
Abstract
In mammals, the oviduct is an important source of factors that play key roles in reproductive and developmental events. The major components of oviduct fluid are oviduct-specific glycoproteins, but other proteins are synthesized and secreted by the oviduct epithelium. Leptin and adiponectin are two hormones originally identified in adipocytes that play a critical role not only in the control of energy balance and metabolism but also in diverse functions such as reproduction. This study investigates the presence and distribution of leptin and adiponectin in the rat oviduct through a combination of immunohistochemistry and reverse transcription–polymerase chain reaction techniques. Using both techniques, it has been detected that the oviduct of cycling rats expresses leptin and adiponectin. Immuno-reactivity for both adipokines appears in the apical region of the secretory epithelial cells, only in the isthmus and ampulla. The immunostain is stronger in the isthmus and changes throughout the estrous cycle in the ampulla, increasing from proestrous to estrous. The results presented here are a further contribution to the identification of leptin and adiponectin produced and secreted by the oviduct epithelium, which must be taken into account for a better understanding of the reproductive events that take place in this organ.
Keywords
T
Oviduct fluid is composed of hundreds of macromolecules derived either by new secretion from oviduct epithelium or serum transudate (Beier 1974; Buhi et al. 2000; Leese et al. 2001; Buhi 2002). Proteins constitute a major component of oviduct secretion, many of them being similar to those in serum (Gerena and Killian 1990). The protein pattern includes many enzymes, protease inhibitors, growth factors, and a group of OGP or oviductins (Hunter 1988; Arias et al. 1994; Buhi 2002; Killian 2004). In this context, identification and characterization of other specific oviduct proteins and the assessment of their potential role in early reproductive events seems justified.
Reproduction in mammals is dependent on the availability of energy. In particular, it has been demonstrated that a minimum weight for height is necessary for normal sexual function in women (Frisch and McArthur 1974). Adipose tissue participates in the regulation of energy homeostasis, secreting a large number of biologically active adipokines (Frühbeck et al. 2001). Leptin and adiponectin are two of the best-characterized adipokines; both affect carbohydrate and lipid metabolism and are associated with increased insulin sensitivity (Frühbeck et al. 2001; Guerre-Millo 2002; Ahima 2005; Mitchell et al. 2005). These hormones have a broad pattern of expression in the organism and are implicated in many different functions, including reproduction. In this context, special attention has been paid in recent years to analysis of the link between reproductive function and molecules involved in energy homeostasis, such as adipocyte-derived hormones (Caprio et al. 2001; Guerre-Millo 2002; Tena-Sempere and Barreiro 2002; Mitchell et al. 2005).
List of antisera used in IHC
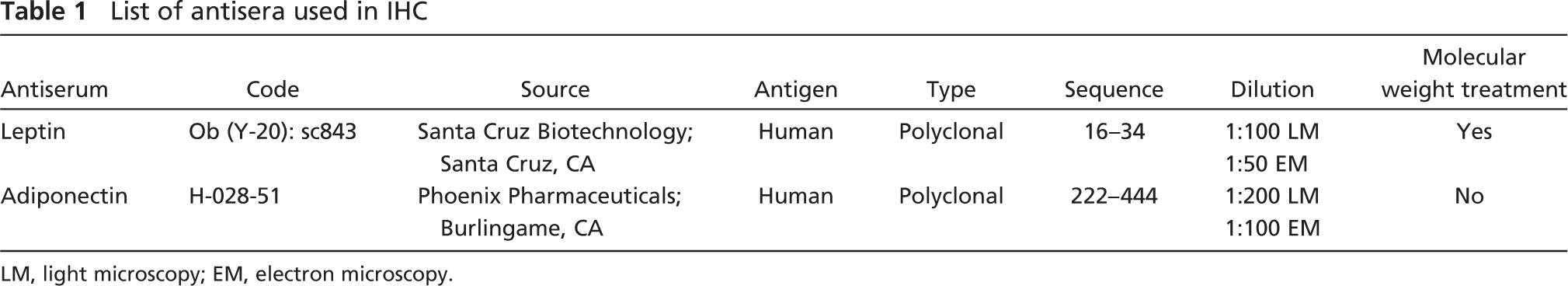
LM, light microscopy; EM, electron microscopy
Leptin is the product of the ob gene (Zhang et al. 1994). Circulating leptin concentrations are coupled to energy stores (Loftus 1999), increasing in the fed state and decreasing during fasting. Leptin is the signal that informs the brain about the degree of adiposity. The relationship between leptin and reproduction was first studied in sterile mice with a homozygous mutation of the ob gene (ob/ob mice) (Zhang et al. 1994). Leptin administration restored fertility in both female (Chehab et al. 1996) and male (Mounzih et al. 1997) ob/ob mice. Leptin and its receptor have been shown to be expressed in ovary (Karlsson et al. 1997; Spicer and Francisco 1997; Löffler et al. 2001; Ryan et al. 2002; Archanco et al. 2003) and testis (Hoggard et al. 1997; Caprio et al. 1999; Tena-Sempere et al. 1999), as well as in several other tissues of the reproductive tract. Identification of leptin in these organs pointed out its participation in the control of reproductive function in both sexes (Caprio et al. 2001; Tena-Sempere and Barreiro 2002). Adiponectin is also synthesized and secreted by adipocytes to the blood stream (Scherer et al. 1995; Berg et al. 2002). This hormone has also been documented in many different tissues of the organism such as the liver (Yoda-Murakami et al. 2001), skeletal muscle (Delaigle et al. 2004), bone tissue (Berner et al. 2004), and cardiac muscle (Pineiro et al. 2005). In addition to leptin, adiponectin has also been described as an important regulator of appetite and energy homeostasis, affecting insulin sensitivity (Hotta et al. 2001; Diez and Iglesias 2003; Heilbronn et al. 2003). In contrast to other adipokines, adiponectin levels are reduced in cases of obesity and increase in response to weight loss (Weyer et al. 1930). Involvement of this hormone in many other biological functions, including reproduction (Meier and Gressner 2004; Mitchell et al. 2005), has been put forward. Adiponectin plasma levels present sexual dimorphism in adult human and mice (Combs et al. 2004; Gui et al. 2004; Mitchell et al. 2005). Furthermore, some pathologies of the reproductive tract (estrogen-dependent diseases) such as endometriosis (Dal Maso et al. 2004; Takemura et al. 2006), breast cancer (Miyoshi et al. 2003; Mantzoros et al. 2004; Harvie and Howell 2006), and uterine leiomyoma (Chen et al. 2004) are associated with changes in serum adiponectin levels.
List of primers used in RT-PCR
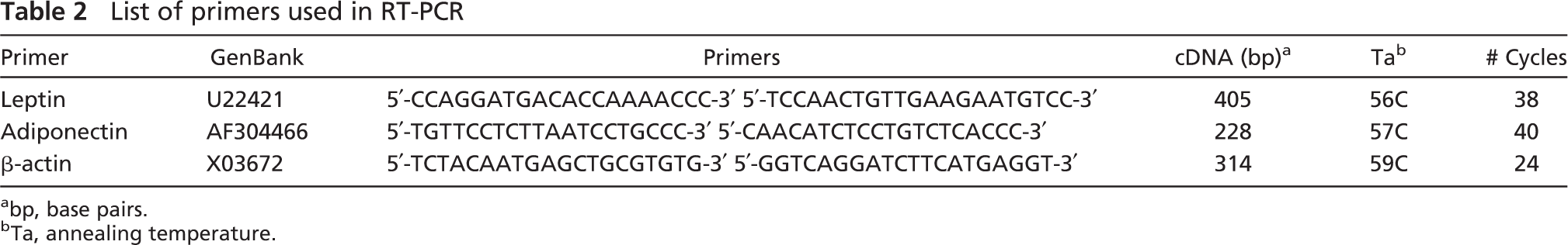
bp, base pairs.
Ta, annealing temperature.
In a previous study, expression of leptin in the rat ovary with a targeted distribution in oocytes and steroid-producing cells was examined (Archanco et al. 2003). Involvement of leptin in gamete maturation and early embryo development in mammals (Antczak and Van Blerkom 1997; Matsuoka et al. 1999; Kawamura et al. 2002, 2003), events that take place in the oviduct, have been proposed. Nonetheless, the pattern of expression of leptin or any other adipokine in the oviduct and their participation in oviduct functions remains to be completely disentangled. Likewise, expression of adiponectin in the reproductive tract and its potential role in reproduction need to be clarified.
To address these issues, detailed identification of leptin and adiponectin expression in the rat oviduct was carried out by a combination of immunohistochemistry (IHC) and reverse transcription–polymerase chain reaction (RT-PCR) techniques. Results obtained in this study provide further information about the implication of leptin and adiponectin in the female reproductive function.
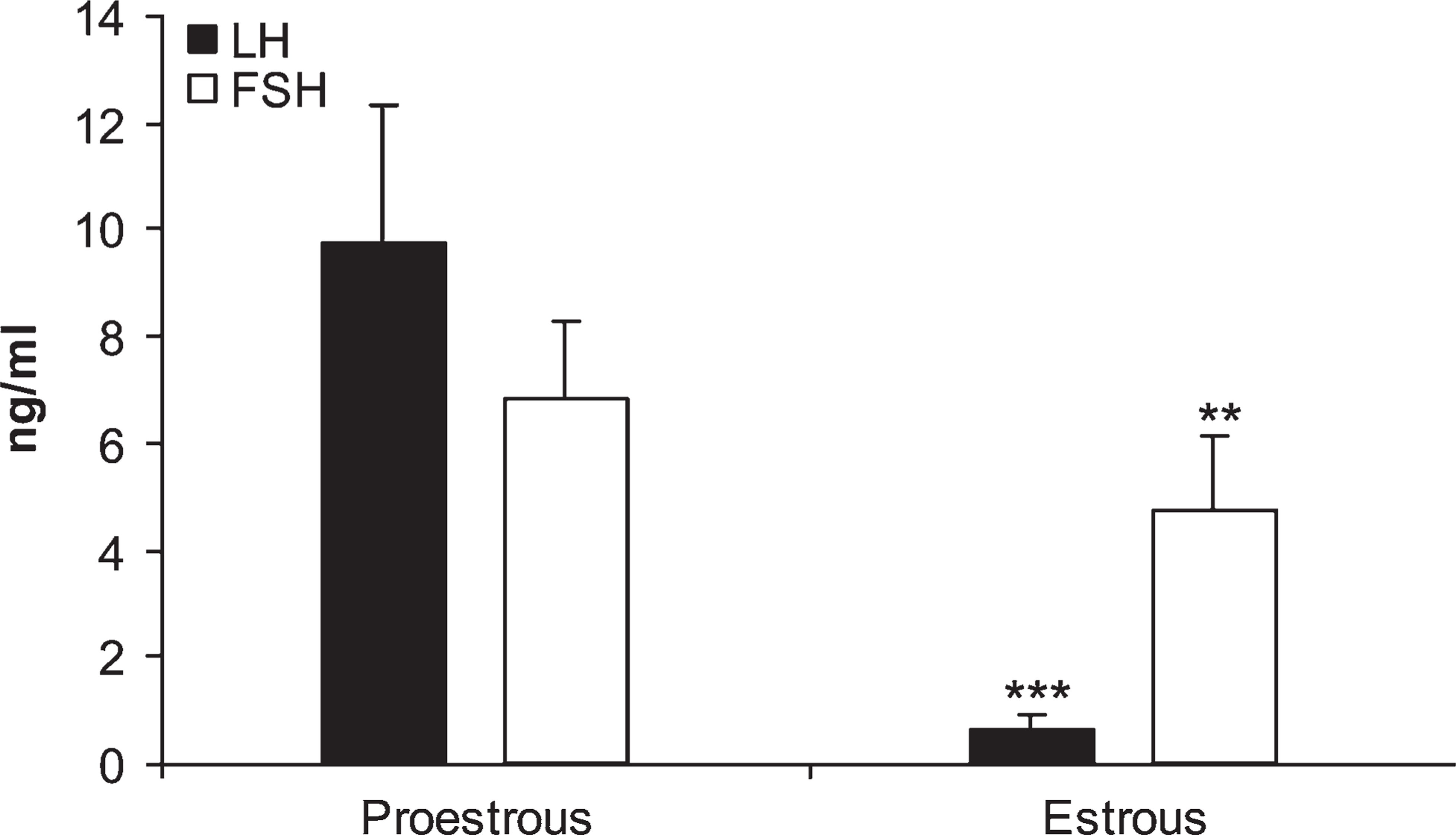
Histograms representing luteinizing hormone (LH) and follicle-stimulating hormone (FSH) concentrations in serum samples of rats in proestrous and estrous phases. Data are mean ± SEM of five rats. ∗∗
Materials and Methods
Animals
Ten Wistar rats, aged 12 weeks and weighing 230 ± 5 g, were used in this study. Animals were maintained under conventional conditions with 12:12 hr light/dark cycle (lights on at 8:00 AM) and at an ambient temperature of 21 ± 2C, with water and standard laboratory diet (Rodent Toxicology Diet; B and K Universal Ltd., Hull, UK) available ad libitum. Vaginal smears were collected daily (between 9:00 AM and 10:00 AM) to determine the correct development of the estrous cycle, and only rats with consistent 4-day cycles were used. All experimental procedures conformed to the European Guidelines for the Care and Use of Laboratory Animals. The study was approved by the Ethical Committee of the University of Navarra.
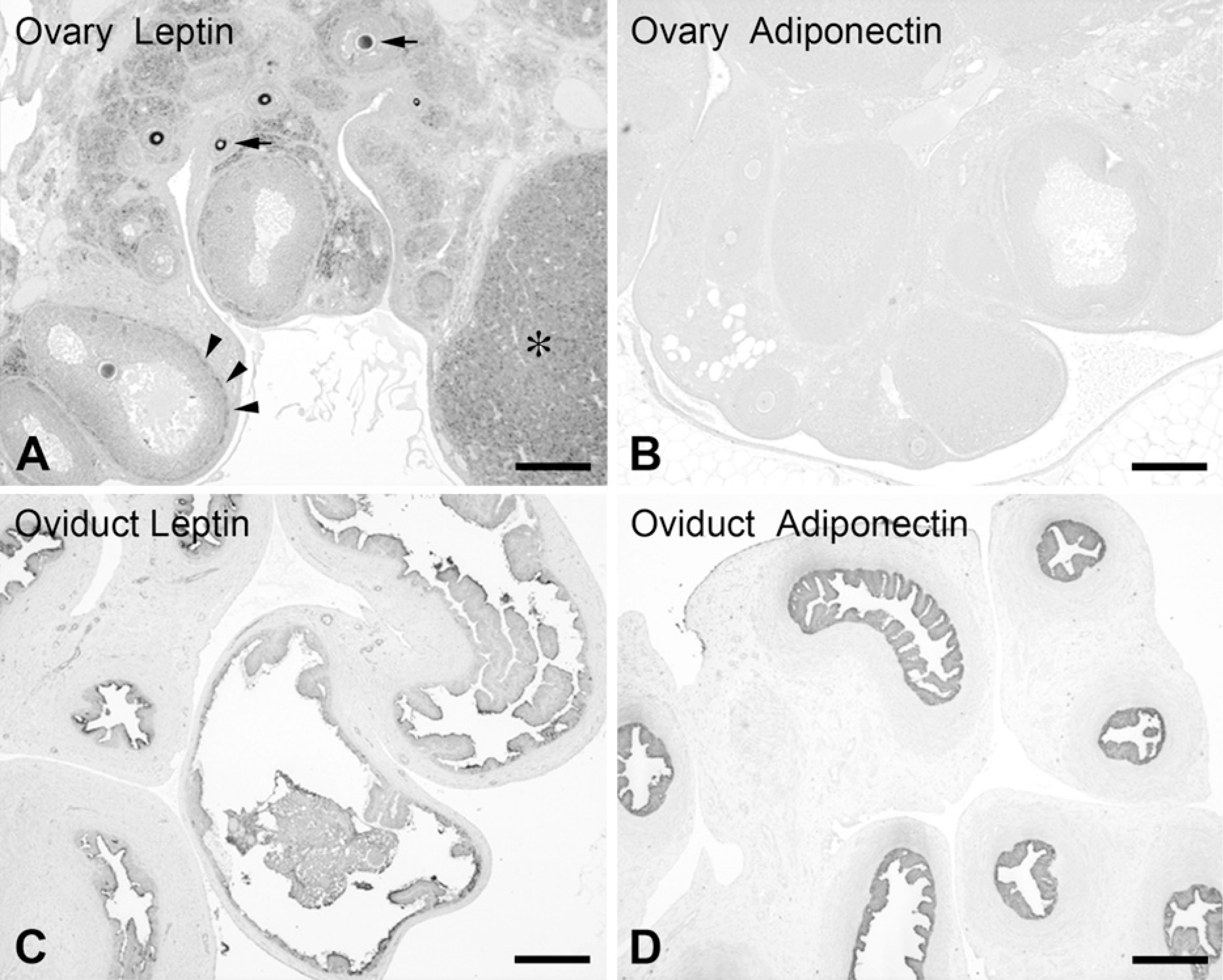
Detection of leptin and adiponectin in the rat ovary (
Animals were sacrificed by decapitation in either proestrous (∼5:00 PM) or estrous (∼11:00 AM) phase (
Part of the material was processed for ultrastructural study. Small pieces were fixed overnight at 4C in 4% glutaraldehyde in 0.1 M sodium cacodylate buffer, pH 7.2, and postfixed for 2.5 hr at 4C in 1% phosphate-buffered osmium tetroxide. For immunocytochemical studies, Zamboni's formaldehyde–picric acid mixture (Stefanini et al. 1967) was used for 12 hr at room temperature. After fixation, all samples were dehydrated with ethanol, washed with propylene oxide, and embedded in Epon (Luft 1961).
RIA of LH and FSH
Serum concentrations were measured in duplicate by RIA using a double-antibody method with kits supplied by the National Institutes of Health (Bethesda, MD) and a previously described microassay method (Sánchez-Criado et al. 1990). Rat LH-I-10 and FSH-I-9 were labeled with 125I by the chloramine T method (Greenwood et al. 1963). All serum samples were assayed in the same assay. The intra-assay coefficients of variation were <8% (6% and 5% for LH and FSH, respectively). Assay sensitivities were 3.75 pg/tube for LH and 20 pg/tube for FSH. Hormone concentrations were expressed as ng/ml. All samples were measured in the same assay.
Results were expressed as mean ± SEM. Because data were not normally distributed, the non-parametric Mann–Whitney U test was performed. Level of significance was set at
Immunohistochemistry
Immunohistochemical controls included negative controls incubated with normal goat serum and diluted 1:20 in place of the primary antiserum and positive controls, using rat adipose and ovarian samples.
RT-PCR
Total RNA was extracted from oviducts by homogenization with an Ultra-Turrax R25 basic (IKA Werke GmbH; Staufen, Germany) using TRIzol reagent (Invitrogen; Carlsbad, CA), and RNA concentrations were determined from absorbance at 260 nm. After initial treatment with DNase I (Roche; Basel, Switzerland), RNA (1 μg) was reverse-transcribed with a random hexamer and M-MLV reverse transcriptase (Gibco BRC; Paisley, UK). Primers used to amplify leptin and adiponectin cDNA (GenBank reference included) are detailed in Table 2. The pair of primers was designed using the Oligo 4.05 Primer Analysis Software (National Biosciences; Plymouth, MN). cDNA was amplified in the appropriate number and range of cycles and temperatures (Table 2). Each cycle included the following parameters: 94C for 30 sec, specific annealing temperature (Table 2) for 30 sec, and 72C for 30 sec, with a final extension step at 72C for 7 min. Amplification was linear under these conditions and was carried out in a GeneAmp PCR System 2400 (Perkin-Elmer; Norwalk, CT). All PCRs for each gene were performed at the same time and with the same batch of Taq polymerase to reduce variations in PCR efficiency. Amplification of rat β-actin (Table 2) was performed simultaneously on the same samples to assess RNA integrity. Negative controls included amplifications: in the presence of the RNA template and Taq polymerase but in the absence of reverse transcriptase and in the presence of distilled water in the place of RNA template. Amplified products were resolved in a 1% agarose gel with ethidium bromide and visualized in a UV Transiluminator (Gel Printer Plus System; TDI Tecnología para Diagnostico e Investigación, Madrid, Spain). RNA obtained from adipose samples was used as positive control for the PCR.
Results
Serum LH and FSH Concentrations
In the present experiment we measured serum LH and FSH levels in rats in the proestrous and estrous phases (
IHC distribution of leptin (
Immunohistochemical Localization of Leptin and Adiponectin
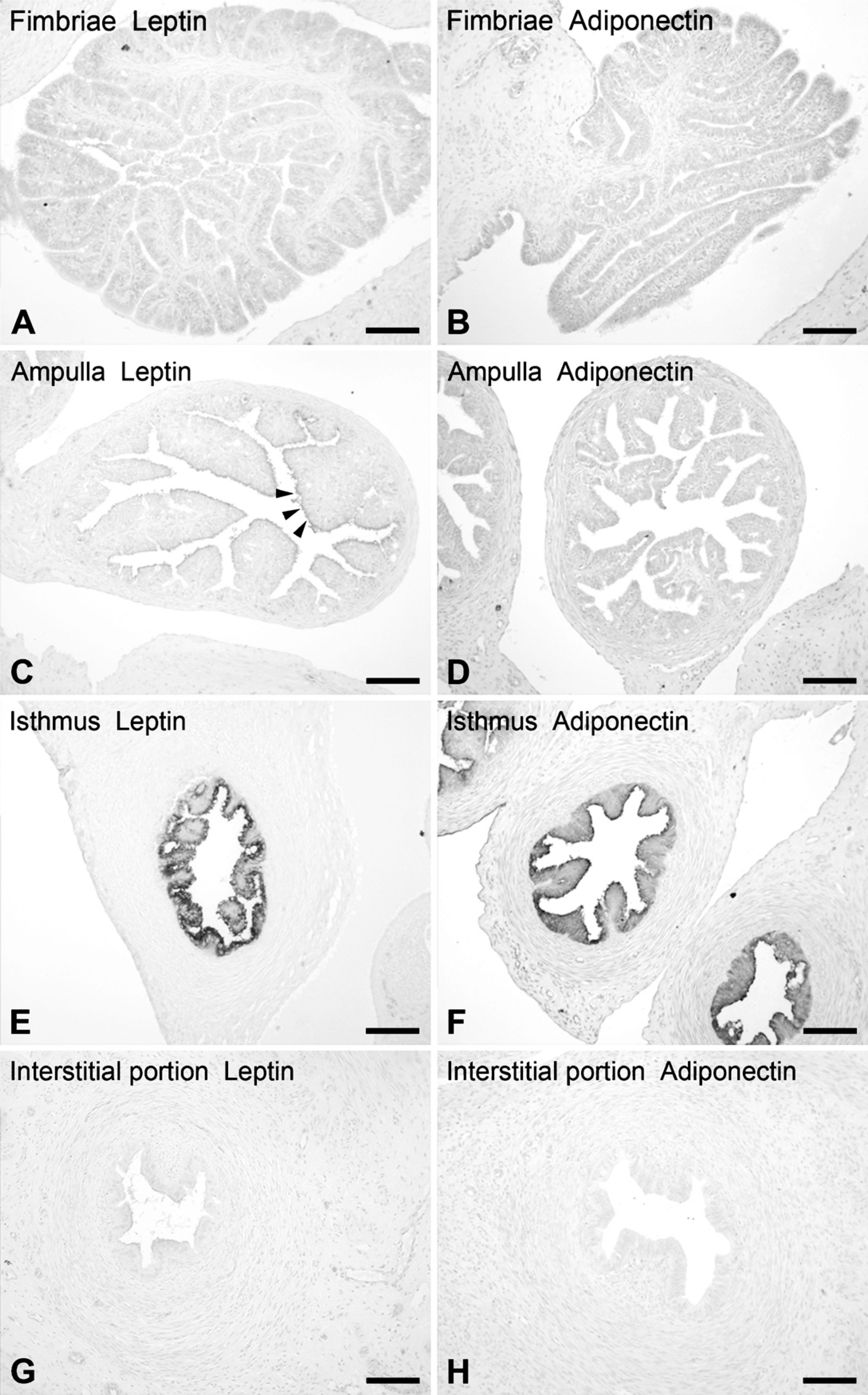
The presence of leptin and adiponectin in the rat oviduct was demonstrated by IHC with light (Figure 2–Figure 5) and electron microscopy (Figure 6). Both adipokines displayed a similar pattern of expression, their immunoreactivities appearing in secretory cells of the oviduct epithelium. Staining intensity was stronger for adiponectin. The other layers of the wall (the thin subepithelial connective tissue, muscularis, and serosa) were negative. Samples of ovary and adipose tissue that appeared near the oviduct in the same sections were used as positive controls (Figure 2). In the ovary, leptin was detected in oocytes, theca cells, corpora lutea, and interstitial glands (Figure 2A). In contrast, IHC for adiponectin was negative in the ovary (Figure 2B). Adipose tissue was always positive for both antibodies. No immunostaining was observed in the negative controls.
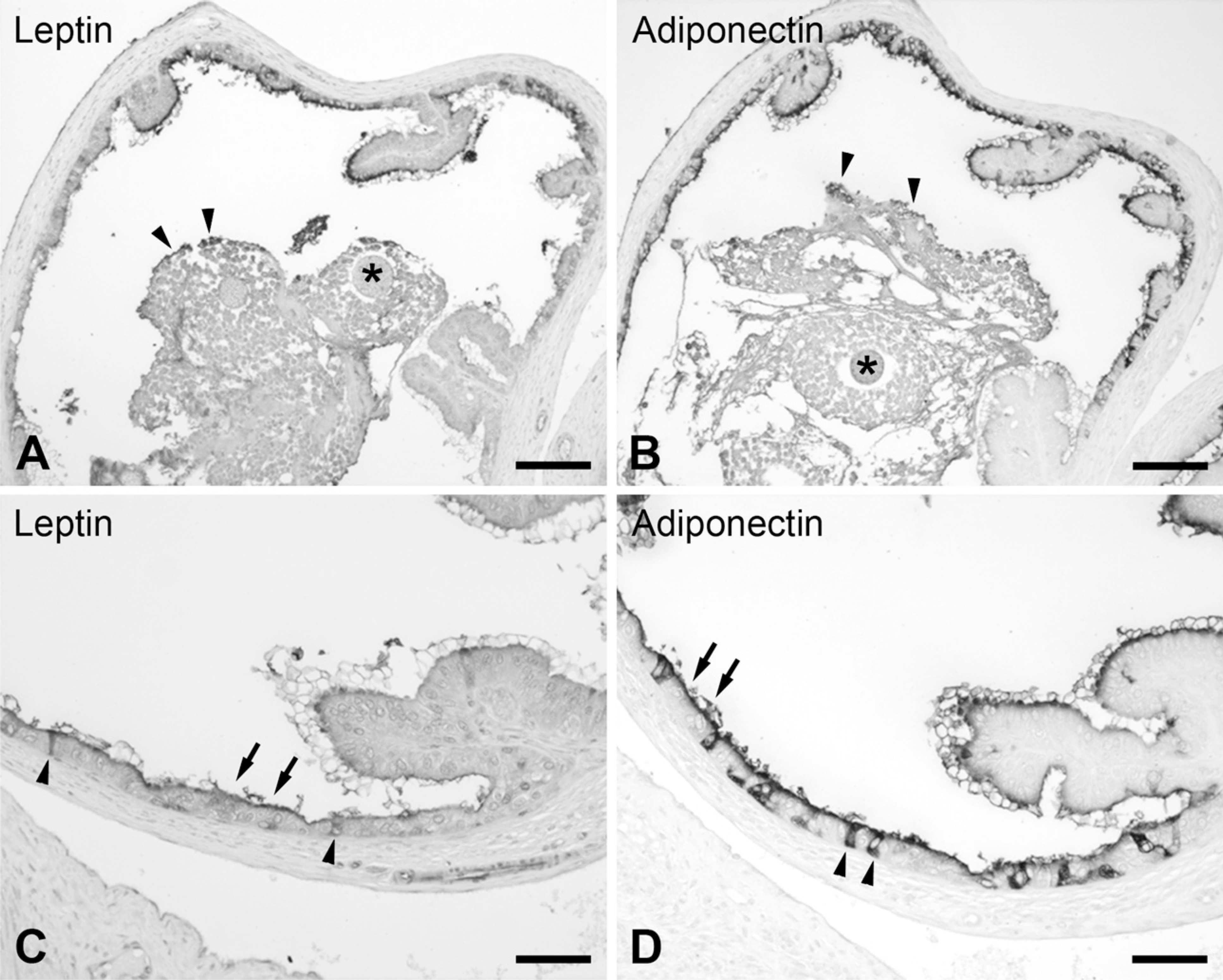
Consecutive sections of the ampulla in estrous, with some oocytes present in its lumen. (
The rat oviduct is ∼1 cm long, extremely convoluted, and is comprised of four morphologically distinct segments: fimbriated infundibulum, ampulla, isthmus, and interstitial portion. The distribution of leptin and adiponectin showed regional differences all through the oviduct (Figure 3). Noteworthy is that the strongest signal both for leptin and adiponectin was observed in the epithelium of the isthmus (Figures 3E and 3F). Immunoreactivity also appeared in the ampulla, with variations in the intensity between leptin and adiponectin (Figures 3C and 3D); adiponectin stains stronger than leptin. Moreover, we observed variations in staining among the different reproductive states studied (Figures 3C and 3D and Figure 4). In the other regions, fimbriated infundibulum (Figures 3A and 3B) and the interstitial portion (Figures 3G and 3H), leptin and adiponectin were absent. Likewise, uterine tissue observed in some slides was always negative for both antisera.
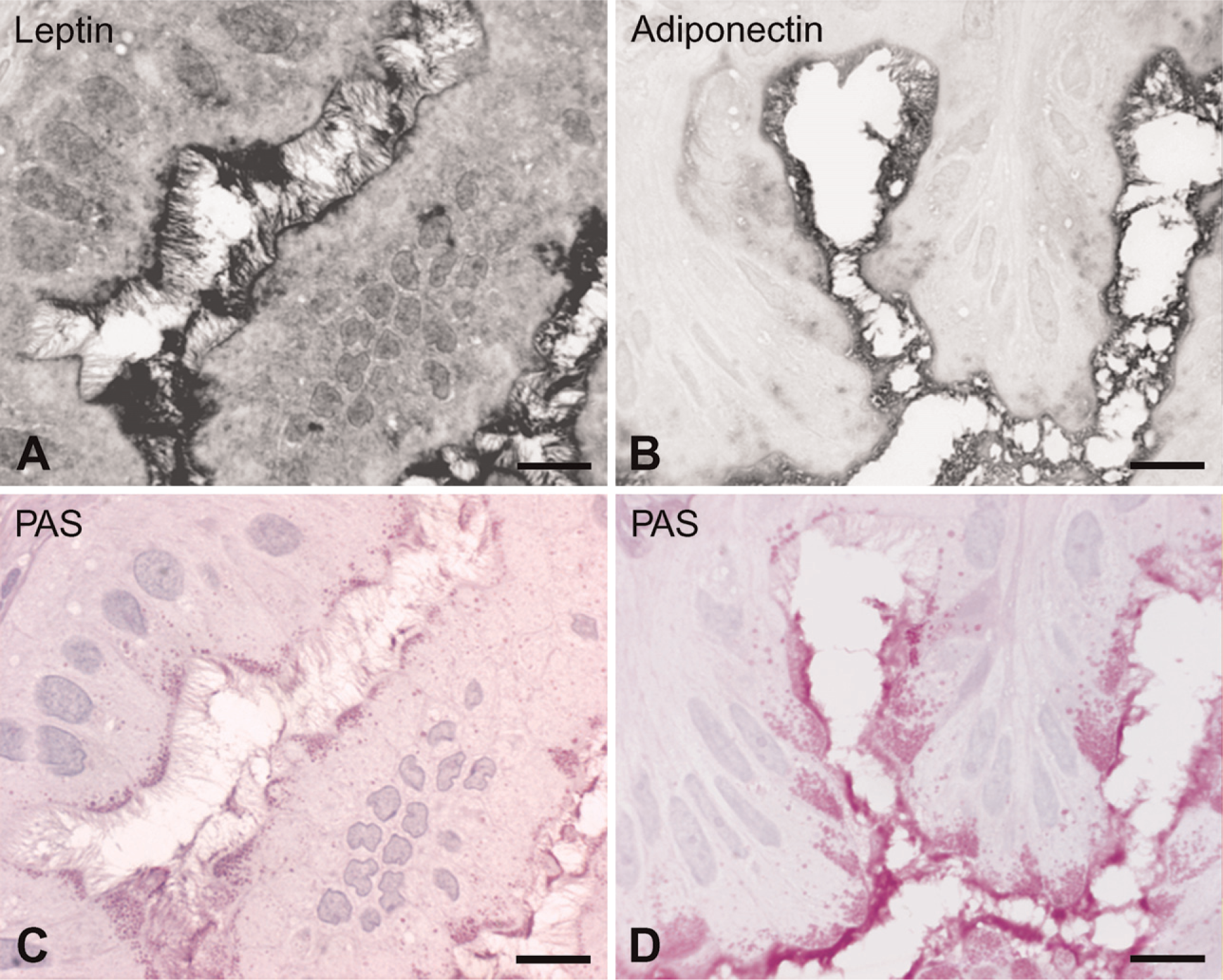
Semithin sections of Epon-embedded isthmus show secretory cells immunostained for leptin (
With the exception of the ampulla, the two phases of the estrous cycle studied in this work, proestrous and estrous, did not show differences in the morphology of the distinct segments of the oviduct or in the localization of leptin and adiponectin. In the ampullary region, significant variations correlated with the stage of the reproductive cycle were observed. In proestrous samples, when the mucosa was highly folded and ciliated epithelial cells predominated, a faint reaction restricted to the apical membrane of some cells in contact with the central lumen of the duct was usually observed for both antisera (immunostaining for leptin shown in Figure 3C). Immunoreaction was absent for both leptin and adiponectin in a number of samples (Figure 3D). In the estrous phase, when postovulatory oocytes were present in the lumen and the mucosa presented less branched folds, the ampulla was strongly labeled (Figure 4). It is worth mentioning that there is a dramatic increase in the number of secretory cells of the epithelium, which is responsible for the labeling (Figures 4C and 4D). In this case, the eggs at the lumen were negative for both antisera and were surrounded by several layers of follicular cells (corona radiata), which interacted directly with the oviduct and showed a few positive cells.
In the isthmus at estrous and proestrous, most of the epithelial cells displayed strong leptin and adiponectin immunoreactivity (Figure 5 and Figure 6). In semithin sections, staining appeared in the apical cytoplasm of the secretory cells and had a granular aspect (Figures 5A and 5B). This IHC pattern of immunoreactivity for both hormones was very similar to the staining obtained with the periodic acid Schiff technique (Figures 5C and 5D). Under the electron microscope, immunoreactivity for both proteins was consistent with the light microscopy results. The signal for leptin and adiponectin appeared in the apical secretory granules and in the long stereocilia characteristic of the secretory cells of the rat oviduct (Figure 6). In both cases, ciliated cells were not labeled.
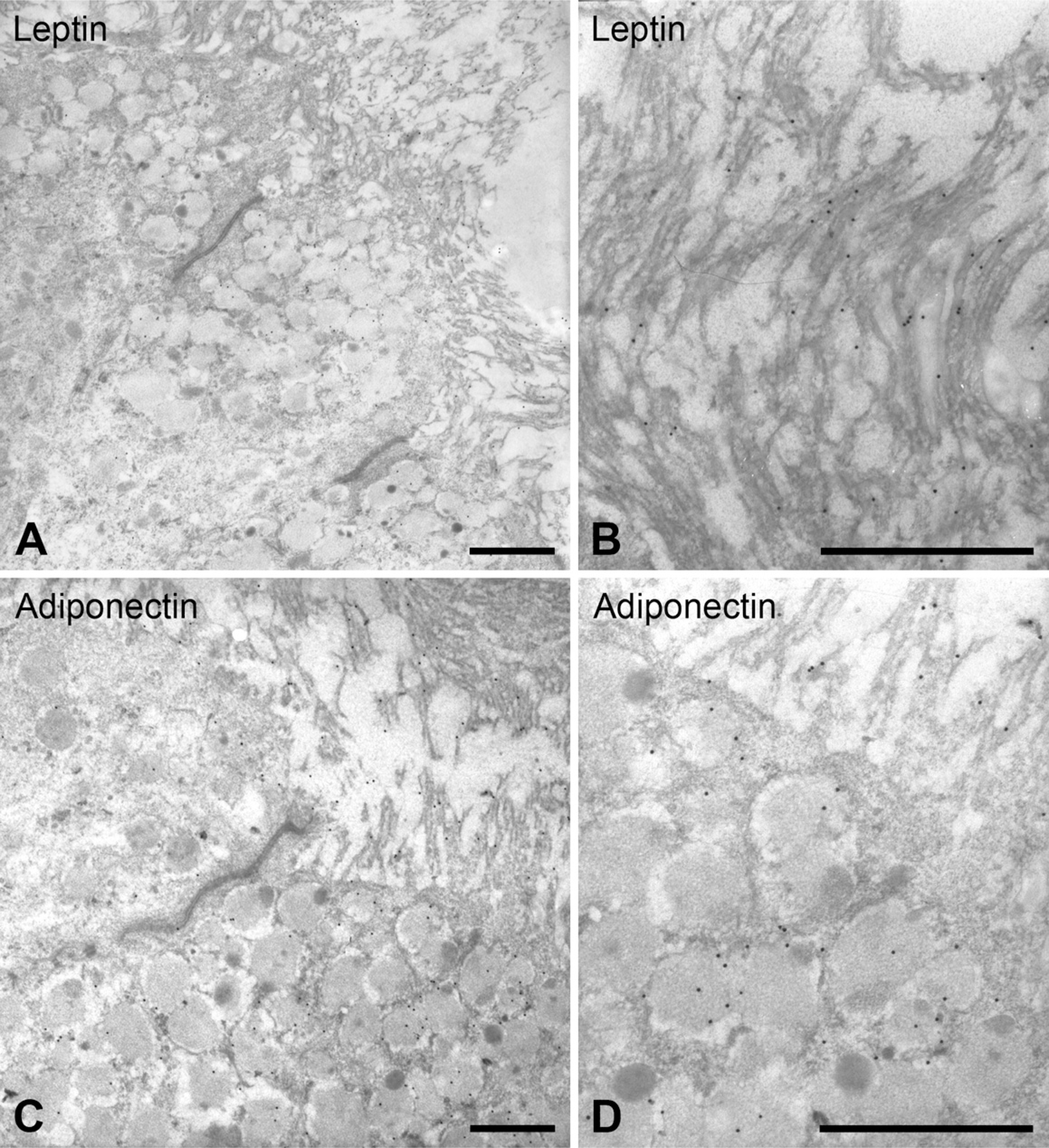
Isthmus secretory cells observed in thin sections immunostained with leptin (
Expression of Leptin and Adiponectin mRNA
PCR products amplified using the specific primers for leptin showed a clear band at the predicted size of 405 base pairs in the oviduct and the ovary extracts, as in the adipose samples used as control tissue (Figure 7).
However, adiponectin expression was found in the oviduct and at very low levels in the rat ovary (Figure 7). These PCR results agreed with the IHC results, where no presence of adiponectin was detected (Figure 2B). As expected, adiponectin mRNA expression was detectable in adipose tissue.
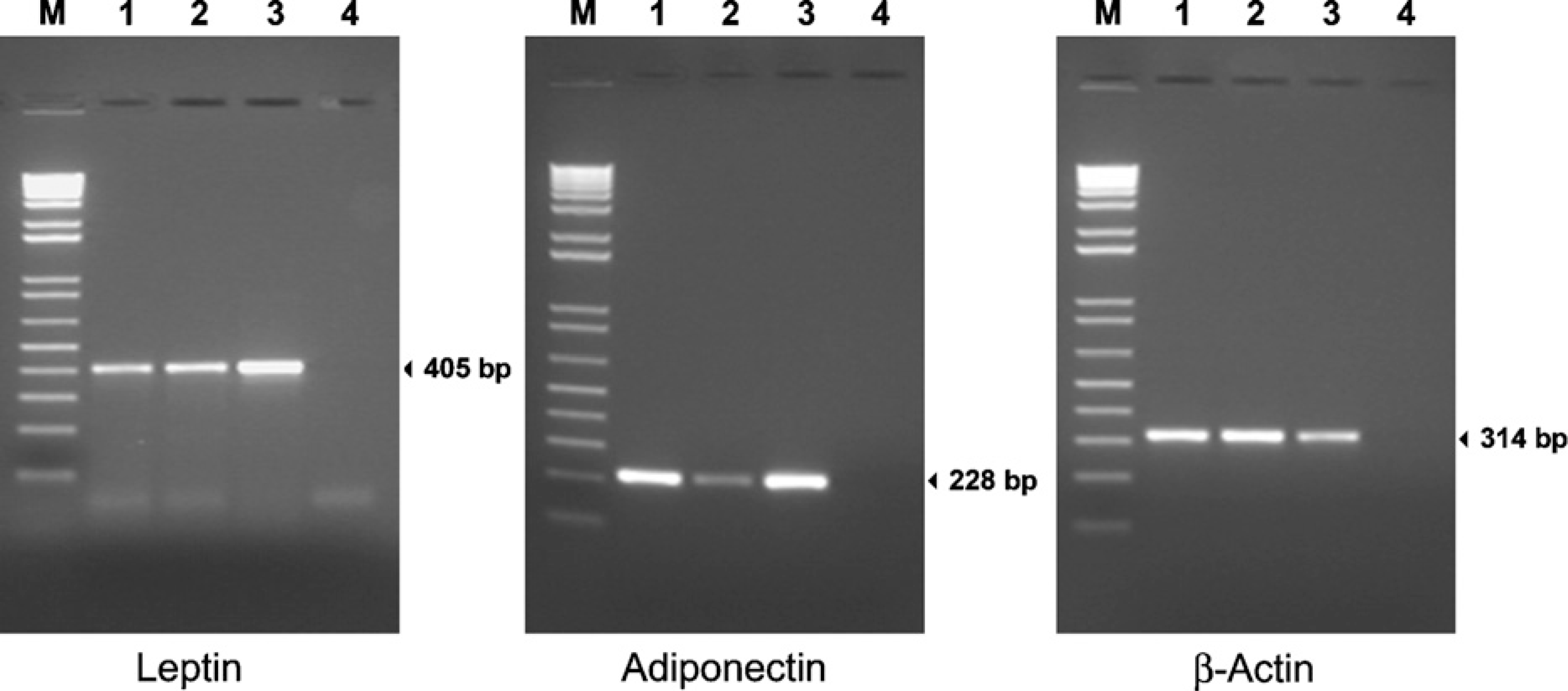
Representative analysis of leptin (left panel) and adiponectin (middle panel) mRNA expression in rat oviduct (Lane 1) and ovary (Lane 2) samples by RT-PCR. The size of the predicted amplified product leptin (405 bp), adiponectin (214 bp), and β-actin (314 bp) are indicated at right. M is the molecular weight marker (Ladder 100). RNA from adipose tissue was used as positive control (Lane 3) and distilled water as negative control (Lane 4).
As a positive control, rat β-actin was assayed in all specimens to verify the efficiency of cDNA synthesis from extracted RNA (Figure 7).
Discussion
This study examined the expression of two adipocyte hormones (adipokines), leptin and adiponectin, throughout the different regions of the rat oviduct. The identification and potential role of these two hormones in the oviduct will be discussed.
The mammalian oviduct is a dynamic organ in which important reproductive functions take place, including gamete transport and maturation, fertilization, and early cleavage-stage embryonic development (Hunter 1998). During late follicular development and estrous, this organ undergoes the most significant physiological and biochemical modifications that contribute to the establishment of an optimal microenvironment for both gametes and the early embryo. Analyses of oviduct fluid have demonstrated temporal and spatial distribution of macromolecules (Buhi 2002). Because the time an embryo spends at the oviduct is essential for its proper development, a more extensive and refined knowledge and understanding of the first environment to which embryos are exposed is necessary.
Different facts indicate that there is a link between adipokines and female reproduction. The two hormones studied in this work, leptin and adiponectin, are two of the best-characterized adipokines and have been related to reproductive performance. Different central and peripheral effects of leptin and adiponectin have been described in relation to reproduction (Caprio et al. 2001; Mitchell et al. 2005). The wide pattern of expression described for leptin suggests that this hormone has a direct role in many different functions such as ovulation, steroidogenesis, and embryo development (Duggal et al. 2000; Caprio et al. 2001; Mitchell et al. 2005). However, the possible implications of adiponectin in ovarian functions have received almost no attention (Mitchell et al. 2005).
In the oviduct, studies of the expression and activity of adipokines are also limited to leptin, suggesting its possible involvement in the early-developing embryo (Kawamura et al. 2002; Craig et al. 2005; Mitchell et al. 2005). Until now, adiponectin expression in the oviduct has not been documented, and only a few studies have addressed the expression of leptin in the human, porcine, and mouse oviduct (Kawamura et al. 2002, 2003; Liu et al. 2003; Craig et al. 2005). First, leptin and its receptor (mRNA and protein) were identified in the oviduct epithelium of the pregnant mouse, with weaker expression in virgin and non-pregnant mice (Kawamura et al. 2002, 2003). However, leptin was not detected in the oviduct fluid because of its very small quantities. In the human oviduct, expression of leptin and its receptor has also been detected by in situ hybridization and IHC (Liu et al. 2003). Moreover, it has been demonstrated that leptin, but not its receptor, is produced in the porcine oviduct (Lin et al. 2000; Craig et al. 2005).
Data obtained in this study show that leptin and adiponectin are expressed in the rat oviduct and follow a superimposable pattern of distribution. Specifically, both the protein and mRNA for the two adipokines have been localized. This is the first report of adiponectin expression in the oviduct. The strongest signal for both leptin and adiponectin appeared in the secretory cells of the isthmus. Moreover, electron microscopy revealed, for the first time, that these hormones are localized in the secretory granules and in the long stereocilia, suggesting that they are secreted to the oviduct fluid. Such an exocrine mechanism of secretion has also been reported for gastric leptin present in the exocrine granules of the chief cells of the stomach (Bado et al. 1998). Furthermore, immunostaining also appears in the epithelium of the ampulla, showing differences associated with the estrous cycle. Staining intensity in the ampulla increased from proestrous to estrous, suggesting that ovarian steroids regulate leptin and adiponectin expression. These results are consistent with the rapid cyclic changes easily recognized in the gross morphology of the rat oviduct and that are most evident in the ampulla (Shirley and Reeder 1996). In this region, ciliated epithelial cells prevail before ovulation, whereas secretory cells are more numerous at the postovulatory stages. In addition, activity of the secretory cells is increased at estrous and metestrous, which may explain our observation that leptin and adiponectin expression are higher in the estrous oviduct.
Our results highlight that the maximum expression for the two adipokines was found in the isthmus (where spermatozoa and early embryos are located) and in the ampulla at estrous (where oocytes are present), which adds further weight to the observation that leptin may have direct effects on oocyte quality and embryo development (Antczak and Van Blerkom 1997; Matsuoka et al. 1999; Kawamura et al. 2002, 2003). Adiponectin also seems to participate in these functions although observations are still preliminary (Downs et al. 2002; Lord et al. 2005). It has been previously shown that both leptin protein and mRNA are present in the preovulatory oocyte in the rat ovary (Archanco et al. 2003). However, no expression of either leptin or adiponectin has been detected in the postovulatory oocyte. In the cumulus cells surrounding the postovulatory oocyte, both hormones stained slightly, supporting the notion that these cells remain active and enhance the processes in the lumen of the oviduct (Hunter 1998). In the literature, the presence of leptin in female gametes or embryos is not always consistent and depends on the different species studied and/or methods used (Antczak and Van Blerkom 1997; Cioffi et al. 1997; Kawamura et al. 2003). Taking all the data into consideration, it is likely that leptin may act in a paracrine and/or autocrine manner, produced by maternal organs and/or by the embryos themselves (Kawamura et al. 2003).
In conclusion, we have described the presence of leptin and adiponectin in the rat oviduct. Although the physiological significance of these two hormones remains to be clarified, it seems plausible that they may have important roles in all local functions from transport and maturation of the oocytes, spermatozoa, and embryos to fertilization.
Footnotes
Acknowledgements
This research project was supported by PIUNA (The Research Plan at the University of Navarra) and the Spanish Ministry of Science and Technology (Project no. BCM2000-1137).
The authors are grateful to Paz Zamora, Silvia González, and Ainhoa Urbiola for their technical work, and especially to Blanca Irigoyen for excellent assistance in electron microscopy techniques. We also thank Cóilín O hAodha for the English revision.