
Abstract
Leptin, a potent anorexigenic hormone, is found in the anterior pituitary (AP). The aim of this study was to determine whether and how pituitary leptin–bearing cells are regulated by nutritional status. Male rats showed 64% reductions in pituitary leptin mRNA 24 hr after fasting, accompanied by significant (30–50%) reductions in growth hormone (GH), prolactin, and luteinizing hormone (LH), and 70–80% reductions in target cells for gonadotropin-releasing hormone or growth hormone-releasing hormone. There was a 2-fold increase in corticotropes. Subsets (22%) of pituitary cells coexpressed leptin and GH, and <5% coexpressed leptin and LH, prolactin, thyroid-stimulating hormone, or adrenocortico-tropic hormone. Fasting resulted in significant (55–75%) losses in cells with leptin proteins or mRNA, and GH or LH. To determine whether restoration of serum glucose could rescue leptin, LH, and GH, additional fasted rats were given 10% glucose water for 24 hr. Restoring serum glucose in fasted rats resulted in pituitary cell populations with normal levels of leptin and GH and LH cells. Similarly, LH and GH cells were restored in vitro after populations from fasted rats were treated for as little as 1 hr in 10–100 pg/ml leptin. These correlative changes in pituitary leptin, LH, and GH, coupled with leptin's rapid restoration of GH and LH in vitro, suggest that pituitary leptin may signal nutritional changes. Collectively, the findings suggest that pituitary leptin expression could be coupled to glucose sensors like glucokinase to facilitate rapid responses by the neuroendocrine system to nutritional cues.
Keywords
L
Theubiquitous distribution of leptin receptors has suggested that this cytokine performs additional functions throughout the body (Fruhbeck 2006). For example, leptin may regulate neuroendocrine systems, particularly in the reproductive system (Ahima et al. 1996; Casanueva and Dieguez 1999; Moschos et al. 2002). Rats without functional leptin receptors (Zucker fa/fa) and mice without functional leptin proteins (ob/ob) both exhibit characteristics of morbid obesity, insulin resistance, delayed or impaired pubertal development, and pituitaries with low numbers of somatotropes or gonadotropes (Isozaki et al. 1999; Popovic et al. 2001). Leptin levels are permissive for puberty, because levels of fat stores must reach a threshold for the animal to reproduce (Ahima et al. 1996; Nagatani et al. 1998, 2000; Casanueva and Dieguez 1999; Gonzalez et al. 1999; Urbanski 2001; Mann and Plant 2002). Without leptin proteins or receptors, rats will not reach full reproductive maturity, but exogenous leptin injections given to leptin-deficient animals will restore fertility (Barash et al. 1996; Cheung et al. 1997).
Leptin regulates reproduction by direct effects on the hypothalamic–pituitary axis. It stimulates the secretion of both luteinizing hormone (LH) and gonadotropin-releasing hormone (GnRH) (Finn et al. 1998; Nagatani et al. 1998, 2000; Gonzalez et al. 1999; Tezuka et al. 2002), and restores LH secretion in fasted mice, rats, hamsters, and monkeys (Ahima et al. 1996; Finn et al. 1998; Nagatani et al. 1998, 2000; Schneider et al. 1998, 2000, 2002; Gonzalez et al. 1999; Schneider and Zhou 1999). Leptin also directly stimulates LH secretion from pituitary cells in vitro (Yu et al. 1997a, b; Walczewska et al. 1999; De Biasi et al. 2001; Ogura et al. 2001). In addition, leptin also regulates growth hormone (GH) (Casanueva and Dieguez 1999), although there is lack of agreement about the exact direction of its regulation (Aubert et al. 1998; Vuagnat et al. 1998; Casanueva and Dieguez 1999; Pombo et al. 1999; Roh et al. 2001). Leptin stimulates GH secretion mediated by GH-releasing peptide (Chen et al. 2001; Roh et al. 2001) and restores GH pulses in fasted rats (Vuagnat et al. 1998; Pombo et al. 1999).
A growing body of evidence suggests, however, that the adipocyte source of leptin may be too sluggish to regulate rapid neuroendocrine responses. For example, the rise in serum leptin is too slow for it to be a regulator of the LH surge (Akhter et al. 2007). Also, Schneider et al. report that LH pulses lost by fasting can be restored within a few hours by nutrition alone (Schneider et al. 2000, 2002). The timing of this response was too rapid for it to have been due to the restoration of adipocyte leptin (Bronson 1986; Bronson and Heideman 1990; Cameron 1996).
This evidence led to the hypothesis that local sources of leptin, such as those in the pituitary, might be part of a neuroendocrine regulatory circuit. Supportive evidence from our laboratory demonstrated a sharp rise in pituitary leptin just before the LH surge, and stimulation of pituitary leptin secretion by GnRH (Akhter et al. 2007). To further investigate its potential role in the pituitary, we tested additional in vivo and in vitro conditions to determine states that might regulate leptin.
We hypothesized that pituitary leptin might be regulated by nutrient status. It is already known that significant decreases in adipocyte leptin mRNA expression can be seen after 8 hr of fasting (Zhang et al. 1994; Kowalska et al. 1999; Gui et al. 2003), with losses in leptin mRNA of 50% (Igel et al. 1996) or 85–90% (MacDougald et al. 1995; Mizuno et al. 1996) after 24 hr of fasting. Therefore, we first developed an in vivo model of food deprivation in which the expression of pituitary leptin was selectively reduced, without changes in serum leptin. After initial studies showed parallel reductions in pituitary leptin, LH, and GH, the experiments were expanded to determine whether changing nutrients, such as serum glucose in vivo or exogenous leptin in vitro could restore the losses in gonadotropes and somatotropes. This report presents the findings of these experiments, which support the hypothesis that pituitary leptin is regulated by nutrient status and that as little as 10 pg/ml exogenous leptin for 1 hr in vitro can restore expression of GH and LH in cells from fasted animals.
Materials and Methods
Collection of Pituitaries
Male and female Sprague-Dawley rats obtained from Harlan Sprague Dawley (Indianapolis, IN) were used throughout this study. Animals were housed three per cage, with a 12-hr light-dark cycle (lights on at 6:00 AM), and maintained at a constant room temperature of 68C. A standard pellet chow diet (rodent diet #8640; Harlan Teklad, Madison, WI) and water were available ad libitum (in all animals, in all experiments, except for the fasting experiments). Animals were allowed to become acclimated for ∼2 weeks before any experiments commenced. The rats were anesthetized with IP injections of sodium pentobarbital (24 mg/kg or 0.06 ml/250-g rat) and then sacrificed via guillotine. The animal care protocol was approved annually by the Animal Use and Care Protocol Committee, University of Arkansas for Medical Sciences.
Fasting Experiments
At least 12 major groups of weight- and age-matched male rats were used for these studies. Some of the groups of rats provided whole anterior pituitaries for either protein or mRNA extracts; others provided pituitary cells for the analysis of hormone stores by cytochemistry. The experimental design for the fasting was similar, irrespective of the end point assays.
At 9:00 AM, male rats were divided into two or three separate groups, with two to three rats in each group. The animals were weighed and given a known amount of water (both controls and fasted). In the last four groups, the third group of animals was given glucose water (glucose fasted). All rats were kept in separate cages for a period of 24 hr. For the controls, a standard pellet chow diet (Harlan Teklad rodent diet) and water were available ad libitum. The animals that were strictly fasted were allowed only water ad libitum, and the fasted rats given glucose were only allowed a 5% or 10% glucose solution (G7021-1KG; Sigma, St. Louis, MO) ad libitum. After a glucometer detected changes in serum glucose in these rats, only those treated with 10% glucose were used, because rats given 5% glucose did not have normal serum glucose. These data are shown in Table 1.
The fasted and fasted glucose conditions were held for a period of 24 hr. Pituitaries were taken during the morning 24 hr after the start of the fast. The rats were anesthetized, reweighed, and sacrificed. Trunk blood was taken, and the amount of water consumed was recorded. Serum was collected for leptin enzyme immunoassay (EIA) from American Laboratory Products Company (ALPCO Diagnostics; Salem, NH) and R&D Systems, Inc. (Minneapolis, MN). Serum glucose was also measured by a glucometer, as stated above.
Dispersion of Pituitary Cells
Pituitaries were removed and dispersed into single suspensions according to the following protocol. After gently teasing away and discarding the neurointermediate lobe, the remaining APs were chopped into small pieces and transferred to a 15-ml tube that contained Dulbecco's modified Eagle's medium (DMEM) and then centrifuged for 1 min at 400 rpm. The anterior pituitaries were then dispersed, as previously described (McDuffie et al. 2004; Akhter et al. 2007), and plated for 1 hr on glass coverslips coated with poly-D-lysine to enhance cell adhesion in DMEM, which is serum free. Viability tests showed 99–100% viability during this brief culture period, and the cells remained rounded after settling on the coverslips. These very short-term (1-hr) cultures are used for reasons outlined in our recent report (Akhter et al. 2007). To avoid stimulation of leptin by additives in the media, the cultures were allowed to settle in as little time as possible in basic DMEM. The cells were fixed with 2.5% glutaraldehyde diluted in 0.1 M of phosphate buffer for 30 min at room temperature. Fixation was followed by four washes of 15 min each with 0.1 M phosphate buffer containing 4.5% sucrose and glycine.
Cytochemical Labeling Protocols
In situ hybridization involved the application of prehybridization reagents, followed by 10–100 ng/ml of a biotinylated probe complementary to leptin mRNA, as described recently (McDuffie et al. 2004; Akhter et al. 2007). GH mRNA was detected with 5–10 ng/ml of biotinylated probe, following the protocol outlined in our recent studies (Childs et al. 2000, 2005; Iruthayanathan et al. 2005), and the LHβ mRNA was detected with 20–50 ng/ml biotinylated probe, as previously described (Childs et al. 1992, 1994b). The probes were produced by GeneDetect.com (www.GeneDetect.com), by Greenstar protocols, which provided high levels of biotin per probe. After 12–15 hr hybridization at 39.6C (leptin probe) or 35.6C (GH or LHβ probes), the probes were detected by monoclonal anti-biotin and streptavidin peroxidase in a sandwich technique, as described previously (Childs et al. 1992, 1994b, 2005; McDuffie et al. 2004; Iruthayanathan et al. 2005; Akhter et al. 2007). Controls included the substitution of sense probes for the antisense probes, or the omission of the antisense probe in the sequence.
Fasting and glucose restoration

Average of three experiments ± SEM; nine rats/group (27 total).
Fasted values<fed or fasted+glucose values; p<0.007.
Fasted or fasted+glucose values<fed values; p<0.001.
Immunocytochemical labeling for leptin proteins involved the protocol described in our previous publications (McDuffie et al. 2004; Akhter et al. 2007) with 1:37,000 anti-rat leptin (Sigma-Aldrich; St. Louis, MO). The controls for leptin immunolabeling are reported and illustrated in our recent publications (McDuffie et al. 2004; Akhter et al. 2007). Dual labeling detected leptin with adrenocorticotropic hormone (ACTH), thyroid-stimulating hormone (TSH), LH, GH, or prolactin, according to recent studies (Childs et al. 2005). The antisera dilutions used were 1:7000 anti-rat prolactin (Childs et al. 1999); 1:70,000 anti-rat TSH-β (Childs et al. 1989); 1:200,000 anti-rat GH (National Institutes of Health, Hormone Distribution Program)(Childs et al. 1994b, 1999, 2005; McDuffie et al. 2004; Iruthayanathan et al. 2005); 1:80,000-100,000 anti-17-39 ACTH (Childs et al. 1989, 1992, 1994b, 1999); and 1:100,000 anti-bLHβ (Akhter et al. 2007) (a gift from Dr. J. G. Pierce). The controls for each of these anti-sera are described in each of these previous studies (Childs et al. 1989, 1992, 1994b, 1999, 2005; McDuffie et al. 2004; Iruthayanathan et al. 2005; Akhter et al. 2007).
Affinity cytochemical labeling for biotinylated analogs of GnRH or growth hormone–releasing hormone (GHRH) was done as described in recent studies (Childs et al. 1994a, 1999). Freshly dispersed, living cultures were incubated in biotinylated GnRH or biotinylated GHRH for 10 min and then fixed in 2% glutaraldehyde for 30 min. These analogs have a potency equal to that of native GnRH or GHRH and only bind receptors in healthy, living pituitary cells (Childs et al. 1994a, 1999) The target cells for each of the analogs were detected by avidin-biotin-peroxidase complex (Vector Laboratories; Burlingame, CA) and nickel-intensified diaminobenzidine, as described previously (Childs et al. 1994a, 1999). Over 90% of target cells for biotinylated GnRH store gonadotropins (Childs et al. 1994a), although a subset of these are somatogonadotropes and express GH. Similarly, >90% of target cells for GHRH store GH (Childs et al. 1999, 2005), although a subset of GHRH target cells express gonadotropins and prolactin (Childs et al. 1999). Controls for these protocols have been described in previous studies (Childs et al. 1994a, 1999, 2005). They demonstrate that 10–100-fold excess of non-biotinylated GnRH or GHRH compete successfully for the receptor site and neutralize labeling for their respective biotinylated neuropeptides. However, the same concentrations of other neuropeptides do not compete for the receptors, and labeling is unaffected.
Image Analysis
The Bioquant Nova image analysis system (Bioquant; Nashville, TN) was used to digitize the images (taken at ×40) and analyze the density of the label, as described previously (Iruthayanathan et al. 2005; Akhter et al. 2007). In addition, counts of labeled cells were done following single or dual labeling, as described previously (Childs et al. 1989, 1992, 1994a, b, 1999, 2005; McDuffie et al. 2004; Iruthayanathan et al. 2005; Akhter et al. 2007) to learn whether the treatment affected percentages of cells bearing leptin with and without pituitary hormones.
Immunoassays
A leptin mouse/rat EIA kit (ALPCO Diagnostics or R&D Systems, Inc.) was used to detect serum leptin proteins, which were frozen in small aliquots after they were collected. In addition, some serum samples were sent to Linco Diagnostics (Millipore; Billerica, MA) for radioimmunoassay. These assays have been described in previous studies (Akhter et al. 2007). Interassay and intra-assay variation coefficients were <4.7% and <4.4%, respectively.
Protein Extraction
Pituitary cells from three rats/group (described above) were dispersed and resuspended in 2000 μl of DMEM as previously described (McDuffie et al. 2004). The cells were spun at 4400 rpm for 10 min and then resuspended in 100 μl of NP40 lysis buffer, containing 1 μl each of the freshly added protease inhibitors aprotinin, leupeptin, and phenylmethyl-sulphonylfluoride. The cell lysate was mixed with a pipette and aliquoted in small tubes. It was then spun at 4C for 10 min. The supernatant was collected and frozen for storage at 280C.
RNA Extraction, cDNA Synthesis, and Quantitative RT-PCR
Whole anterior pituitaries from three rats/group of three groups were placed in RNeasy lysis tissue buffer (Qiagen; Valencia, CA) containing β-mercaptoethanol (as per the manufacturer's protocol) and homogenized. The extraction and cDNA synthesis were as described recently (Iruthayanathan et al. 2005; Akhter et al. 2007). Leptin mRNA was assayed by quantitative RT-PCR (QRT-PCR) as described in previous studies (Iruthayanathan et al. 2005; Akhter et al. 2007) in a Roche Light Cycler 1.5 (Roche; Indianapolis, IN) with the FAST-START DNA Master SYBR Green I enzyme mix (Roche). Housekeeping genes used to normalize the readings were rat ribosomal protein subunit 9 (Rps9) or hypoxanthine guanine phosphoribosyltransferase (Hprt). Tests of each housekeeping gene showed no changes in expression with fasting, and the primers for these genes were described in a previous publication (Iruthayanathan et al. 2005).
The forward primer for rat leptin was 5′-CCAGGATCAATGACATTTCACA-3′ (179 to 200), and the reverse primer was 5′-AATGAAGTCCAAACCGGTGA-3′ (230 to 249) (accession number NM_013076) (Akhter et al. 2007). The primers and conditions for GH were described previously (Iruthayanathan et al. 2005). Values were expressed as a ratio of the number of transcripts of the gene to that of one of the housekeeping genes.
Statistics
One-Way ANOVA was used to detect differences in a given set of experimental groups. Significant differences were then detected by Fisher's least significant differences post-hoc test, or Student's t-test; p<0.05 was considered significant. A posthoc power analysis was done to establish the number of replicates needed, as described previously (Akhter et al. 2007).
Results
Pituitary Cell Types Affected by Food Deprivation
The pilot studies done on the first three groups of rats (five rats/group) showed a weight loss in the fasted rats of 30 gm over the 24 hr period. Serum leptin in the fed group averaged 1.866 ± 0.467 ng/ml and that in the fasted group averaged 2.3 ± 0.7 ng/ml. There were no significant differences in serum leptin between the first two groups. However, there was a decrease in serum leptin in the latest groups of rats fasted with and without 10% glucose in their water (see later section).
The first objective of this study was to determine the effect of short-term fasting on the pituitary cells themselves. Figure 1 illustrates the cell counts, showing that fasting caused declines in percentages of pituitary cells with GH (by 31%; p<0.01), prolactin (by 53%; p<0.001), and LH (by 50%; p<0.001). In contrast however, fasting increased percentages of corticotropes more than 2-fold (p<0.001), from 10% to 21% of AP cells. The percentages of thyrotropes appeared unaffected by fasting. Figures 1B–1G illustrate the changes in immunolabeled corticotropes, somatotropes, and gonadotropes.
Focused Studies of Changes in Gonadotropes and Somatotropes
The studies then focused on other changes in gonadotropes and somatotropes, because they were affected most severely by the food deprivation. Detection of GH mRNA showed that somatotropes had not disappeared from the population. The counts showed a slight increase, from 33 ± 2% to 40 ± 2% (±SD; p<0.001) of AP cells. In contrast, cells with LH mRNA detected by in situ hybridization showed a 50% decline in expression, from 14 ± 2% to 7 ± 3% (±SD, p<0.001).
Tests of gonadotrope and somatotrope function also included the detection of changes in binding sites for GnRH or GHRH with protocols that detect biotinylated analogs of GnRH or GHRH (Childs et al. 1983a, b, 1994a, 1999). Figure 2 shows that fasting caused an 80% reduction in expression of GHRH-receptive cells (p<0.001) and a 73% reduction in GnRH-receptive cells (p<0.001). Figures 2B and 2C illustrate these cell populations in fed and fasted rat pituitary populations.
Fasting Effects on Leptin Proteins and mRNA
Food deprivation brought about a significant 64% reduction in the overall percentages of pituitary cells with leptin mRNA, from 33 ± 2% to 12 ± 1% of AP cells (p<0.001). Image analyses also showed a 40% reduction in the integrated optical density of label (p=0.03) (data not shown). In these first groups of fasted animals, there was also a 22% reduction in the percentages of cells with leptin proteins, from 36 ± 2% to 29 ± 2% of AP cells (±SE; p<0.029) (Figure 3A). Figures 3B–3G illustrate the in situ hybridization and the reduction in cells with leptin mRNA.
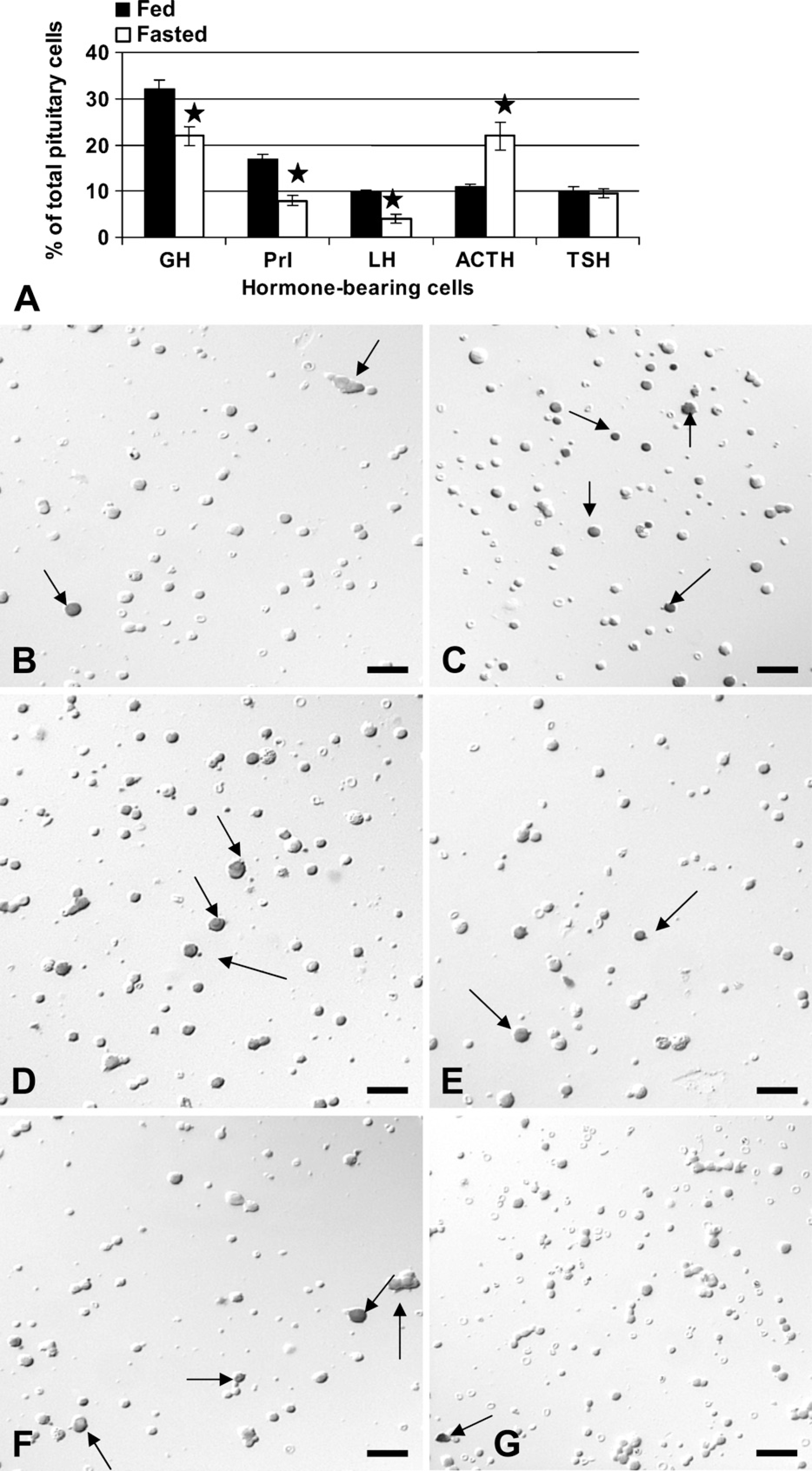
Counts and photographs show the effects of a 24-hr food deprivation on pituitary cells detected by immunolabeling. (
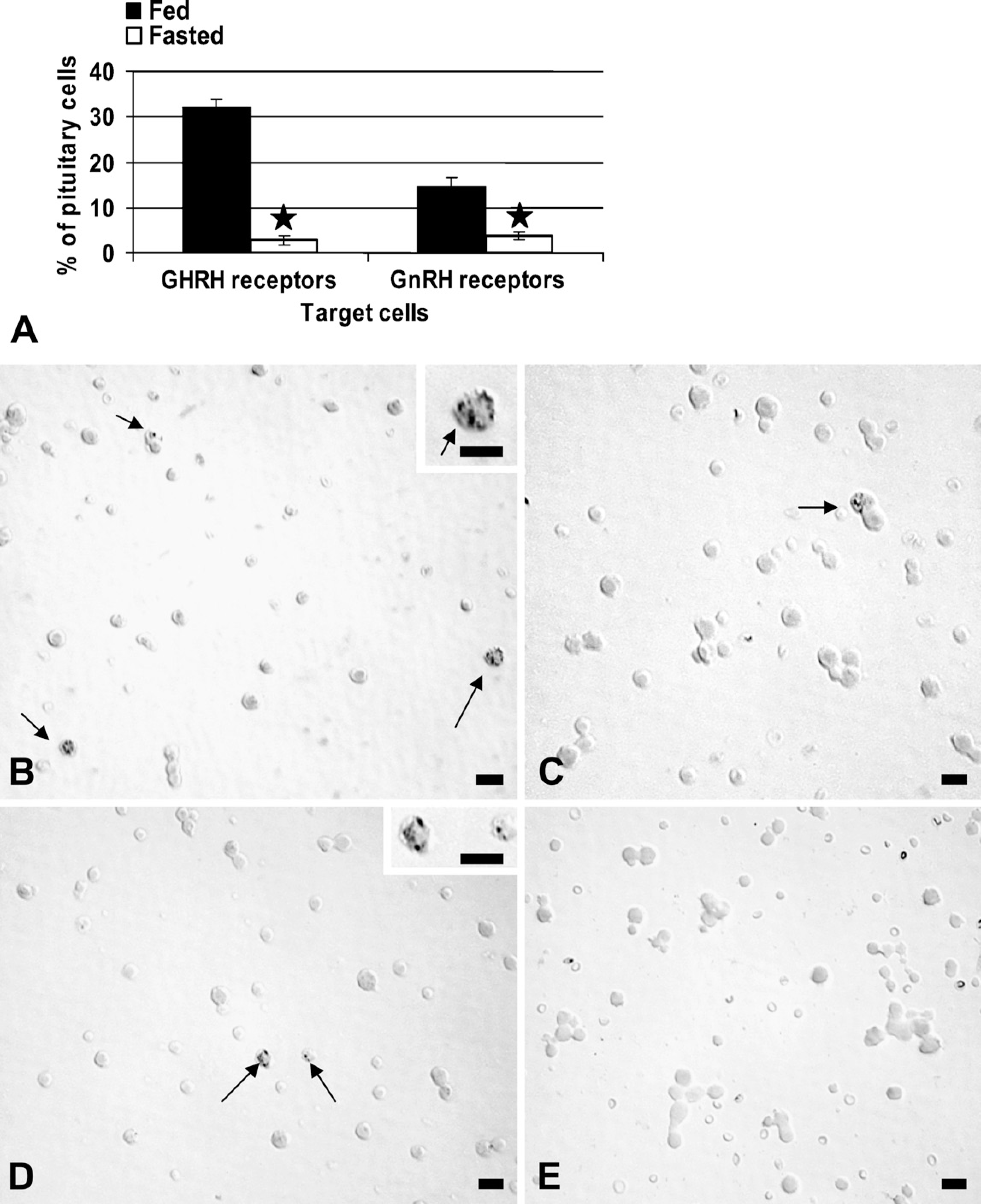
Living, freshly collected AP cells were plated for 45 min and exposed to 1 nM biotinylated gonadotropin-releasing hormone (GnRH) or growth hormone–releasing hormone (GHRH) for 10 min. After fixation, the biotinylated analogs were detected at their binding sites by avidin-biotin complex, and counts of labeled target cells were then performed. (
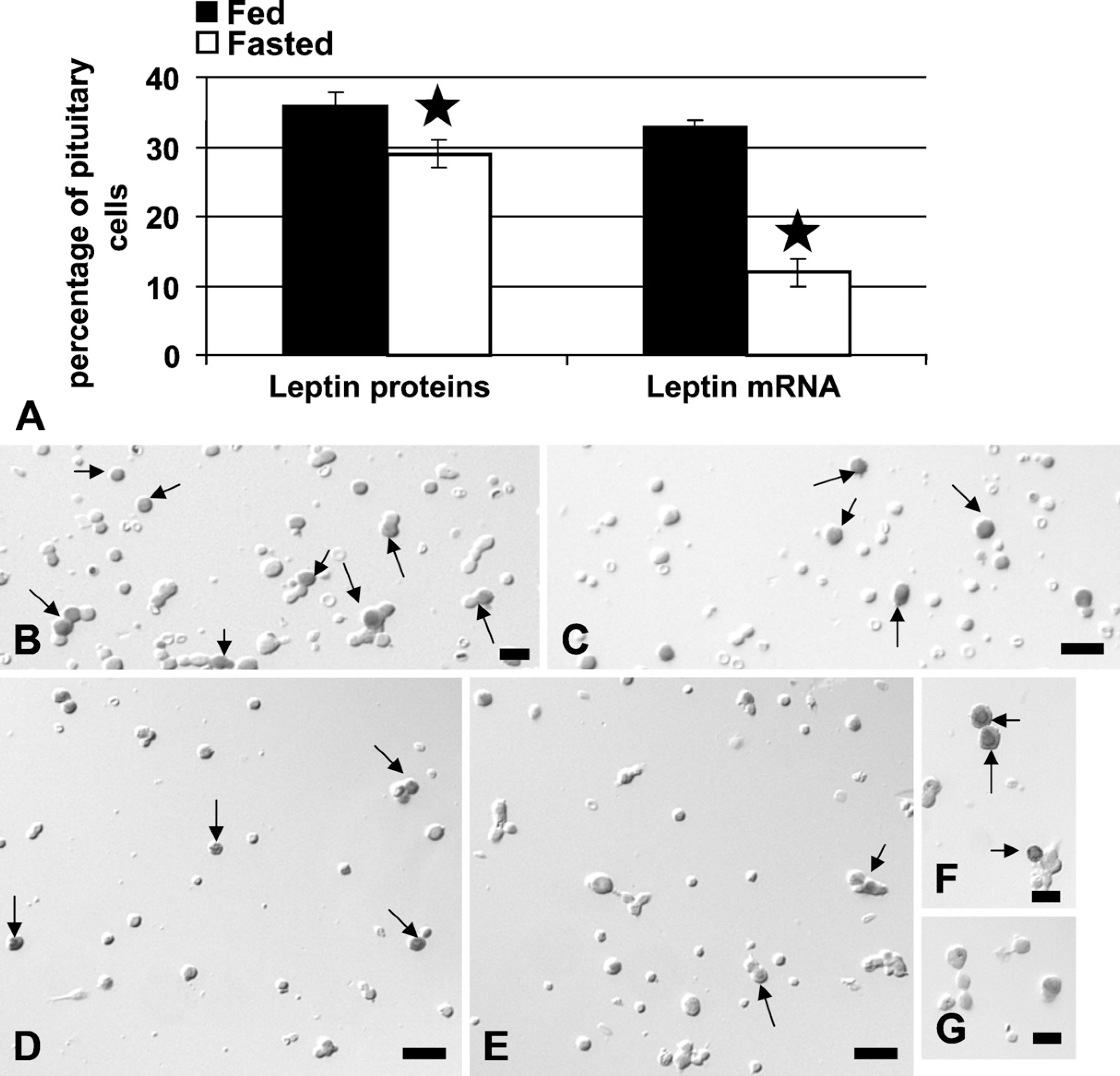
Analysis of the expression of pituitary leptin after 24 hr of food deprivation. (
Effect of Fasting on Differential Expression of Leptin by AP Cells
The next objective of these studies was to learn which pituitary cell types were most affected by the food deprivation, specifically in their production of leptin. Figure 4 illustrates the changes in the percentages of cells with leptin and each pituitary hormone in the AP. In normal, fed populations, cells that coexpress leptin and GH are 22% of the AP population, and this subset represents 84% of GH cells; cells that coexpress leptin with LH, TSH, or ACTH are 2–3% of the pituitary population (20–30% of each of these cell types); those with leptin and prolactin constitute only 1% of pituitary cells or 0.6% of prolactin cells.
When the analysis focused on the population of leptin-bearing cells, the dual labeling accounted for over 90% of cells with leptin proteins, with 61% of leptin-bearing cells coexpressing GH, 8–10% coexpressing LH, TSH, or ACTH, and only 3% coexpressing prolactin. The remaining 10% could be folliculostellate cells, or monohormonal FSHβ cells, which were not detected in these dual-labeling studies.
Figure 4A also shows that food deprivation brought about major losses in leptin protein expression by somatotropes or gonadotropes, reducing the percentages of AP cells with leptin and GH from 22% to 13% and those with leptin and LH from 3% to 1% (p<0.001). In contrast, corticotropes maintained their expression of leptin at 30% of ACTH cells. The overall increase in corticotropes resulted in a 1.94-fold increase in AP cells with leptin and ACTH, from 3.6% to 7% of AP cells (p=0.03). Figure 4 illustrates this increased expression of leptin proteins in corticotropes. The analysis of leptin in fasted rat pituitary populations accounted for 86% of the leptin cell population, with 45% of leptin cells storing GH (reduced from 61%) and 24% storing ACTH (increased from 8%).
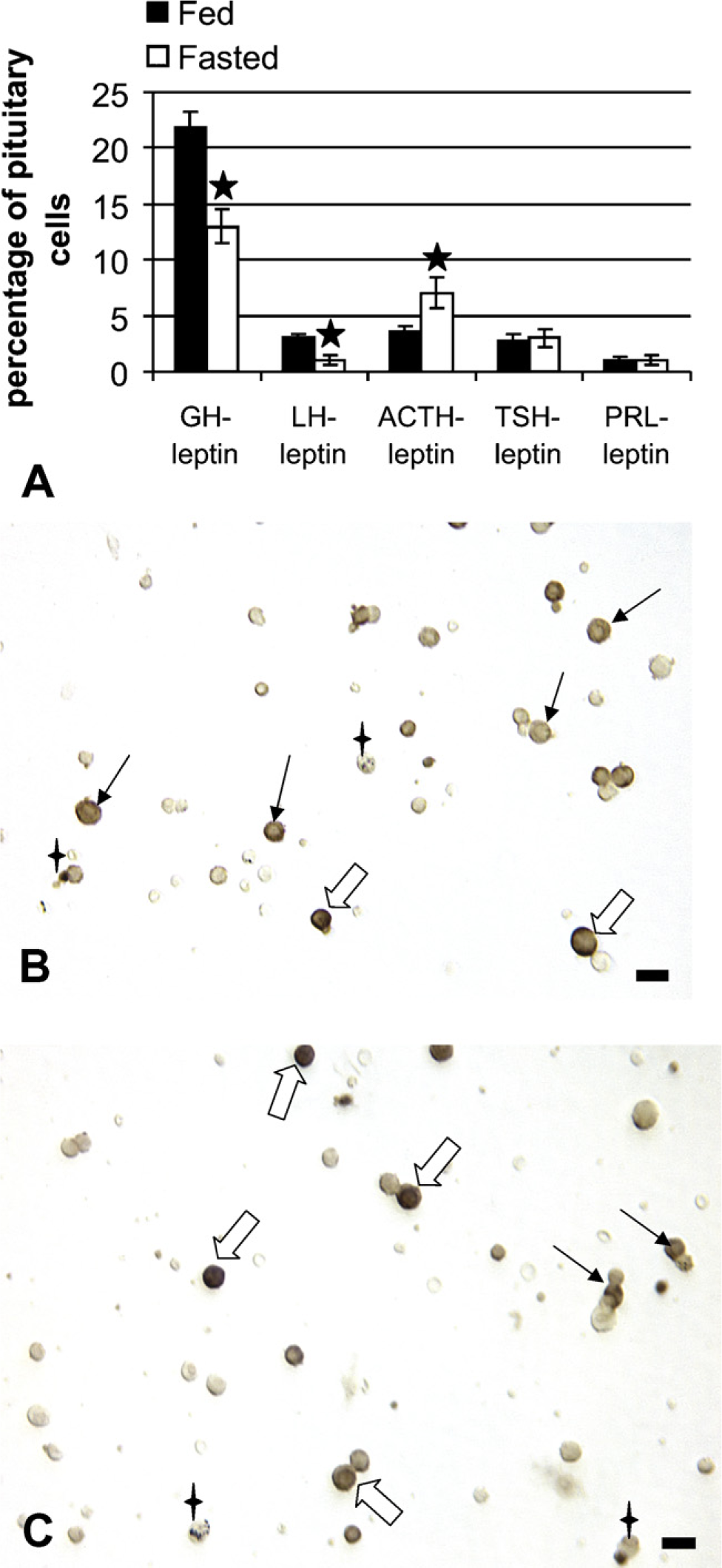
Dual labeling for leptin proteins and each of the pituitary hormones was done on freshly dispersed pituitary cells. (
Because gonadotropes and somatotropes were most severely affected by the food deprivation, the studies then determined whether the reduced leptin mRNA was seen in these populations. Figure 5 shows an 80% decline in cells that coexpressed leptin mRNA and GH proteins, from 25 ± 3% to 5 ± 2% of AP cells (p<0.014). Similarly, fasting caused a significant decline in the coexpression of leptin mRNA in gonadotropes, from 3 ± 0.5% to 0.7 ± 0.02% of AP cells (p<0.029). The sum of the losses in gonadotropes and somatotropes accounted for the overall loss in leptin mRNA. Figures 5B and 5C illustrate the dual labeling for leptin mRNA and GH in fed and fasted rat populations.
Effects of Glucose on Pituitary Leptin-bearing Cells In Vivo
After the parallel fasting–induced reductions in leptin, LH, and GH were detected, the experiments were expanded to test the hypothesis that nutrients such as glucose might regulate this system. This involved the addition of a third set of fasted animals provided with 10% glucose water during the period of food deprivation. Table 1 shows data from three groups of rats (three rats/group). Both groups of fasted rats lost weight, but serum glucose was restored in the animals given 10% glucose water. Figure 6A shows that, in this group of fasted rats, serum leptin was reduced to 26% (p<0.001) of levels in fed rats. Rats given 10% glucose water showed increased leptin, partially restoring it to levels that were 46% (p<0.008) of those in fed rats and significantly higher (p=0.01) than those in fasted rats without glucose.
Whole pituitaries from three separate sets of animals were committed to assays for mRNA by QRT-PCR, to obtain the entire pituitary for the mRNA extracts. This precluded any sampling errors that might be caused by regional differences in the pituitary. Figure 6B shows that leptin mRNA detected by QRT-PCR was reduced in the fasted animals (p<0.026) and restored in the fasted animals given 10% glucose water (p<0.03). The data are the ratio of the number of leptin transcripts to the number of transcripts of the Hprt housekeeping gene. (A similar result was seen if Rps9 was used as the housekeeping gene; data not shown.) Glucose also restored leptin mRNA, detected by in situ hybridization (p<0.001 fasted vs glucose-fasted) (Figure 6C). Figure 6C also shows a parallel restoration of LH mRNA, as detected by in situ hybridization and counts of mRNA-bearing cells (p=0.02 fed vs fasted and p=0.004 fasted vs glucose-fasted). QRT-PCR assays of GH mRNA showed that there were no significant changes with fasting or with glucose water (data not shown).
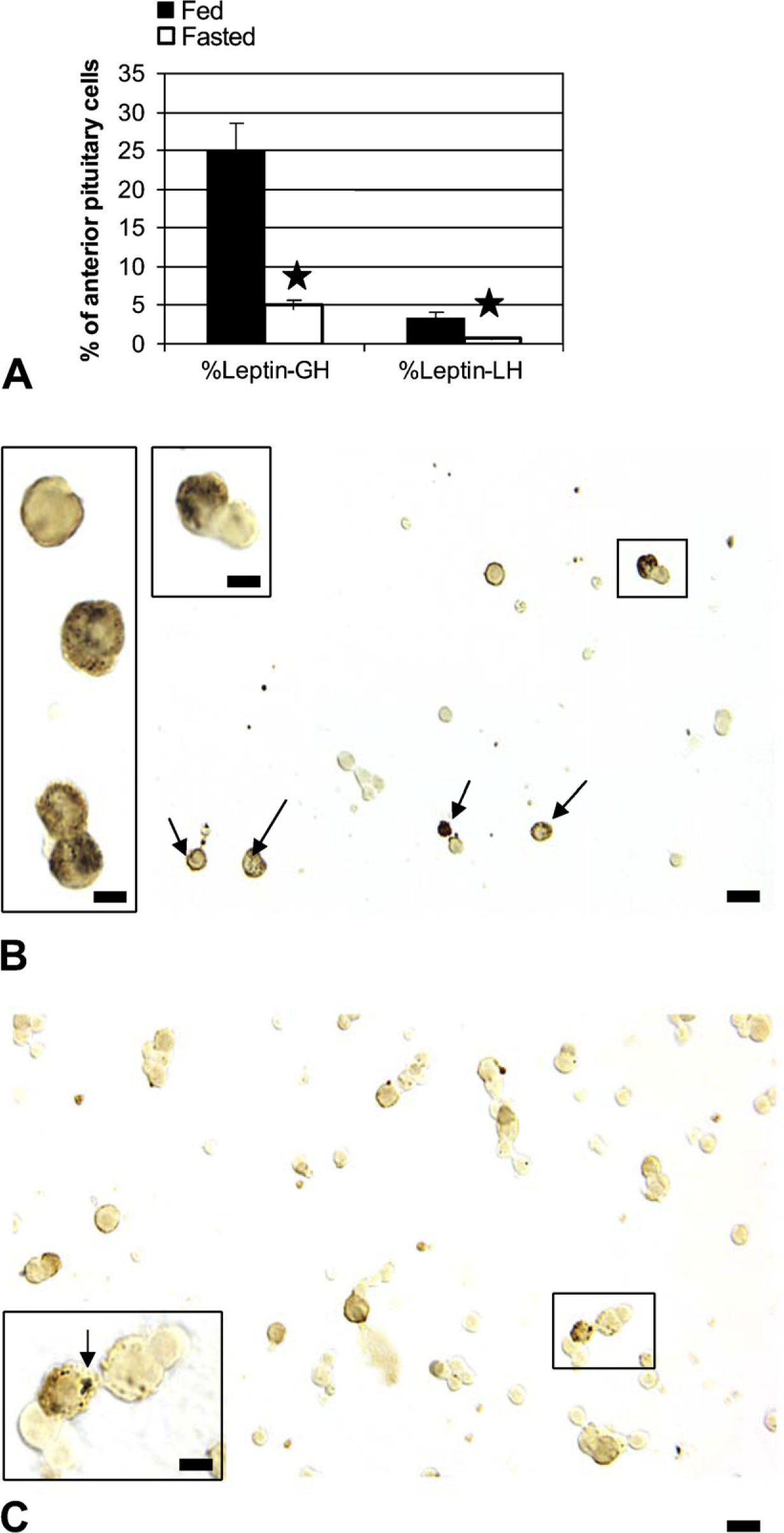
Dual labeling for leptin mRNA and GH or LH antigens was done to learn whether the losses were focused in the somatotrope or gonadotrope population. (
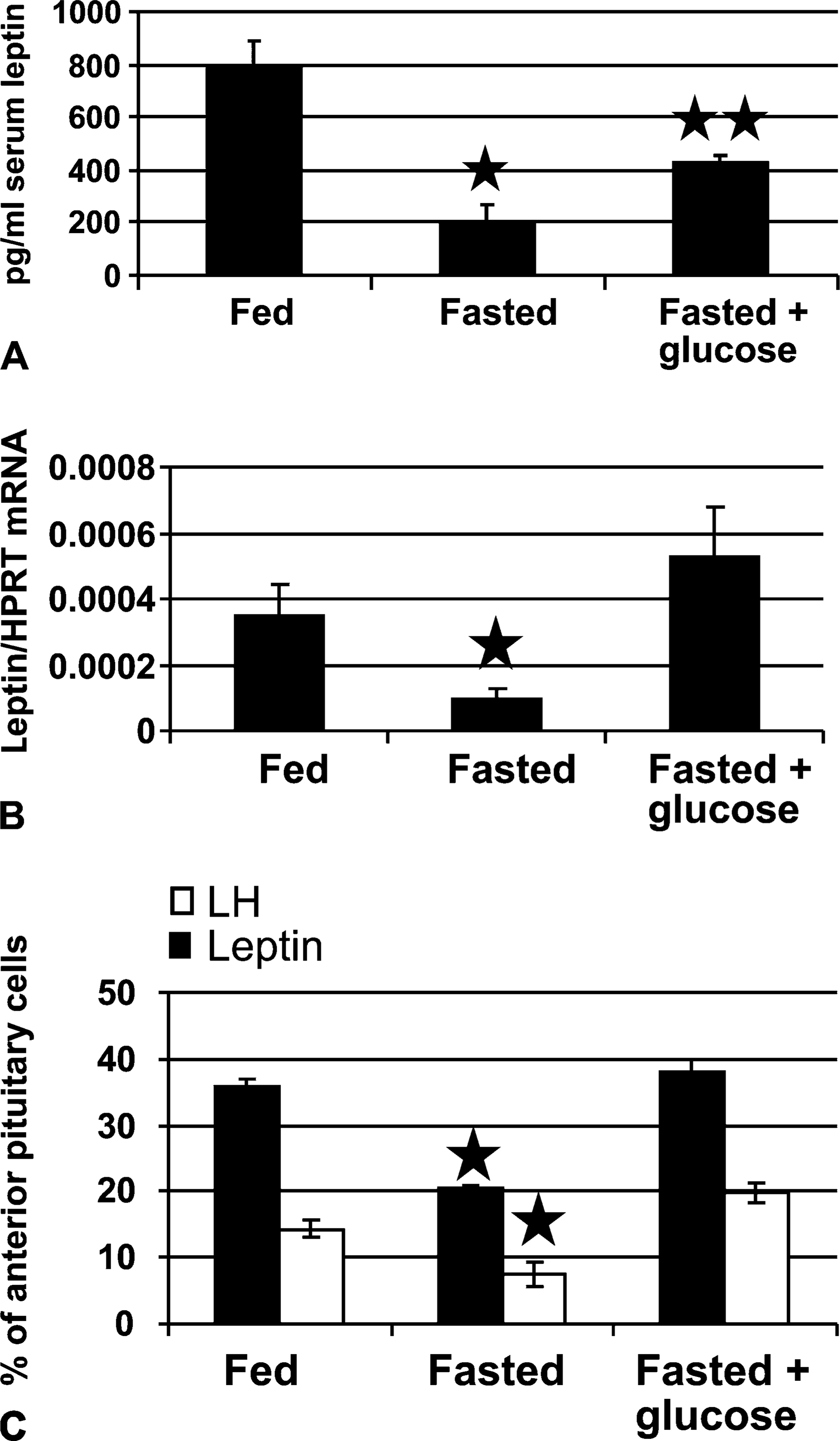
Tests of a second group of fasted rats that received 10% glucose water during the 24-hr period of food deprivation. (
Finally, when additional sets of fresh cultures of pituitary cells were committed to immunolabeling for leptin, LH, and GH, glucose also restored expression of all three protein hormones, as detected by cell counts. Figure 7A illustrates this restoration graphically. Note that the fasting-mediated reduction in leptin-bearing cells in these groups of rats is more dramatic than in the first group (Figure 3A). Figures 7B–7E depict immunolabeled cultures from fasted rats with and without glucose water, showing the restoration of somatotropes (Figures 7B and 7D) and gonadotropes (Figures 7C and 7E) in the population from fasted rats given 10% glucose.
Effects of Leptin on Pituitary Cells In Vitro
The final set of experiments tested the efficacy of leptin in vitro on the possible restoration of GH and LH hormone expression in three groups of freshly dispersed cells from additional fasted animals (six rats total). Leptin was added for 1 hr, with a concentration range (10–100 pg/ml) that matched those assayed in pituitary cell culture media (Akhter et al. 2007). Figure 7A shows that as little as 10 pg/ml leptin for 1 hr restored the percentages of LH and GH hormone-bearing cells to values not different from those in cultures from fed or fasted rats treated with glucose in vivo. Exogenous leptin for 1 hr partially restored the percentages of leptin-bearing cells, with values midway between those from fed rats and fasted rats. Figures 7F and 7G illustrate the restoration of GH and LH cells in the fasted groups treated with leptin in vitro.
Discussion
Development of the Short-term Fasting Model
The overall objective of our ongoing studies has been to determine the physiological significance of pituitary leptin. This is difficult to do in vivo because of the abundance of circulating leptin from adipocytes and the presence of both leptin and leptin receptors in all pituitary cell types. Animal models with leptin or leptin receptor deficiencies have other confounding variables, like hyperglycemia and diabetes, which affect pituitary cells independently. In addition, these models are deficient in gonadotropes and somatotropes and do not respond normally to tests like fasting (Hardie et al. 1996).
Selective knockouts in the pituitary would be needed to fully test the significance of leptin. While these are being developed, this in vivo study was designed to test conditions that might change pituitary leptin in normal rats. The present report is the first to show that pituitary leptin mRNA and proteins are reduced 24 hr after fasting in directions similar to those seen in adipocytes (Dallman et al. 1999). As the model was developed, we recognized that if serum leptin was also reduced, we could not discriminate between effects of pituitary leptin and serum leptin. However, the first two groups of male Sprague-Dawley rats used in our study did not show lower serum leptin after the short 24-hr fasting period. Thus, the timing of the decrease in pituitary leptin mRNA and proteins could be correlated with changes in the different pituitary cell types to begin to develop a hypothesis about a role for pituitary leptin.
The first set of results with fasted animals showed that somatotropes and gonadotropes were most severely reduced. The losses in GH or LH proteins, LH mRNA, and GnRH or GHRH receptor-expressing cells correlate well with the absence of GH or LH pulses during food deprivation described by others (Ahima et al. 1996; Finn et al. 1998; Nagatani et al. 1998, 2000; Schneider et al. 1998, 2000, 2002; Vuagnat et al. 1998; Gonzalez et al. 1999; Pombo et al. 1999; Schneider and Zhou 1999; Maciel et al. 2004). In addition, this study is the first to show selective losses of leptin mRNA and proteins in gonadotropes, somatotropes, and lactotropes with fasting.
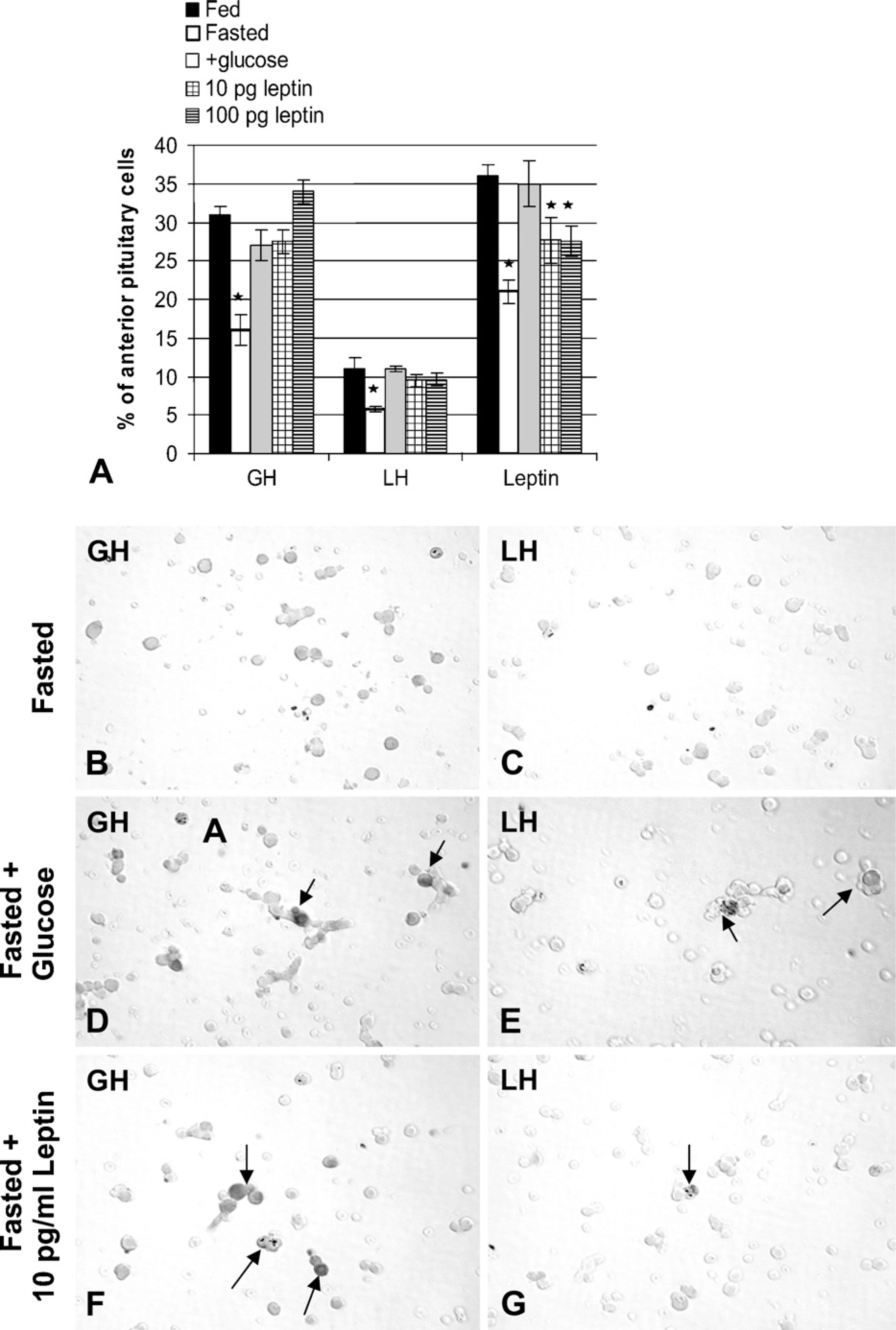
Counts of cells with leptin, GH, and LH proteins labeled by immunocytochemistry. (
Whereas we detected no changes in serum leptin in the pilot studies, the most recent groups that included tests of glucose showed a 74% decrease in leptin. This agrees with most previous studies of fasted or food-deprived animals that found reductions in serum leptin; however, they tested periods longer than 24 hr (Ahima et al. 1996; Casanueva and Dieguez 1999; Dallman et al. 1999; Schneider and Zhou 1999; Schneider et al. 2000, 2002; Maciel et al. 2004; Luque et al. 2007). Fasting-mediated reductions in serum leptin after 24 hr were reported, however, in lean Zucker rats (Hardie et al. 1996), or sham, brain-lesioned female Sprague-Dawley rats (Suga et al. 1999). In both of these studies, the control rats had higher resting levels of leptin (4 ng/ml) than were found in our Sprague-Dawley rats (2 ng/ml). The differences in resting leptin could relate to the differences in the strain, gender, or in the latter study (Suga et al. 1999), to the stress of sham surgery. We have no explanation for the differences between our early and recent groups.
Responses by Pituitary Cells to Fasting
As stated above, the 24-hr fast produced reductions in somatotrope and gonadotrope functions, and at the same time, the short-term fast resulted in a nearly 2fold increase in corticotropes. This increase correlates well with previous reports that show higher spikes of ACTH secretion during the first 24 hr of food deprivation (Akana et al. 1994). The mechanisms underlying this increase could also stem from the increased corticotrophin-releasing hormone mRNA assayed 24 hr after fasting (Luque et al. 2007). At this point, it is not clear whether this increase is from mitotic activity (Childs et al. 1995) or from the differentiation of corticotrope precursors; however, the increase is similar to that seen following a period of acute stress (Childs 1992).
The Fasting Model Tests the Significance of Pituitary Leptin
The fact that the fasting-mediated reduction in pituitary leptin mRNA and proteins occurred rapidly implicates it as a potential regulator in the early responses to fasting. If pituitary leptin functions as a metabolic signal for nutritional changes, the findings in the present study favor this source because of its rapid responses. In addition, the dual-labeling evidence shows that the reduction in leptin following food deprivation is mostly in somatotropes. Perhaps somatotrope leptin is uniquely responsive to changes in the nutritional state. Future studies of somatotrope regulators (including Ghrelin and GHRH) will be needed to provide more clues about regulators for this response.
In contrast, leptin was not reduced in the expanding population of corticotropes. This finding suggests that the regulation of the leptin gene is not uniform across all pituitary cell types. It should be noted that the corticotrope expansion added only 4% leptin-bearing cells to the population, whereas over 10–15% leptin-bearing cells were lost from somatotropes. Thus, assuming the new corticotropes could secrete their leptin in this environment, it was clearly not sufficient to correct for the losses in somatotropes and gonadotropes.
However, the losses in somatotropes or gonadotropes could be corrected rapidly by a 1-hr incubation in as little as 10 pg/ml exogenous leptin. These levels match the levels secreted by normal rat pituitary cultures. Higher levels (100 pg/ml) did not cause further increases in LH or GH cells. [Note: A full set of dose-response tests in normal rat pituitary cells (1 pg/ml to 1 ng/ml) showed that 10 pg/ml was a plateau point for increases in GH cells and 100 pg/ml was the plateau point for increases in LH cells.] Many previous studies have demonstrated exogenous leptin restoration of LH or GH pulses in vivo (Ahima et al. 1996; Finn et al. 1998; Vuagnat et al. 1998; Gonzalez et al. 1999; Pombo et al. 1999; Nagatani et al. 2000), with the assumption that its actions are mainly on the hypothalamus. Our studies suggest that leptin may also act directly on pituitary target cells and that restoration is not limited to actions on the hypothalamus.
Exogenous leptin was not able to fully restore the expression of pituitary leptin, although there was a partial recovery with 100 pg/ml leptin. This result points to independent regulators for pituitary leptin, and our previous studies showed that they include estrogen, GHRH (McDuffie et al. 2004), and GnRH (Akhter et al. 2007). These regulators would be reduced in a fasted state (Ahima et al. 1996; Finn et al. 1998; Vuagnat et al. 1998; Gonzalez et al. 1999; Pombo et al. 1999; Nagatani et al. 2000; Maciel et al. 2004; Luque et al. 2007).
The parallel restoration of leptin and LH mRNA, or leptin, LH, and GH proteins in the glucose-treated rats provided circumstantial evidence that further supports an association between these gene products in the pituitary. Food deprivation (signaled by a drop in nutrients like glucose) is known to result in the attenuation of LH or GH pulses, and it is thought that some of this is signaled by a drop in serum leptin. In vivo studies have shown that the LH pulses can be restored within hours of giving nutrition. It can also be restored in vivo in fasted animals by exogenous leptin as long as glucose can be utilized (Schneider and Zhou 1999; Schneider et al. 2000, 2002). Thus, workers have theorized that leptin reports nutritional information to the hypothalamic and pituitary cells, permitting reproduction and normal GH or LH cell pulses if nutrition is adequate.
However, this present study reports new and unexpected findings, in which gonadotropes and somatotropes show deficiencies in response to food deprivation in the face of a decline in pituitary leptin. Perhaps pituitary leptin is needed to maintain LH and GH cell functions. If so, our dual-labeling studies suggest that important sources would be the somatotropes or gonadotropes themselves. These findings lead us to hypothesize that a decline in somatotrope or gonadotrope leptin below certain threshold levels might signal nutritional distress and cause reductions in LH or GH cell functions. Endogneous pituitary leptin could thus serve as either a paracrine regulator or an autocrine regulator.
The studies of glucose restoration confirmed this hypothesis, because they showed parallel increases in pituitary leptin, GH, and LH once serum glucose was restored to normal levels. Serum leptin was only partially restored (Figure 6A) while pituitary leptin was completely recovered (Figure 7A). This result suggests that pituitary leptin might serve as a glucostat, which would provide one mechanism by which it senses the nutritional state. Recent landmark studies (Zelent et al. 2006; Sorenson et al. 2007) reported glucokinase expression in a subset of pituitary cells, including thyrotropes and gonadotropes. This important glucose sensor may allow gonadotropes to monitor the changes in serum glucose. Our studies suggest that leptin may also be affected by this sensor and may play a role in facilitating gonadotrope responses.
Finally, as stated in the beginning of this discussion, we recognize that the evidence for the involvement of pituitary leptin in the regulation of LH and GH in this in vivo model is circumstantial at this point. Selective knockouts that remove leptin or its receptor from the pituitary would be needed to test the hypothesis about pituitary leptin directly. However, the present studies provide an important basis for such tests. They also add to the growing body of evidence that suggests local roles for leptin as an important cytokine in multiple organs and systems. Finally, the differential responses to fasting by corticotropes and somatotropes indicate that any future studies of pituitary leptin must recognize that overall changes in pituitary leptin reflect the net result of changes in diverse AP cell types. They may not represent the unique responses shown by each of the cell types.
Footnotes
Acknowledgements
This publication was made possible by studies funded by NSF IBN 0240907, NIH R03 HD 44875, and 1 P20 RR020146 from the National Center for Research Resources (NCRR), a component of the National Institutes of Health (NIH). Its contents are solely the responsibility of the authors and do not necessarily represent the official views of the NCRR or the NIH.
The authors thank the National Institute of Diabetes and Digestive and Kidney Diseases Hormone Distribution office and Dr. A. Parlow for the anti-rat GH, TSHβ, FSHβ, prolactin, and anti-rat/mouse leptin serum. They also thank J. G. Pierce, PhD, for the anti-bovine LHβ. The authors appreciate the help and advice of Paul Hughes of ![]() in the design, preparation, and application of the oligonucleotide probes. This study was submitted in partial fulfillment for the degree of master of science by C.C. It was presented as a poster at the 89th Annual Meeting of the Endocrine Society, Toronto, Canada, June 2007.
in the design, preparation, and application of the oligonucleotide probes. This study was submitted in partial fulfillment for the degree of master of science by C.C. It was presented as a poster at the 89th Annual Meeting of the Endocrine Society, Toronto, Canada, June 2007.