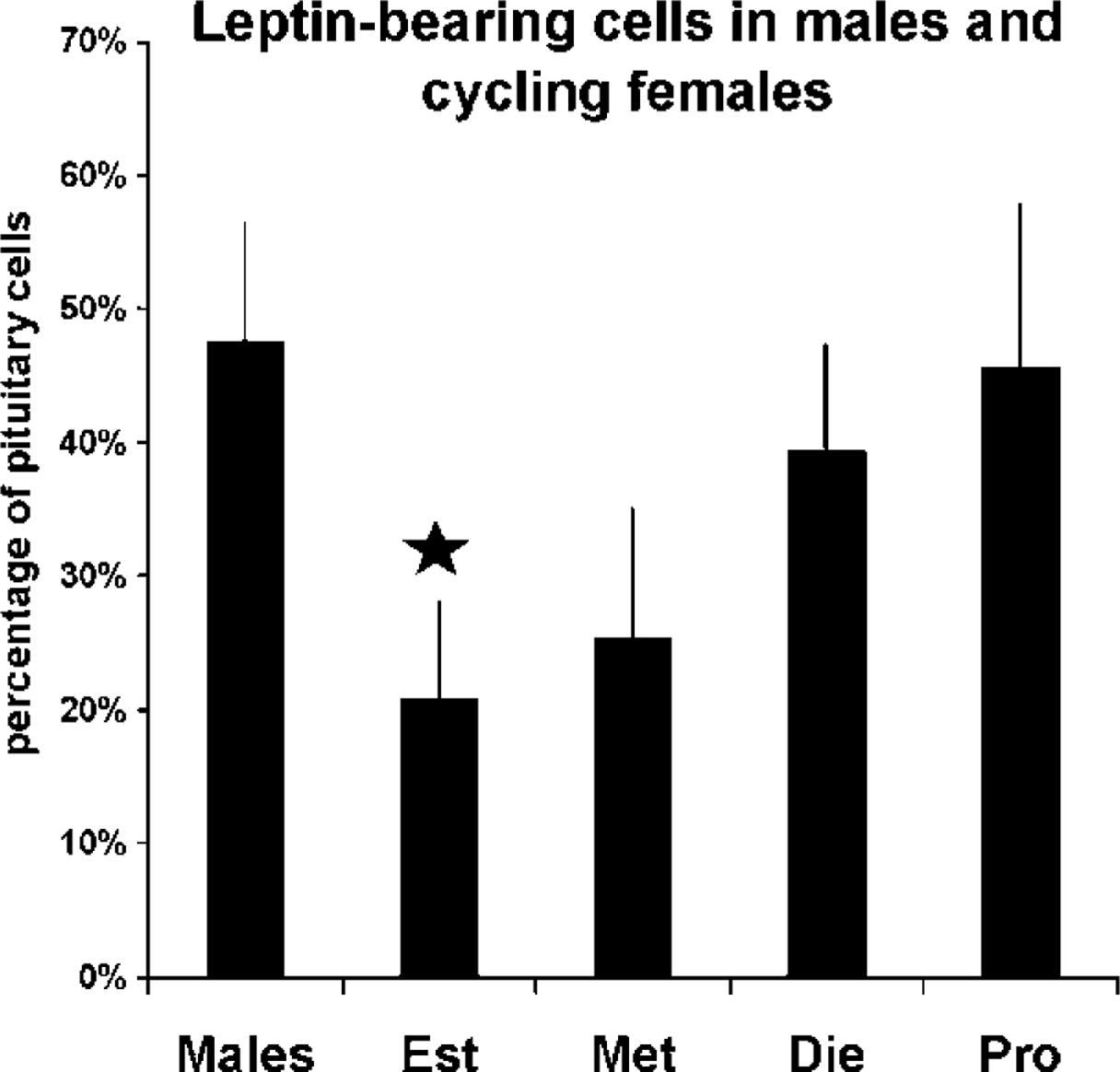
Counts of cells labeled for leptin in 1-day pituitary cultures from male rats or female rats taken in the morning of different stages of the estrous cycle. Values are average ± SEM (n = 3 experiments). The lowest value (different from all other values) is that in the estrous population (star).
Leptin Expression in GH Cells
Double-labeled cells were initially counted in diestrous animals because this was a peak time of production for leptin. In these populations, 27 ± 4% of pituitary cells co-expressed GH and leptin (values ± SD), which is about 60% of the total population of leptin-bearing cells. These values could be seen in cultures taken after 24 or 48 hr of incubation. The analysis showed that 92 ± 2% of GH cells (values ± SD) contained leptin antigens suggesting that, in the diestrous rat population, most GH cells express this peptide. Figure 3 shows double labeling for GH and leptin in these cultures. It also shows that some of the labeled cells are leptin-rich, i.e., they contain very little GH. Others have more GH.
Leptin Expression in GHRH Target Cells
Detection of GHRH binding provided a second phenotypic marker for leptin-bearing somatotropes. Quantification of dual labeling for Bio-GHRH and leptin showed that, in diestrous animals, 24 ± 3% (values ± SD) of pituitary cells bound Bio-GHRH and also stored leptin. This is illustrated in Figure 4. This co-expression involved 84 ± 10% (values ± SD) of biotinylated GHRH target cells. Analysis of populations from estrous rats (in which leptin was found in only 21% of pituitary cells), showed that 11 ± 1.7% of pituitary cells co-expressed bio-GHRH binding sites and leptin. Thus, about half of the GHRH-receptive cells stored leptin in the estrous animals. The patches of peripheral labeling could be on the surface, or in vesicles and early endosomes. This distribution is typical of that seen after a 10-min pulse of biotinylated GHRH.
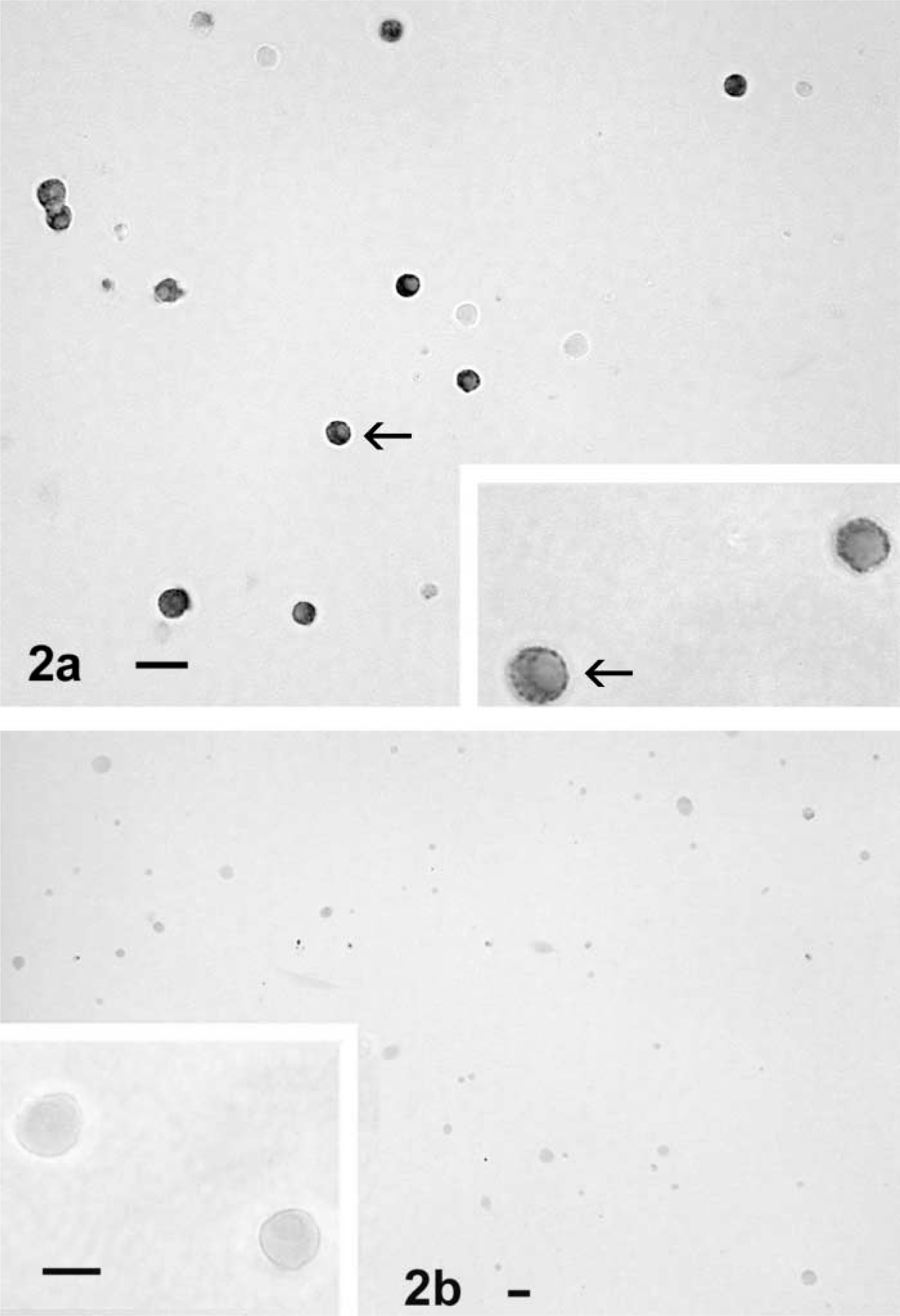
Labeling for leptin in 1-day pituitary cultures from proestrous female rats. (
Estrogen and GHRH Regulation of Leptin Expression
As stated previously, estrous rats were used for these experiments because they contained the lowest percentages of leptin protein-bearing cells. Figure 5 shows the changes in leptin expression in estrous rats induced by estrogen plus GHRH. Neither estradiol (100 pM) overnight nor 2 nM GHRH alone for 3 hr stimulated expression of more leptin-bearing cells. However, stimulation with both estrogen and GHRH increased expression of leptin to levels similar to those found in diestrous or proestrous animals (41 ± 9%; see Figure 1).
RIA data on media leptin after 3 hr of GHRH (with or without estradiol) showed no detectable leptin proteins in the media and no effects of GHRH or estradiol on media leptin. Figure 6 illustrates the vehicle-treated control, showing the few scattered leptin-bearing cells in estrous rat cultures. Figure 7 shows the increase in numbers of leptin-bearing cells along with an increase in the density of label in the cells. In addition, there were scattered enlarged stellate cells that labeled for leptin in these GHRH-estradiol-treated cultures. Therefore, whereas GHRH and estrogen stimulated more leptin-bearing cells, they did not stimulate secretion of leptin, as detected by RIA.
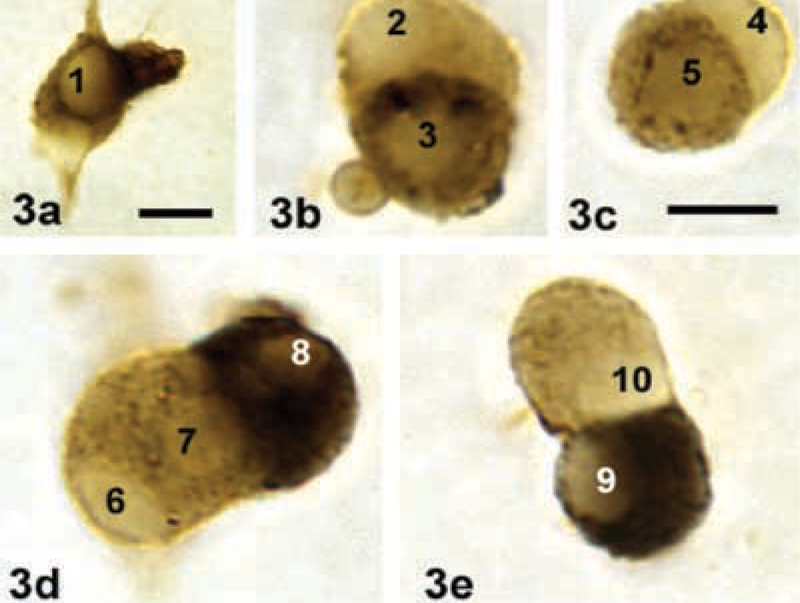
(
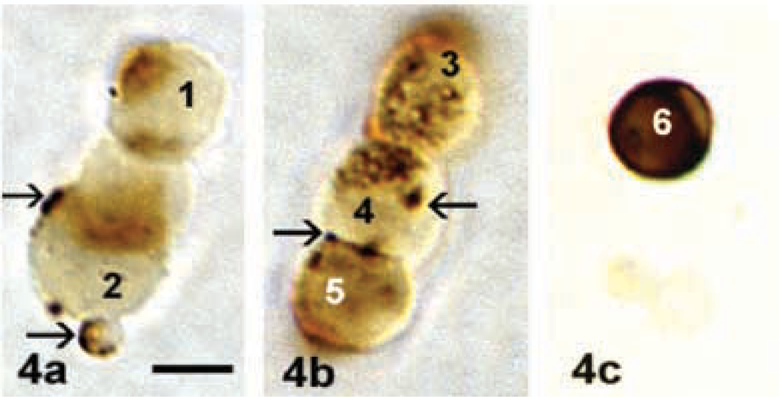
(
Detection of Leptin mRNA in Somatotropes
The fact that leptin proteins were not detected in the media suggested that either leptin was an autocrine or paracrine mediator or that the leptin found in the somatotropes was bound to receptors. Therefore, to learn more about the leptin being detected, the study was expanded to include the detection of leptin mRNA in populations from five additional estrous female rats. Only ISH would allow us to differentiate a binding site from a site of production. Second, this expansion would enable us to learn if GHRH and estrogen upregulated leptin at the level of expression of leptin mRNA.
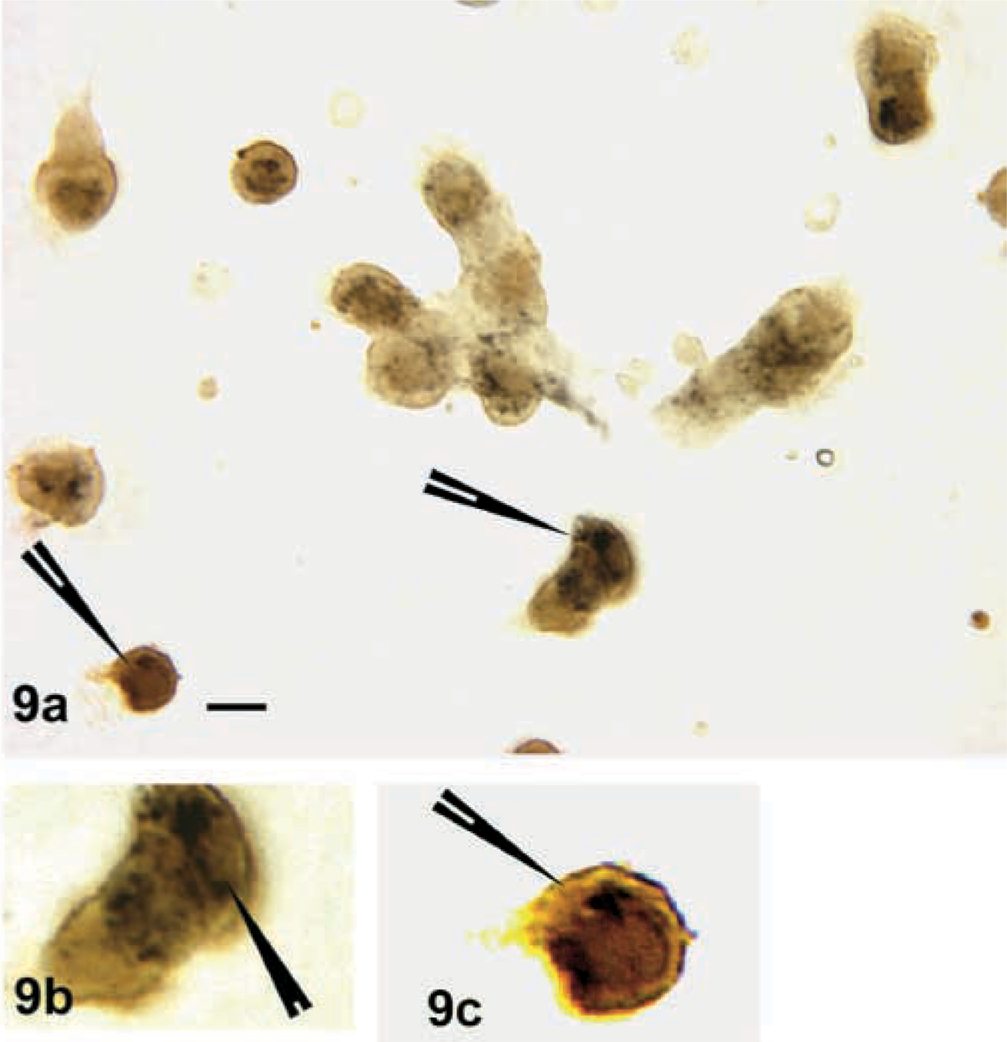
Counts of leptin mRNA-expressing cells in diestrous or estrous populations showed that 39 ± 0.8% of AP cells expressed leptin mRNA; 20 ± 2% of AP cells expressed leptin mRNA and GH antigens. This is not significantly different from percentages of cells with leptin and GH proteins. After stimulation with GHRH and estrogen, there was a significant (p<0.003) increase in the percentage of cells with leptin mRNA (57 ± 4%) as well as those that produced leptin and GH (36 ± 5%). Figure 8 illustrates the dual labeling for leptin mRNA and GH antigens in cultures that were vehicle treated. Figure 9 shows the increase in cells with leptin and/or leptin and GH after treatment with estrogen and GHRH.
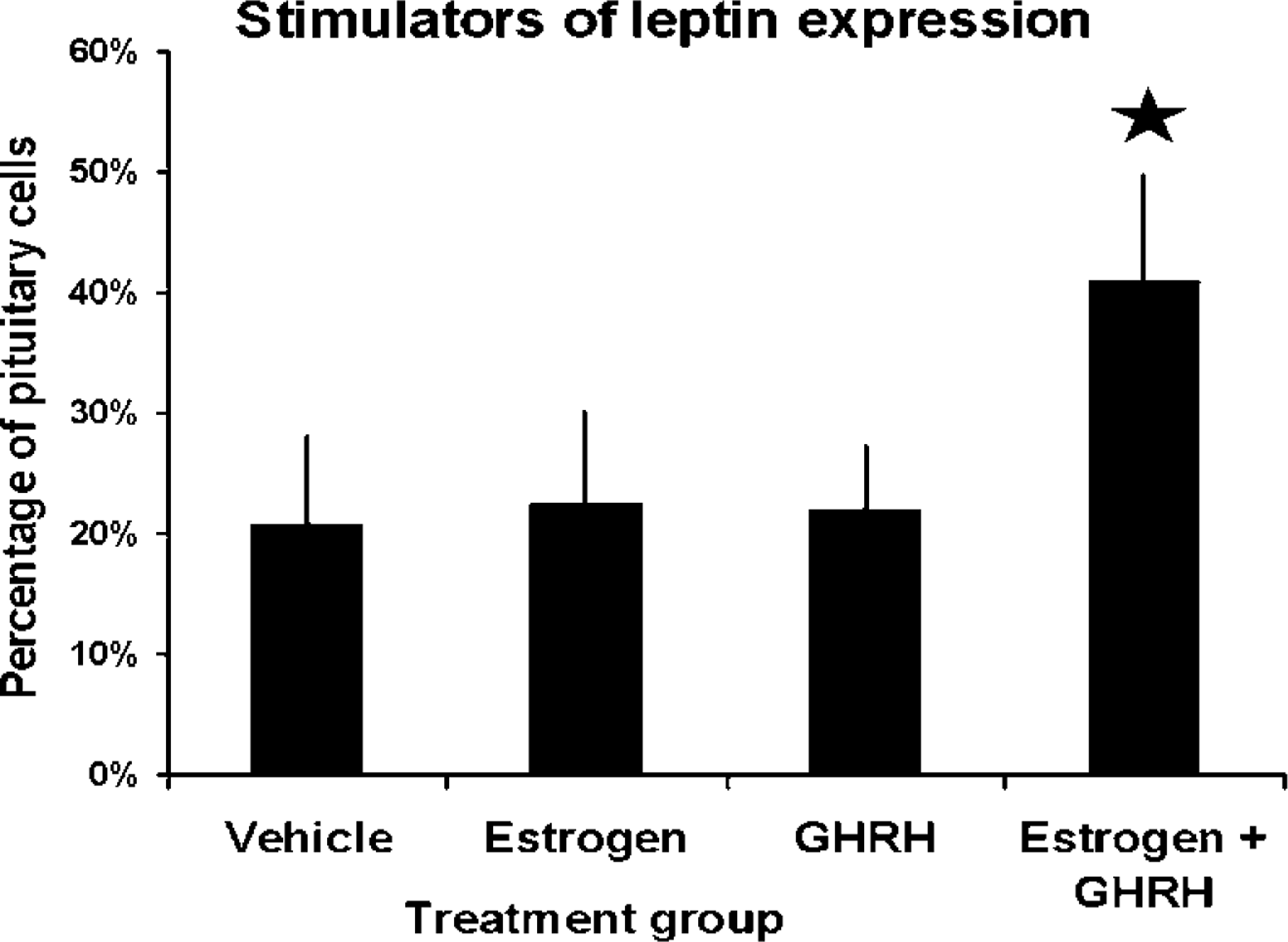
Counts of leptin-bearing cells after overnight stimulation with 100 pM estradiol and/or 3 hr in 2 nM GHRH. Only the combination of GHRH and estradiol increased the percentages of leptin-bearing cells to levels similar to those seen in diestrous rats. Values are averages ± SEM (n = 3 experiments). Star, significantly different from vehicle control.
Discussion
These in vitro studies show that leptin expression is found in 20–40% of pituitary cells in cycling female rats (depending on the stage of the cycle) and in 48% of AP cells in male rats. Furthermore, the populations with the lowest percentages can be induced to produce more leptin mRNA or antigens if these AP cells are exposed to estrogen and GHRH. The studies also show that many somatotropes co-express leptin proteins or mRNA, whether they are detected by their content of GH antigens or GHRH receptors. These studies agree in part with the studies of human pituitaries that showed that a subset of GH cells express leptin (Popovic et al. 2001). They do not confirm studies of rats or mice that show no leptin expression by GH cells (Sone et al. 2001). The differences may reflect the different preparation techniques. The present studies used glutaraldehyde as a fixative, known to stabilize polypeptides. Furthermore, we labeled whole cells that had not been dehydrated or embedded, thus preventing the deleterious effects of these reagents. Leptin may be labile and not as easily detected in routine tissue sections. Finally, the whole cells presented ample leptin mRNA and protein stores for detection, allowing identification of cells with lower storage levels.
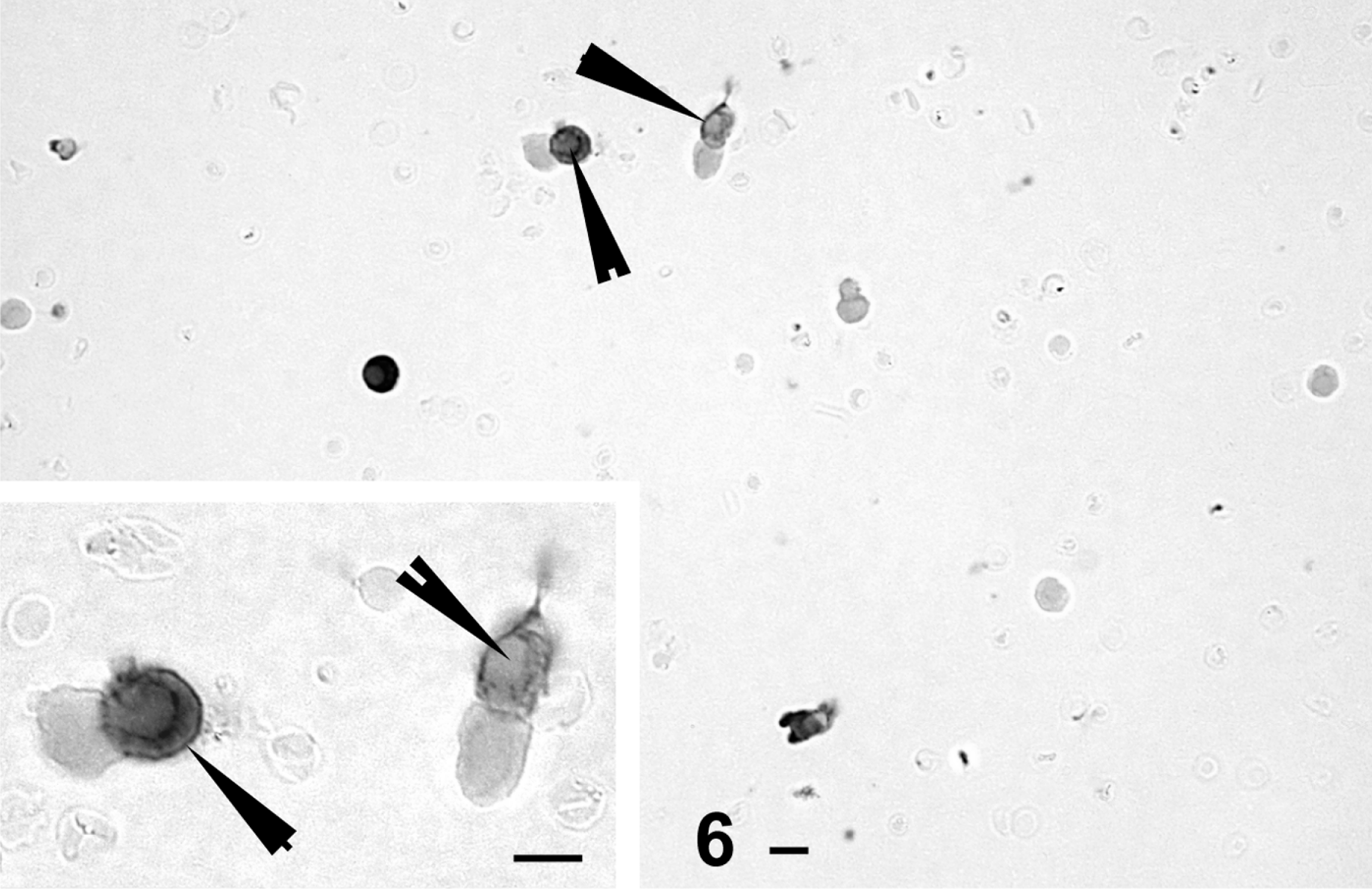
Labeling for leptin in cells from estrous female rats treated overnight with vehicle. Bar = 20 μm.
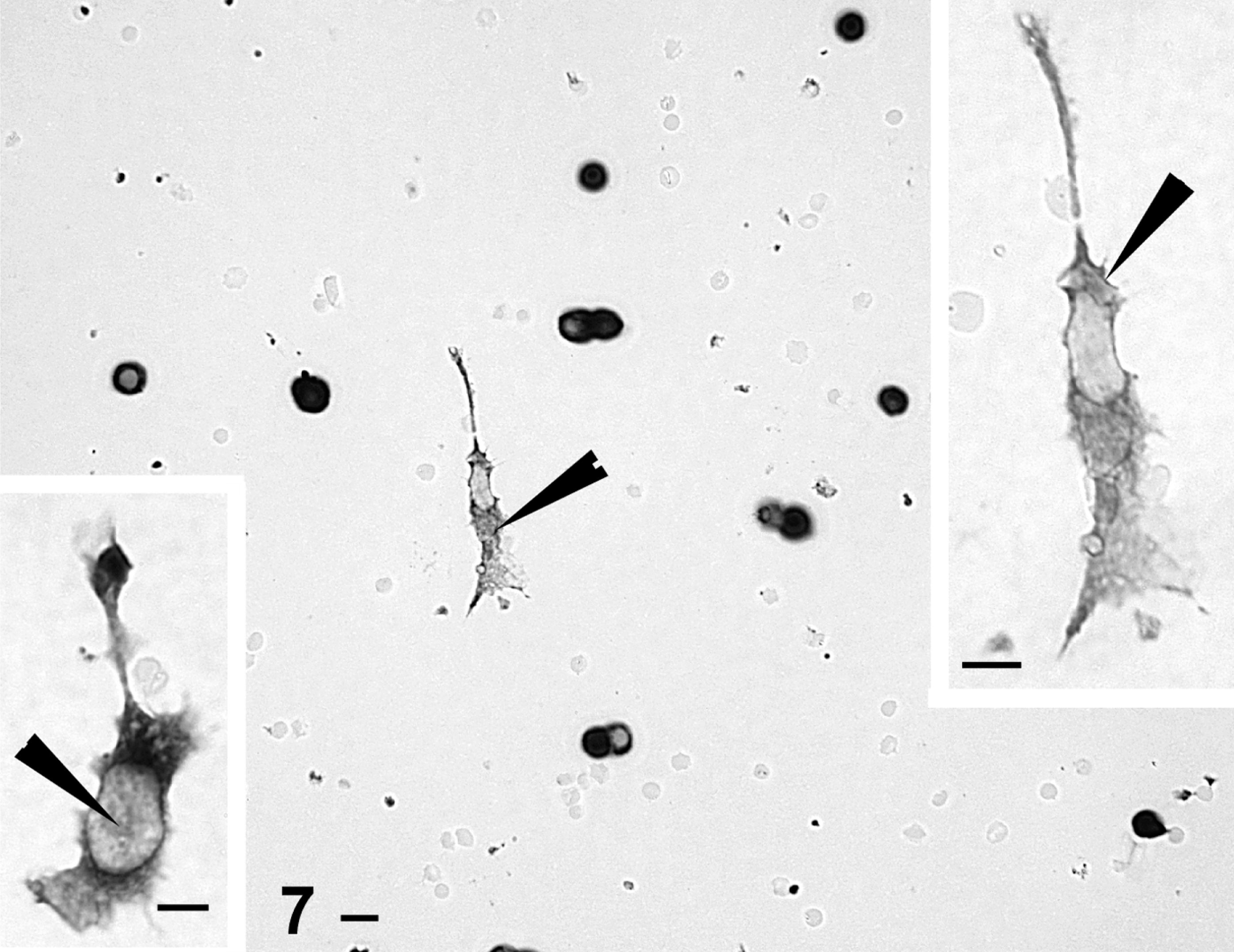
Leptin labeling in cells from estrous female rats treated with 100 pM estradiol followed by 2 nM GHRH for 3 hr before fixation. Significant increases bring the population to numbers similar to those in AP cells from diestrous females. In the estradiol-treated fields, more stellate leptin-bearing cells are evident (arrowheads). Bars = 20 μm.
We continue to recognize that it is possible that some of the leptin proteins are present in cells bound to leptin receptors. This would be particularly true for cells known to have leptin receptors (Iqbal et al. 2000; Sone et al. 2001). The studies were thereby strengthened by the detection of leptin mRNA. Protocols for the non-radioactive detection of leptin mRNA were developed and applied during the studies of leptin regulation. The studies showed that about 40% of the AP cells produce leptin mRNA; about half of these co-express GH antigens. The percentages of cells that co-express leptin mRNA and GH antigens are not significantly different from those that co-express leptin and GH proteins.
The dispersion technique was required to detect biotinylated GHRH on whole living cells in vitro (Childs et al. 1999). We recognize that the dispersion and brief culturing may have altered the cells, stimulating them to produce more leptin. However, if one compares populations of cells taken from 1 hr to those taken 48 hr after dispersion, the percentages of leptin-bearing cells do not change. This was also true when we compared expression of leptin mRNA in freshly dispersed cells with those grown overnight.
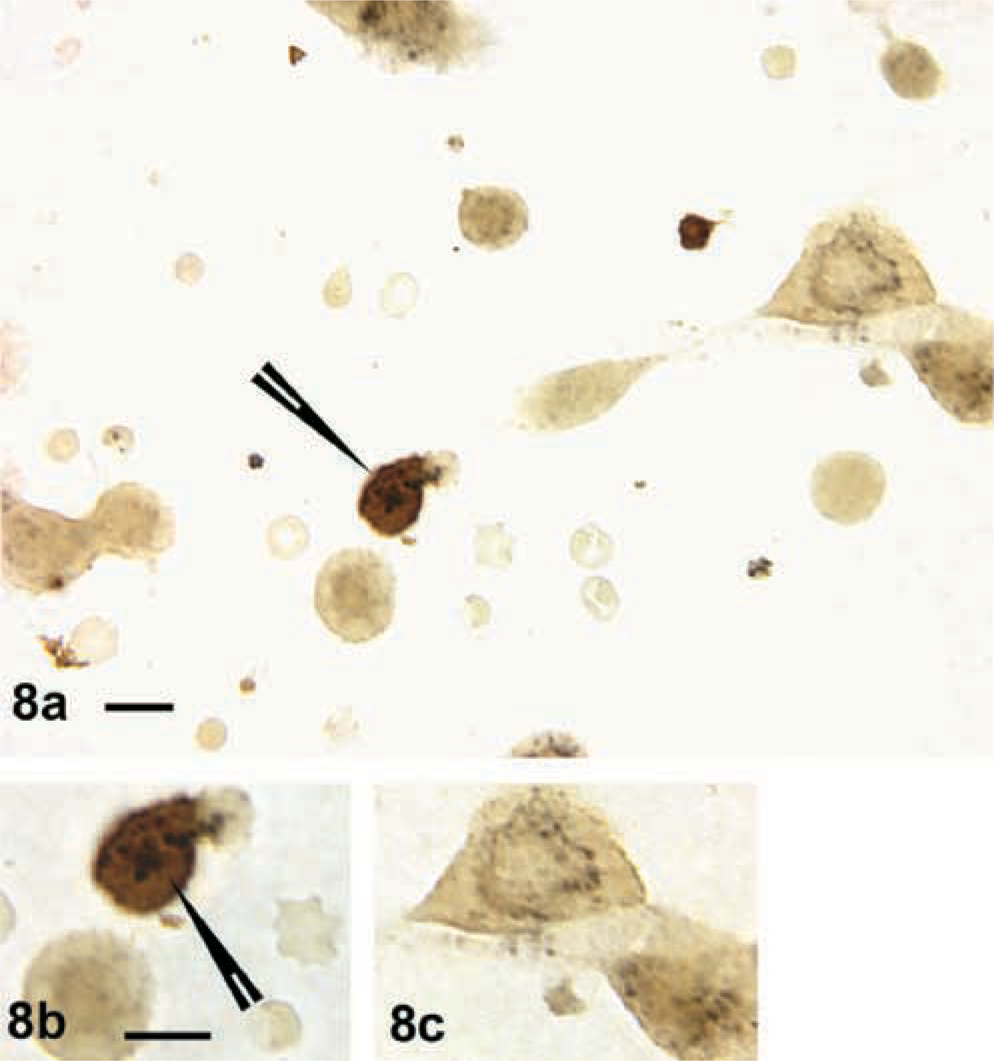
Dual labeling for leptin mRNA (dark gray-black) with biotinylated complementary oligonucleotide probe and GH antigens (orange/amber) in vehicle-treated AP cells from estrous rats. (
(
The ISH evidence indicates that most of the immunolabeling represents leptin that has been synthesized by GH cells. An analysis of leptin mRNA in the estrous rat population shows that 39% of the cells express the transcript. In contrast, the immunolabeling for the proteins shows a drop to only 20% of the population for that population. This suggests that leptin protein stores may be down in the estrous female AP cells. Clearly the leptin cells can still be identified by their content of mRNA.
Collectively, these data provide information about potential new mechanisms behind the somatotropic regulation of metabolic functions. Only estrogen and GHRH together enhanced GH cell labeling for leptin proteins or mRNA. This suggests that leptin expression is regulated in GH cells. Estrogen is a potent stimulator of GH cells (Webb et al. 1983; Simard et al. 1986; Silverman et al. 1988; Jin et al. 1989; Bethea 1991). We have recently reported that estrogen increases the overall percentages of GHRH-bearing cells (Childs and Unabia 2002). Therefore, estrogen may have increased the percentages of GHRH target cells during the overnight incubation. This would then potentiate the actions of GHRH seen the next day and may explain the additive regulatory effects.
The evidence thus far suggests that somatotropes represent a principal leptin-bearing cell in the AP. This does not rule out leptin production by other cell types because a number of studies by us and others have shown that somatotropes may co-express prolactin, gonadotropins, or TSH (Childs et al. 1994, 1999, 2000). Only 24% of pituitary cells from diestrous animals contain GH and leptin proteins, which is 61% of the leptin-containing cells in the diestrous rat pituitary. Hence, the remaining leptin-bearing cells could include gonadotropes or thyrotropes.
Finally, our analysis showed that only half of the leptin-bearing cells in estrous rats bind GHRH. In contrast, in diestrous rats there is an increase to 92% of leptin-bearing cells. Thus, leptin-bearing cells may be a subset of GHRH target cells under regulatory control of estrogen in vivo (rising estrogen is seen from estrus to diestrus) or in vitro.
Functional Significance of Estrogen and Leptin Regulation
The studies comparing differences in expression of leptin during the cycle correlate well with changes in serum leptin levels seen during the estrous cycle of rats (Tanaka et al. 2001). Tanaka et al. (2001) showed that serum leptin levels (presumably due to secretion from adipocytes) increase in proestrus and decrease during estrus. A reduction in body weight due to a decrease in fat intake in the proestrous rats has also been documented by this group. Therefore, leptin may limit food intake at this time.
However, it is widely known that additional energy is required to maintain reproduction and a new pregnancy. Because proestrus immediately precedes the time of ovulation (early morning of estrus), a decrease in leptin at that time might have a rebound effect and promote feeding and energy reserves. This could be under the regulation of progesterone, which is high during the time leptin is low. Therefore, the significance of these changes may be related to the need for appetite regulation to support and enhance reproduction (Martini et al. 1994; Friedman and Halass 1998; Ruhl and Everhart 2001; Tanaka et al. 2001). Tanaka et al. (2001) also reported that estrogen increased leptin production in adipocytes, which suggests that it might be a regulatory factor for systemic leptin. Their findings correlate well with our in vitro evidence showing that estrogen-treated cells from estrous animals responded to GHRH by increased leptin expression. However, our studies suggest that estrogen alone is not sufficient to increase leptin from GH cells. This may indicate the need for additional neuroendocrine modulators, such as GHRH.
Other Roles for Pituitary Leptin, an Autocrine Regulator of GH Cells
Roh et al. (2001) demonstrated that 60% of somatotrophs exhibited positive staining for leptin receptors. This suggests that leptin may regulate secretion from somatotropes. GH, in turn, is also a vital regulator of body fat composition and lean muscle mass. It interacts directly with adipocytes, is lipolytic, and reduces leptin production (Isozaki et al. 1999; Asada et al. 2000).
The regulation of leptin and GH may be closely linked because of their metabolic roles. In humans, GH increases during changes in nutritional status under various conditions such as starvation, food deprivation, and reduced blood sugar level. (Popovic et al. 2001; Ruhl and Everhart 2001). These are conditions in which leptin levels are lower. GH levels decrease during obesity and in states where nutrition is high; leptin levels are generally high during these states. Therefore, there may be an inverse relationship between serum GH and serum leptin, at least in humans. GH-deficient humans show higher leptin and increased adiposity, whereas acromegalics have reduced body fat and lower leptin (Asada et al. 2000; Marzullo et al. 2002).
This inverse relationship has led workers to propose that leptin inhibits GH secretion (Chen et al. 2001; Roh et al. 2001), and the hypothesis has been proved experimentally. Chen et al. (2001) reported that somatotropes treated with recombinant leptin for 3 days showed a decrease in GH responses to GHRH. However, other workers have shown stimulatory effects of leptin on GH (Simard et al. 1986). Therefore, the role of leptin in the regulation of GH secretion remains to be determined.
Leptin expression is increased in GH cells by estradiol and GHRH, both of which are regulators of somatotropes. The significance of this production site is unclear. Perhaps leptin is produced by GH cells to add to that produced by adipocytes. However there is little evidence for AP-mediated secretion of leptin in the present studies of secretion. The lack of evidence for secretion, coupled with the fact that there are leptin receptors on GH cells, raises the question of whether it might be an autocrine regulator (Iqbal et al. 2000; Sone et al. 2001).
Because the pituitary is bathed in leptin from the adipocytes, an additional source of AP leptin is difficult to explain. Future studies are needed to learn if leptin can be secreted from pituitary cells. One can speculate that a tightly regulated source (by the neuroendocrine system) that works locally in an autocrine or intracrine manner might provide transitional control of populations of cells as needed. Thus, as others have suggested, leptin may serve as a local cytokine mediator in the pituitary.
In conclusion, our results demonstrated leptin expression by 20–45% of AP cells in male and cycling female rats. Dual labeling detected leptin expression in cells with GH antigens or receptors for bio-GHRH. The detection of leptin proteins at sites of production was further confirmed by the detection of leptin mRNA. In addition, this study is the first to report potential regulatory mechanisms for leptin proteins and mRNA in the pituitary. The findings suggest that regulators such as estrogen and GHRH may be one of the triggers for these changes. The functional significance of pituitary leptin remains to be determined. However, the collective evidence suggests that it may be an autocrine or paracrine regulator.
Footnotes
Acknowledgements
This study was supported by NIH HD 33915, by a research assistantship from the VA, and by the Department of Nutrition (IM). It is now supported by NSF IBN 0240707 (GVC).
We wish to thank A.F. Parlow and the Hormone Distribution Program for the anti-rat GH antiserum. We also thank Mary Iruthayanathan and Diana Rougeau for help with the protocols.