
Abstract
We investigated the mechanism by which diabetes renders the capillary endothelium more permeable to macromolecules in the lungs of short-term diabetic rats. We used quantitative immunocytochemistry (ICC) to comparatively assess the permeability of alveolar capillaries to serum albumin in diabetic and normoglycemic animals. The effect of diabetes on the population of endothelial caveolae was evaluated by morphometry and by ICC and immunochemical quantification of the amount of caveolin in the whole cell or associated with the purified endothelial plasma membrane. A net increase in the amount of serum albumin taken up by the plasmalemmal vesicles of alveolar endothelial cells and transported to the interstitium was documented in diabetic animals. Interendothelial junctions were not permeated by albumin molecules. The alveolar endothelial cells of hyperglycemic rats contain more caveolae (1.3-fold), accounting for a larger (1.5-fold) fraction of the endothelial volume than those of normal animals. The hypertrophy of the caveolar compartment is accompanied by overexpression of endothelial caveolin 1. Although the aggregated thickness of the endothelial and alveolar epithelium basement membranes increases in diabetes (1.3-fold), the porosity of this structure appears to be unchanged. Capillary hyperpermeability to plasma macromolecules recorded in the early phase of diabetes is explained by an intensification of transendothelial vesicular transport and not by the destabilization of the interendothelial junctions.
The main function of the microvasculature is the exchange of substances between blood and tissue. Because it is directly exposed to the bloodstream, the endothelium represents the element of the vascular wall that performs or controls these exchanges. The only exception is the renal glomerulus, in which the effector is not the endothelial cell per se but its underlying product, the basement membrane.
The morbidity and mortality caused by diabetes mellitus are mainly related to vascular complications (Turner 1998). Whereas at the level of large vessels the disease appears to be expressed as an acceleration of an atherosclerotic process (Lopez-Virella 2001), the microangiopathy shows a more diverse picture, with specific facets for each affected organ (Libby 1997; Naruse and King 2001). Because of their dominant clinical incidence and pattern, the diabetes-induced alterations in the microvasculature of three vascular beds (retina, renal glomerulus, and nerves) have been the focus of a vast number of studies. From these we have learned that the direction in which the diabetic microangiopathy develops and the sequence of events vary among these vascular beds but that several elements, such as an increase in capillary permeability and the thickness of the endothelial basement membrane, are common to all three (Marre 1997). A continuously increasing number of investigations show that, even in the absence of a significant clinical impact, diabetes does affect the microvasculature of other organs, such as the mesentery (Arshi et al. 2000), the skin (Oomen et al. 1999), the skeletal muscle (Beals et al. 1993), and the brain (Bouchard et al. 2002). In these vascular beds, changes in vascular permeability appear to represent a hallmark of the diabetic endothelial dysfunction.
The mechanism by which such alterations take place is not precisely known. Our study aims to investigate this aspect by correlating the modifications induced by diabetes in the transendothelial transport of macromolecules to the alterations of the subcellular endothelial vesicular compartment as revealed by the biochemical marker for the latter, i.e., caveolin. This was done according to the following experimental strategy. The amount of caveolin 1 present in the alveolar endothelium of normal and diabetic rats was comparatively assessed by immunoblotting performed on the purified luminal aspect of lung endothelial plasma membranes. The presence of the same protein was assessed independently by quantitative immuno-cytochemistry performed either before embedding on purified membranes or after embedding on the entire endothelial cells. The correlation between the expression of caveolin 1 and the population of plasmalemmal vesicles was investigated morphometrically by counting the number of endothelial caveolae and measuring the partial cell volume occupied by them. The functional implication of the recorded variation in the numerical density of the plasmalemmal vesicles was further tested in relation to the endothelial permeability to macromolecules. This was achieved by comparatively measuring, in diabetic and normoglycemic animals, the in situ distribution of either endogenous albumin or its exogenous variant, a hapten-labeled bovine serum albumin, between the vascular lumen and the interstitium.
The results presented below show that, in the early stages of experimentally induced diabetes, the permeability of the lung microvascular bed to albumin increases. This modification is due to a significant hypertrophy of the plasmalemmal vesicle compartment of the alveolar endothelium, accompanied by a net overexpression of caveolin 1.
Materials and Methods
Animal Model
Twenty-nine Sprague-Dawley male rats, 125 g body weight (Charles River; St-Laurent, Quebec, Canada) were used throughout the experiments. Eleven were kept as control and the rest were rendered diabetic by a single IP injection (70 mg/kg bw) of streptozotocin (STZ). Animals were maintained hyperglycemic without insulin treatment for 3 months. They were individually tested for glycemia 48 hr after the STZ injection and just before sacrifice using the GlycoTest II kit (Roche Diagnostics; Laval, Quebec, Canada); glycosuria was monitored weekly during the 3-month interval, using Uriscan (VWR-Canlab; Montréal, Quebec, Canada) strips. Animals had free access to standard diet and water. All the experiments were conducted with the approval of the institutional Committee of Deontology for the Experimentation on Animals.
Purification of the Lung Endothelial Plasma Membrane
The luminal aspect of the endothelial plasma membrane was isolated from the microvascular pulmonary bed of normal and diabetic rats by the cationic colloidal silica method (Jacobson et al. 1992; Ghitescu et al. 1997). This procedure involves perfusion of the lung microvasculature with a suspension of cationic silica nanoparticles, which adhere to the accessible surface (blood front only) of the endothelium. The high density imparted by the colloidal silica to the labeled plasmalemma facilitates purification of the latter by simple centrifugation. The purity of the isolated membrane fractions, operationally labeled P2, was assessed by electron microscopy as previously described (Ghitescu et al. 1997). Each preparation was derived from a single animal and was treated as an independent sample.
Immunochemical Evaluation of Caveolin in Purified Lung Endothelial Plasmalemma
To make a precise appraisal of the protein concentration, the measurements were made in quadruplicate by the Amido black method (Schaffner and Weissmann 1973) in the supernatants of the plasma membrane fractions solubilized in 1% SDS, sonicated, boiled, and microfuged to remove the interfering silica beads. Equal amounts (5 μg) of P2 fractions from diabetic and normal rats were loaded in Laemmli buffer on 10% acrylamide Minigels (BioRad; Mississauga, Ontario, Canada), resolved by electrophoresis, and electro-transferred to nitrocellulose membranes. The membranes were probed overnight with 1:4000 anti-caveolin 1α rabbit polyclonal antibody (Santa Cruz Biotechnology; Santa Cruz, CA), followed for 1 hr by a 1:5000 dilution of anti-rabbit Ig-HRP (Jackson ImmunoResearch; West Grove, PA). The quencher, washing solutions, and enhanced chemiluminescence (ECL) substrate, as well as the detailed immunoblotting protocol, were those of the Roche Diagnostics BM chemiluminescence blotting substrate kit. The ECL signal was recorded at various exposures on X-ray films and the intensity of the individual bands was measured by densitometry in a BioRad GS800 calibrated densitometer.
Similar detections were performed for two other plasma membrane-associated proteins by using antibodies to a C-terminal epitope common to all three isoforms of actin (product A2066; Sigma Aldrich, St Louis, MO) and to annexin II (clone C-16; Santa Cruz Biotechnology), respectively.
Pre-embedding immunocytochemistry for caveolin was performed on lung endothelial plasma membranes (P2 fractions) from three diabetic and three healthy rats. Freshly purified membranes were lightly fixed for 1 hr in 1% paraformaldehyde in 0.1 M phosphate buffer (PB), then washed and quenched in PB containing 0.1 M glycine. Given the high density of the silica-decorated membranes, low-speed centrifugation (400 × g/min) sufficed to separate them from the different fluids in which they were sequentially dispersed. The membranes were incubated overnight at 4C in a 1:100 dilution of anti-caveolin antibody and washed three times in PB, followed by 1-hr incubation in the cold in 10 nm protein A-gold complexes at a concentration corresponding to A520 = 0.5. After several cycles of resuspension in PB and sedimentation at low speed to remove unattached protein A-gold conjugates, the membranes were tightly packed by microcentrifugation at 12,000 × g for 10 min, fixed again overnight at 4C in a mixture of 2.5% paraformaldehyde and 1.5% glutaraldehyde, postfixed in 1% OsO4, dehydrated, and embedded in Epon. In this experiment and in all ICC approaches further described, the specificity of the labeling was tested in control samples that were not exposed to the primary antibody but only to the protein A-gold conjugate. Three ultrathin sections for each animal, normal or diabetic, provided the 119 images (×31,000 final magnification) used for counting the number of gold particles decorating the unit length of the plasma membrane cytoplasmic face. The morphometry was performed on a Zeiss Videoplan 2 Image Analysis System (Carl Zeiss Canada; Don Mills, Ontario, Canada). The statistical significance of the difference found between the two lots was validated by the unpaired Student's t-test here, as well as in all other subsequent assays.
Postembedding ICC Detection of Caveolin
Six animals (three normal and three diabetic) served as donors for lung tissue samples. Under deep anesthesia, the thorax was opened and the lung was fixed in situ for 10 min with warm 4% paraformaldehyde. Small blocks of tissue were harvested from the same anatomic site (periphery of the lower left lobe), and further fixed for another hour at 4C. The tissue was rinsed in PB and embedded at low temperature in Lowicryl K4M as previously described (Bendayan 1984). For each animal, three ultrathin sections, each from a different block, mounted on Parlodion-coated nickel grids were sequentially incubated in 1% ovalbumin in PBS for 10 min, a 1:100 dilution of anti-caveolin 1α antibody (2 hr) followed by 10 nm protein A-gold conjugated (30 min) with extensive PBS washing in between. The intensity of the labeling was measured over the entire profile of endothelial cell cytoplasm and was expressed as number of gold particles per unit area. The alveolar endothelium of the rodent lung exhibits zones of extremely attenuated cell body (down to 0.1 μm) that do not contain any vesicles or organelles (Simionescu and Simionescu 1983). These “avesicular zones,” as well as those containing the nuclei, were excluded from the morphometrical evaluation.
Morphometry of the caveolar compartment of the alveolar endothelial cells was also performed on three diabetic and three age-matched normoglycemic rats. The lung tissue was prepared as described above, except that, to improve the visibility of the vesicular compartment, a mixture of 1.5% glutaraldehyde and 2.5% paraformaldehyde was used for the primary fixation and a postfixation step in 1% OsO4 was added. The lung specimens were embedded in Epon and three ultrathin sections from different blocks were examined for each animal. The numbers of plasmalemmal vesicles (caveolae) per unit surface of endothelial cell profile, as well as the total area of these vesicles, were measured by planimetry for each of the 188 images (×55,000 final magnification) taken into consideration. The first parameter represents a measure of the numerical density of the endothelial plasmalemmal vesicles. For each micrograph, the ratio between the aggregated surface of sectioned caveolae and the corresponding endothelial profile was calculated. This parameter gives an estimate of the partial volume occupied by the caveolae in endothelial cells.
Transendothelial Distribution of Endogenous Albumin
The same mildly fixed, low-temperature Lowicryl K4M-embedded lung tissue blocks, previously used for the post-embedding ICC detection of caveolin, also served for the experiment in which the steady-state distribution of the endogenous albumin across the endothelium was monitored with an anti-rat albumin antibody and protein A-gold. The ultrathin sections, mounted on grids, were sequentially incubated (30 min, room temperature) in PBS containing 150 mM glycine and 1% fish gelatin, a 1:100 dilution of anti-rat serum albumin antibody (Cappel; West Chester, PA) in PBS (overnight at 4C), followed by 10-nm protein A-gold conjugates (30 min, RT) with thorough washing in PBS in between. The intensity of labeling (number of gold particles per unit surface) was counted by planimetry (Ghitescu and Bendayan 1992) over the endothelial cell and basement membrane profiles present in the 215 images taken at ×38,000 final magnification. The specificity of the approach was assessed by omitting the anti-albumin antibody in the labeling protocol and incubating the grids in a suspension of protein A-gold only.
Evaluation of the Transcytosis of Dinitrophenylated Albumin (DNP-BSA)
An exogenous tracer for transcytosis, bovine serum albumin (BSA), was tagged with dinitrophenol (Ghitescu and Bendayan 1992); 150 mg of this marker in 1.5 ml PBS at 37C was injected into the bloodstream of three diabetic and three normal rats. Before the injection, a volume of blood identical to that of the tracer was withdrawn from the jugular vein. After a delay of 30 min, the animals were sacrificed by cervical dislocation and a warm solution of 1.5% glutaraldehyde, 2.5% paraformaldehyde in 0.1 M PB was injected into the thoracic cavity to rapidly fix the lung in situ. After 5 min the lung was harvested, and small pieces from the periphery of the lower left lobe were further fixed overnight in the same mixture of aldehydes. Postfixation in 1% OsO4, dehydration in graded ethanol, and embedding in Epon followed. For each animal, three grids were prepared and immunolabeled with anti-DNP antibody followed by protein A-gold, as described elsewhere (Ghitescu and Bendayan 1992). The intensity of labeling for the exogenous tracer was measured over the capillary lumen, within the endothelial cell contour, and over the endothelial basement membrane, and was expressed as number of gold particles per surface unit. The “porosity” of the endothelial basement membrane was evaluated according to an established test (Bendayan et al. 1986), in which the position of each gold particle detected over this structure was defined by the ratio between the distance separating the particle from the abluminal front of the endothelium and the whole thickness of the basal lamina measured at that location.
Results
The two lots of animals used throughout this study (diabetic and age-matched normal rats) differed significantly in their body weight and glycemia, both measured at the moment of sacrifice (Table 1). As expected, additional signs of diabetic pathology were encountered in the STZ-injected animals: polyuria, cataract, and a significant level of glycation of the intracellular endothelial proteins (Ghitescu et al. 2001).
Characteristics of diabetic and control rats
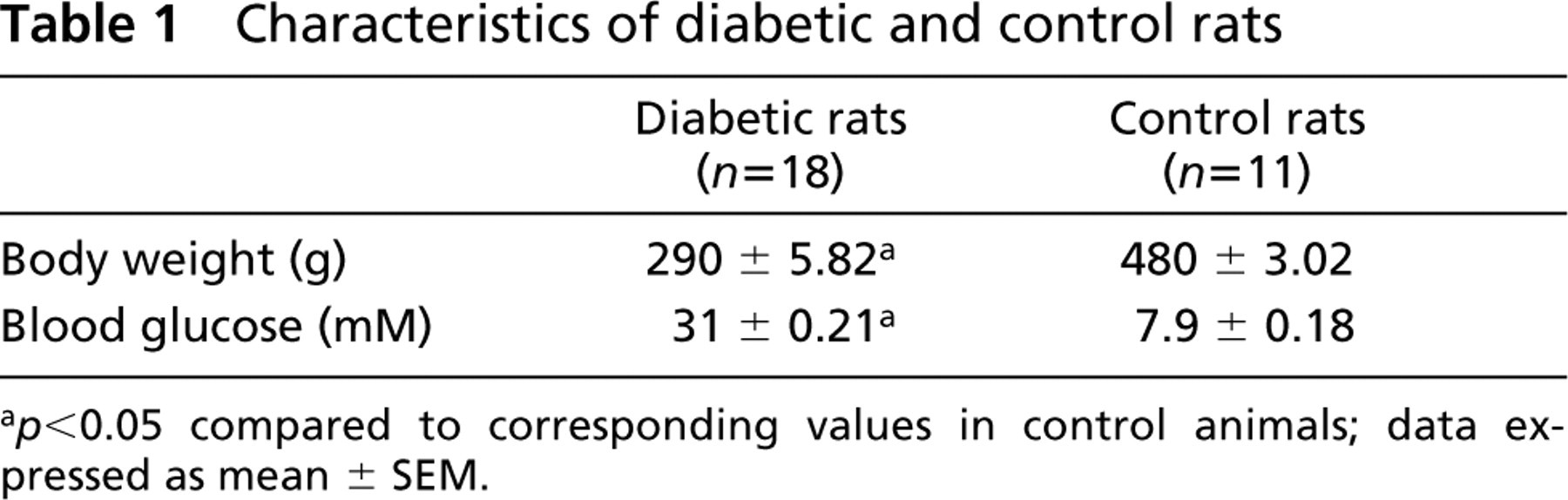
a p < 0.05 compared to corresponding values in control animals; data expressed as mean ± SEM.
Electron microscopic inspection of the endothelial plasma membrane (blood front) purified from the lungs of diabetic and normal rats showed that this fraction, operationally named P2, was virtually devoid of contamination (Figure 1). All the material was present in the form of large sheets, covered on one side by the cationic colloidal silica particles. The silica particles perfused through the vasculature have access and bind solely to the luminal front of the endothelial cells. Their presence on all the membranes found in our preparation is therefore an irrefutable argument for the nature of the material used in the subsequent experiments (Jacobson et al. 1992; Ghitescu et al. 1997).
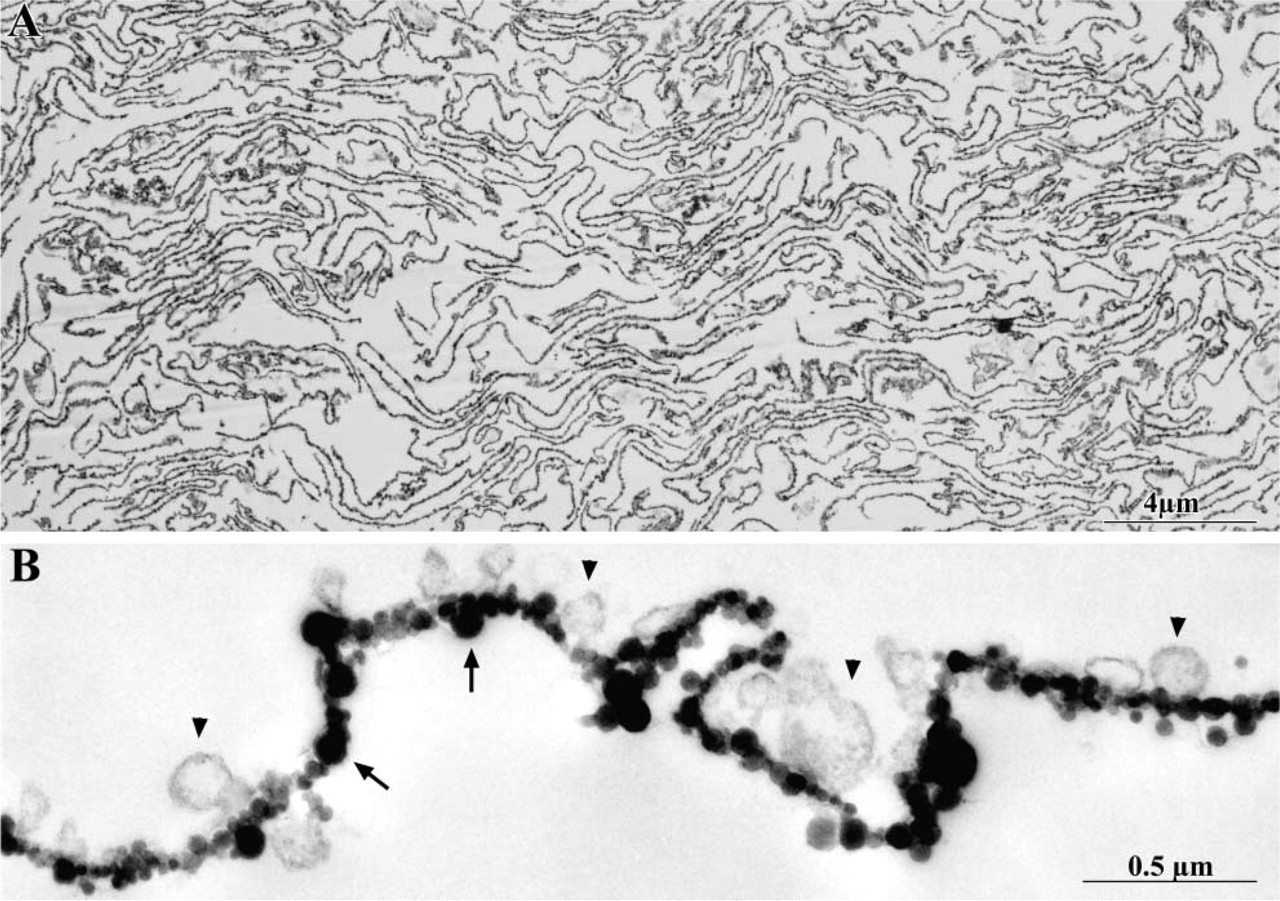
(
Equal amounts of protein of P2 fractions, each originating from an individual healthy or diseased animal, were resolved by electrophoresis and either silver-stained to further assess the uniformity of the loading between lanes (Figure 2A) or electrotransferred and probed with an anti-caveolin 1α antibody (Figure 2B). A single band, whose intensity varied between the samples from the diabetic and normal rats, was revealed at the position corresponding to 21 kD. Densitometry performed on both gel and immunoblot showed that, at an identical protein load (Figure 2C), the intensity of the signal for caveolin 1α in samples from the diabetic rats was 1.9 ± 0.17-fold higher than in the membranes purified from the normal animals (Figure 2D).
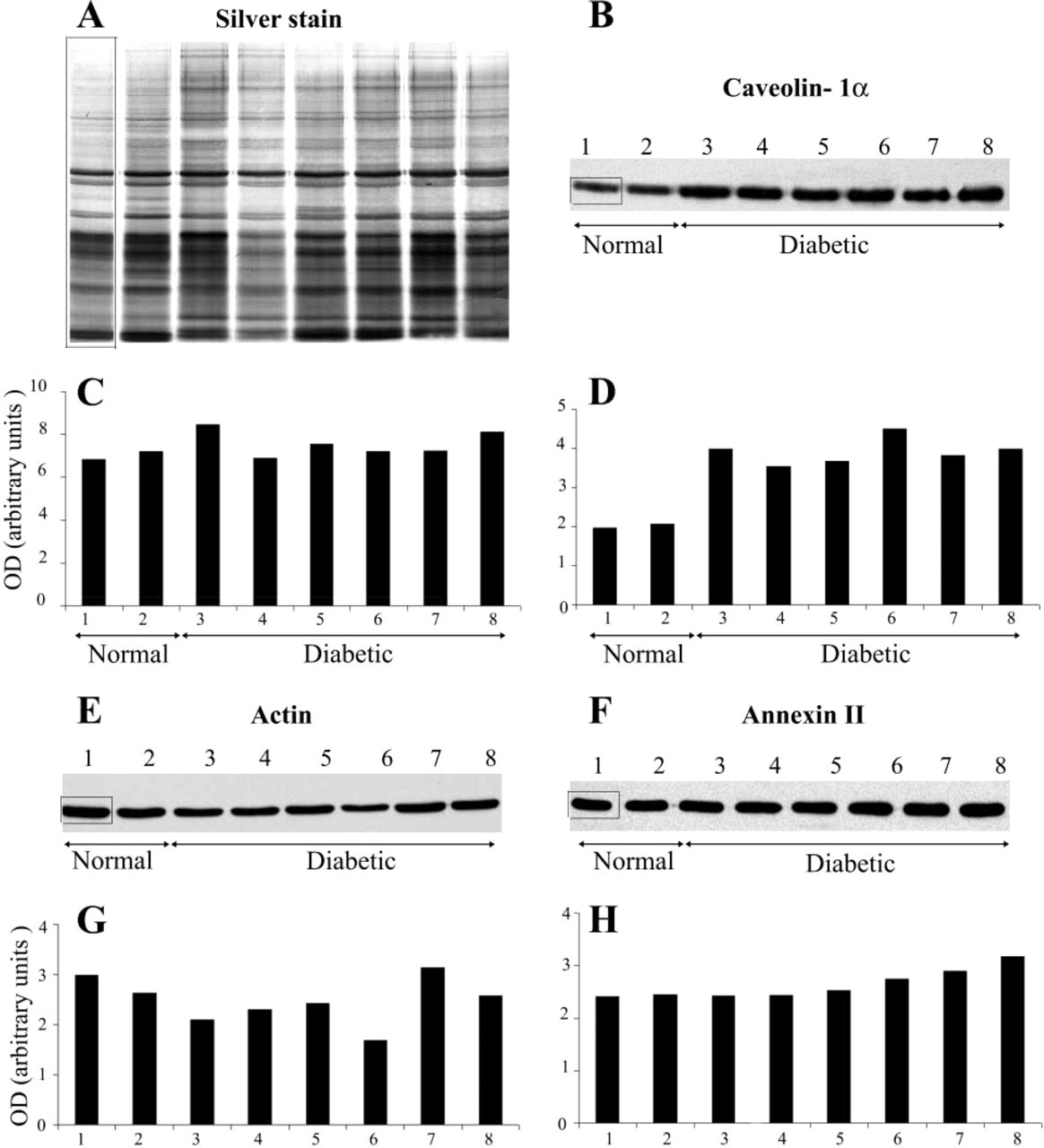
Comparative ICC detection of caveolin 1α, actin, and annexin II in the endothelial membranes of normal (Lanes 1 and 2) and 3-month diabetic (Lanes 3-8) rats. Five μg of each fraction P2 was submitted to electrophoresis and either silver-stained (
When a similar setting was used to detect and quantify comparatively two other proteins, actin and annexin II, which, like caveolin, are also associated with the cytoplasmic face of the endothelial plasmalemma, the data indicated no consistent variations between the normal and diabetic lots (Figures 2E-2H). The ratio normal:diabetic was of 0.86 ± 0.16 for actin and 1.11 ± 0.12 for annexin II.
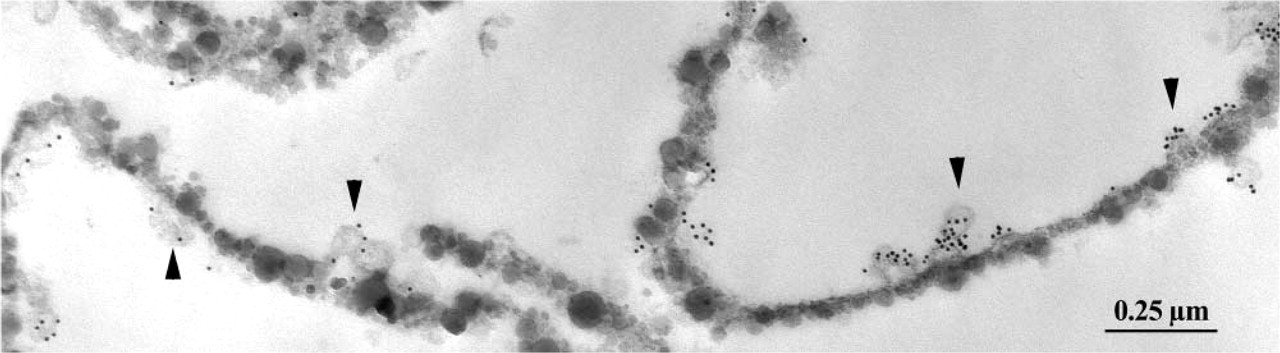
ICC labeling for caveolin 1 of the purified lung endothelial membranes of a diabetic rat. Note the association of the signal (10-nm gold particles) with the identifiable caveolar profiles (arrowheads). Some of the purified membrane sheets twist in the plane of the section, resulted in the false impression that caveolae are present on both their faces. Original magnification ×51,500.
To strengthen these data, a pre-embedding quantitative ICC detection of caveolin was performed on endothelial membranes purified from healthy and diabetic rats. Only the exocytoplasmic aspect of these membranes carries the silica particles. This distribution leaves the cytoplasmic face freely accessible to all elements used for immunodetection, in this case anti-caveolin 1 antibody and protein A-gold conjugates. The vast majority of the gold particles, reporting the presence of caveolin, were observed as clusters around the caveolae associated with the membrane sheets (Figure 3). Membranes that, as a control for the specificity of the immunolabeling, were incubated in protein A-gold only were virtually devoid of gold particles (0.17 ± 0.06 particles/μm; n = 26; 101.5 μm examined membrane length). The density of the labeling for caveolin sharply differed between the experimental lots (Table 2). On average, 1.73-fold more gold particles were found to decorate the cytoplasmic face of the membranes derived from the diabetic animals than those of normoglycemic rats.
Whether the variations in the amounts of caveolin found by the previous assays were not an artifact of plasma membrane purification was further tested by performing postembedding quantitative ICC for this protein on lung tissue fixed in situ. The advantage of this technique resides in the detection of molecules of interest present within the cell body and not only at the cell surface. Labeling of the Lowicryl-embedded lung tissue with anti-caveolin 1α antibody and protein A-gold conjugates consistently produced decoration of the endothelial cells (Figure 4). Given the inherently low contrast of membranes in the Lowicryl-embedded specimens, it is virtually impossible to distinguish all plasmalemmal vesicle profiles. For that reason, the intensity of the signal (number of gold particles/μm2) was quantified over the entire cytoplasm and was found to be on average 1.24-fold higher in diabetic than in normal rats (Table 3). This shows that, in diabetic animals, the alveolar endothelium contains measurably higher amounts of caveolin. Very few gold particles (2.05 ± 0.25 particles/μm2) were located over the nuclei (57.7 μm2 nuclear surface examined; n = 19). Moreover, a low level of nonspecific labeling (0.95 ± 0.22 particles/μm2) was observed over the endothelium in cross-section when the primary antibody was omitted and only the protein A-gold conjugates were used.
By morphometry, we found that, concurrent with the overexpression of caveolin, the plasmalemmal vesicle compartment was significantly more developed in the alveolar endothelium of the diabetic rats. The measurement was performed on lung specimens prepared under conditions (osmium postfixation, Epon-embedding) that maximize the preservation and visibility of the caveolae. The diabetes-induced development of the vesicular compartment is manifest in terms of both numerical density of the vesicles and the fraction of endothelial cell volume occupied by them. According to our measurements, the lung capillaries of normal rats display an average of 37.86 caveolae and plasmalemmal vesicles profiles per μm2 cross-section of endothelial cell (Table 4). This accounts for a partial caveolar volume of 15.5 ± 0.7% estimated, according to stereology principles, by the ratio between the aggregated surface occupied in random sections by the plasmalemmal vesicles and the entire area of the endothelial cell profile (Williams 1977) measured in and computed for each of the 113 images considered. In diabetic rats these figures were on average 1.3- and 1.4-fold higher, respectively (Table 4).
Density of pre-embedding immunolabeling (gold particles/μm membrane length) for caveolin-1 in purified endothelial plasma membranes
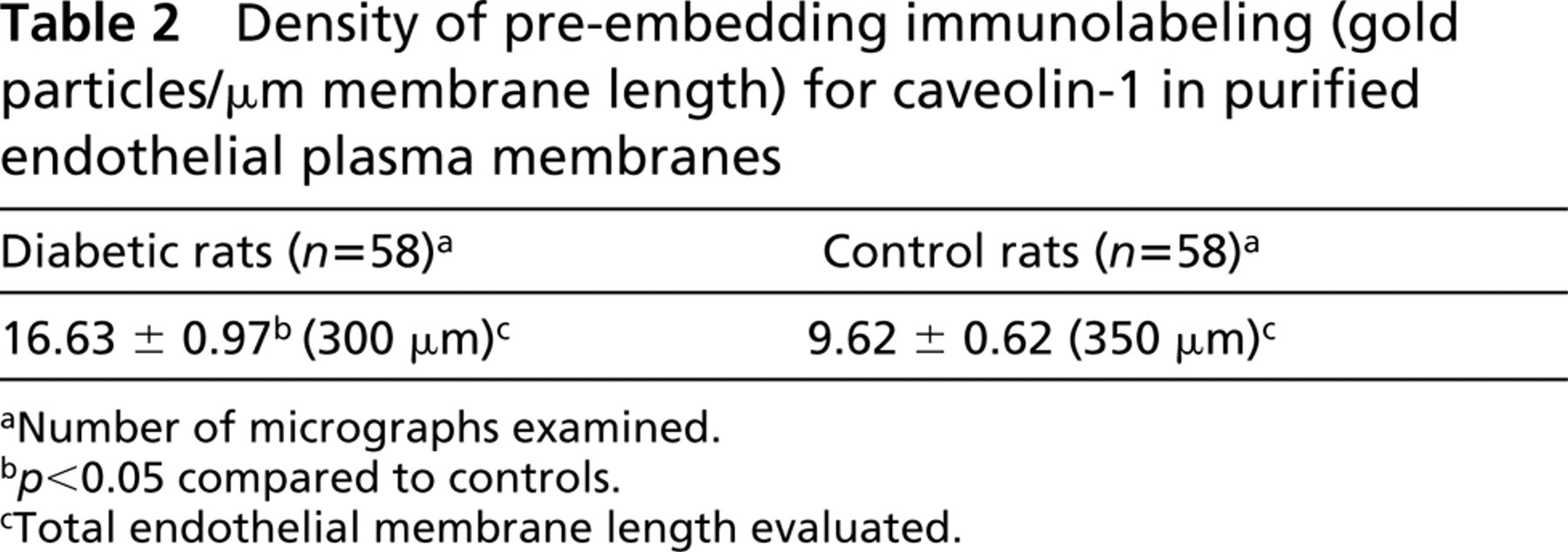
aNumber of micrographs examined.
b p<0.05 compared to controls.
cTotal endothelial membrane length evaluated.
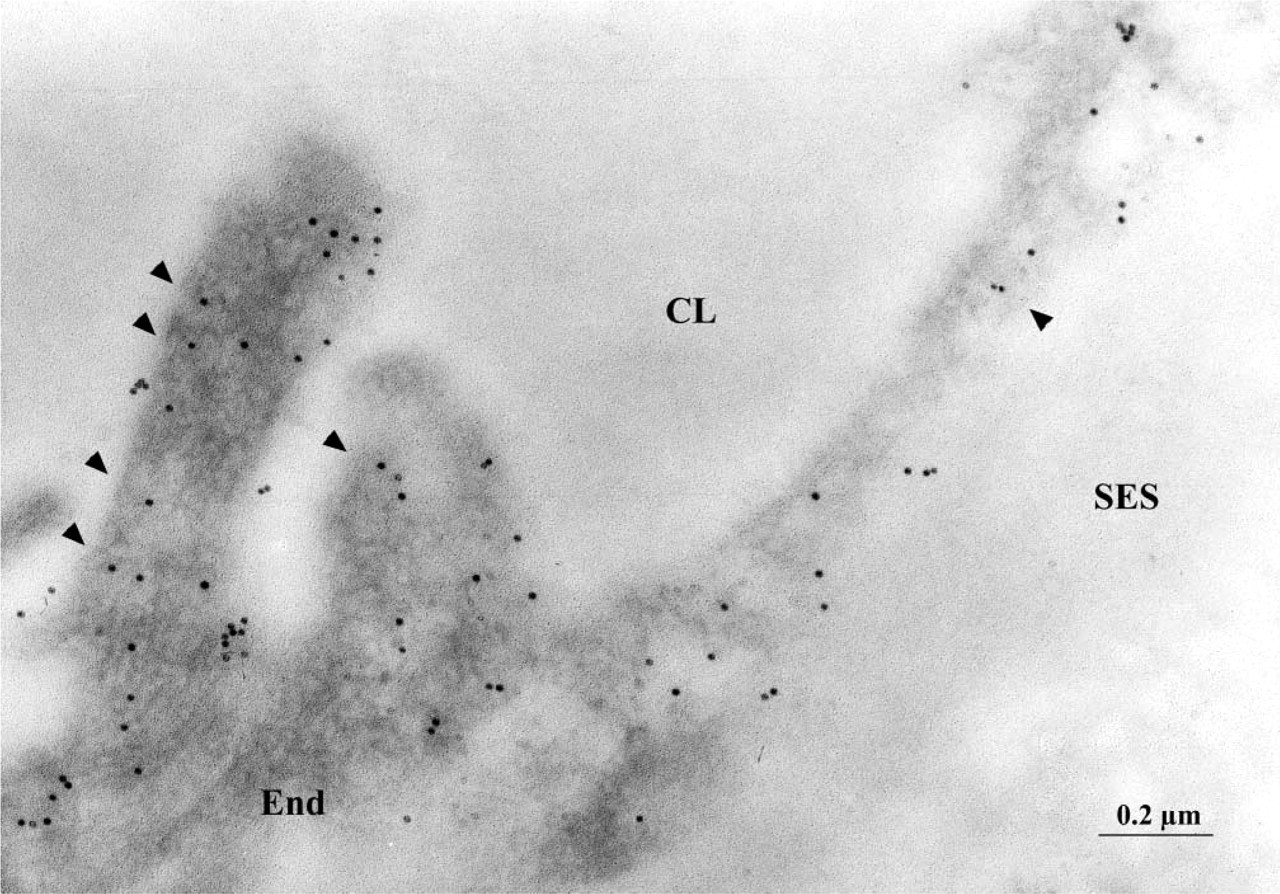
Postembedding ICC labeling for caveolin 1 over the endothelial cell profile in the lung of a diabetic animal. Because of the inherently low contrast of membranes, only some of the plasmalemmal vesicles are visible (arrowheads). CL, capillary lumen; End, endothelium; SES, subendothelial space. Original magnification ×73,600.
Whether or not this diabetes-induced increase in plasmalemmal vesicles number and aggregated volume correlates with meaningful variations in the transendothelial transport of macromolecules was tested by monitoring (a) the steady-state transendothelial distribution of the endogenous albumin and (b) the egress of an exogenous tracer, DNP-BSA, from the circulation under conditions of non-equilibrium. The tracer was injected into the bloodstream of the animals to preserve as much as possible the physiological conditions of the transendothelial transport. Its detection was performed with anti-DNP antibodies that neatly differentiate the haptenated albumin from the endogenous plasma pool (Figure 5). Both endogenous and exogenous albumins were detected in the capillary lumen and in the interstitial space. Over the endothelial and epithelial cell profiles, most of the gold particles were associated with the plasmalemmal vesicles. No open interendothelial junctions permeated by the tracer were detected throughout the examined areas.
Morphometrical evaluation (gold particles/μm2) postembedding immunolabeling for caveolin-1 over endothelial cell profiles
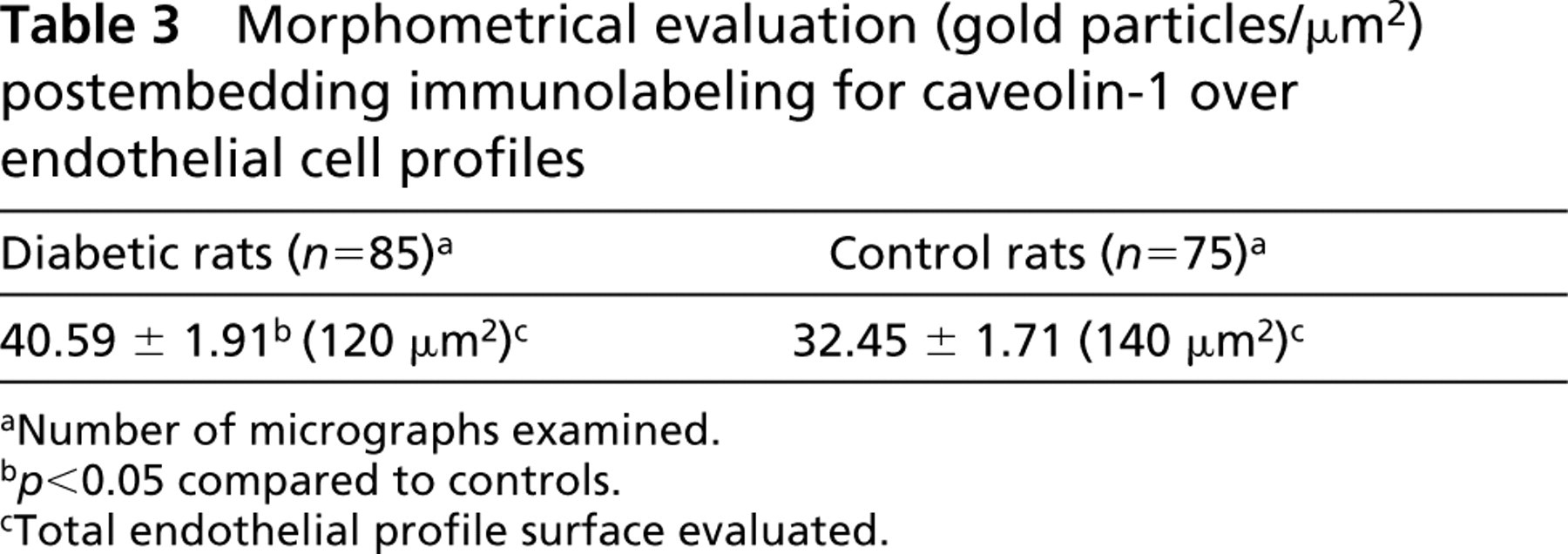
aNumber of micrographs examined.
b p<0.05 compared to controls.
cTotal endothelial profile surface evaluated.
Morphometrical evaluation of numerical density (number of vesicle profiles/μm2 of endothelial profile) and fractional volume (percent of endothelial volume) occupied by plasmalemmal vesicles (PV) in lung alveolar endothelium
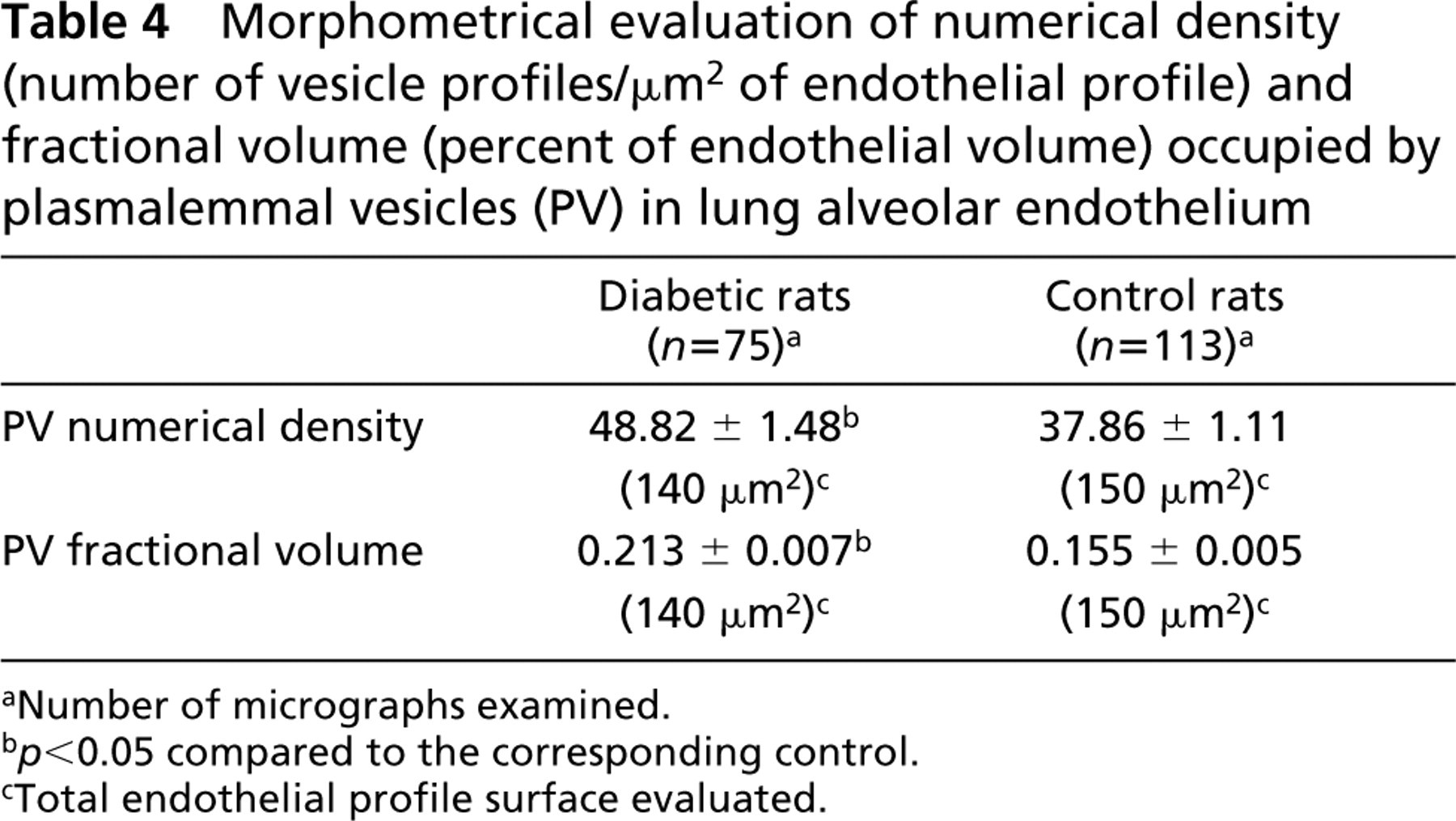
aNumber of micrographs examined.
b p < 0.05 compared to the corresponding control.
cTotal endothelial profile surface evaluated.
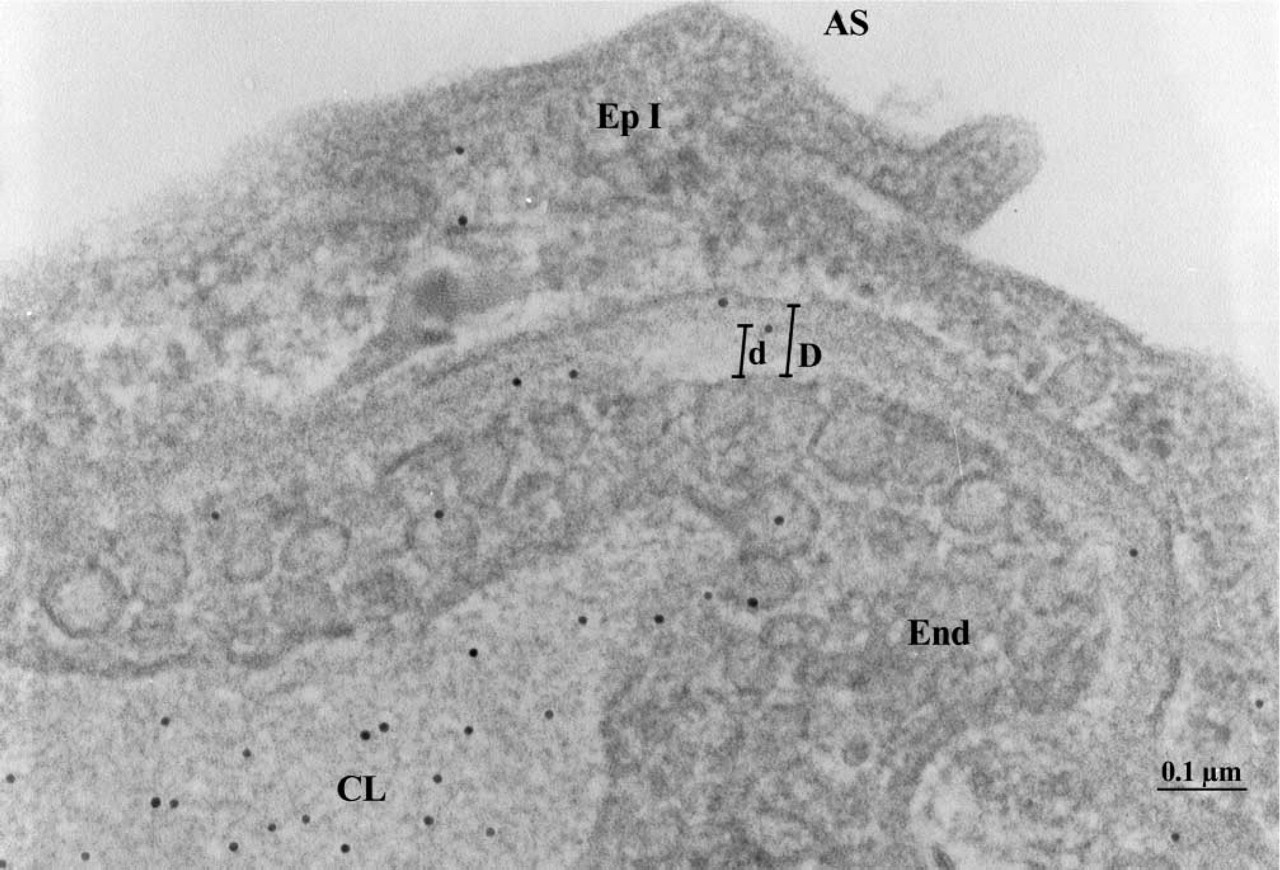
ICC detection of DPN-BSA in the alveolar wall of a diabetic rat. AS, air space; CL, capillary lumen; End, endothelium; EpI, epithelial cell type I; D, aggregated thickness of the endothelial and epithelial basement membrane; d, position of the DNP-BSA molecule with respect to the abluminal endothelial plasmalemma. Original magnification ×122,500.
The quantitative morphometric evaluation of the labeling for endogenous albumin (Table 5) shows that approximately 1.4-fold more gold particles were detected per unit surface of endothelial cells in diabetic animals than in the control. Similarly, the density of labeling over the endothelial basement membrane was 1.6-fold superior in diabetic animals. The same tendency for increased endothelial uptake of macromolecules from the pulmonary circulation of diabetic rats was registered when the transendothelial transport was monitored before the tracer reached a steady-state distribution between lumen and interstitium (Table 6). Identical densities of labeling for DNP-BSA were found in the capillary lumen of both diabetic and normal animals. However, the presence of the tracer over the endothelial cell profiles and in the space between endothelium and alveolar epithelium was higher in diabetes, 1.5-fold and 2.1-fold, respectively.
Intensity of the labeling for endogenous albumin (gold particles/μm2) over endothelium and its basement membrane in lung capillaries
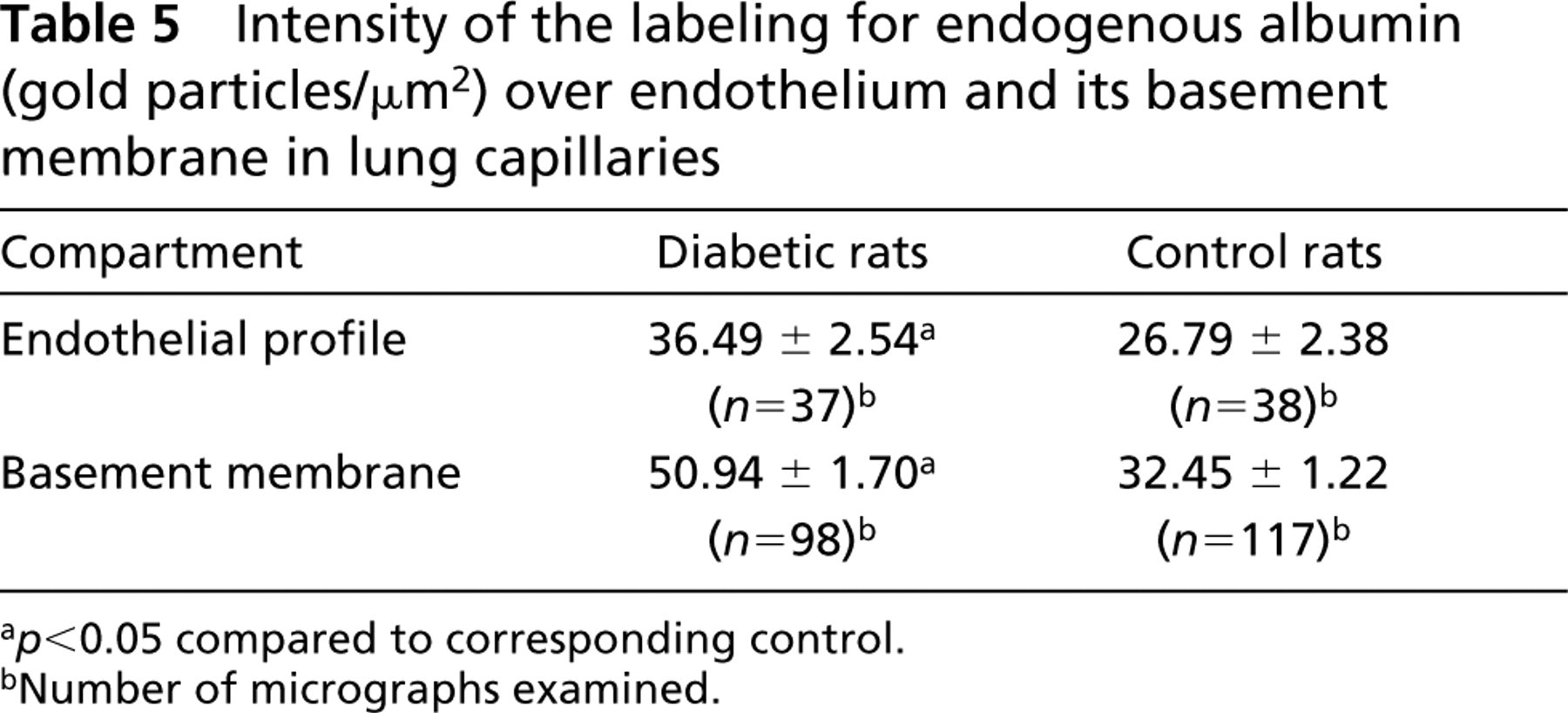
a p<0.05 compared to corresponding control.
bNumber of micrographs examined.
Density of labeling (gold particles/μm2) for DNP-BSA in lung capillaries at 15 min after IV injection of tracer
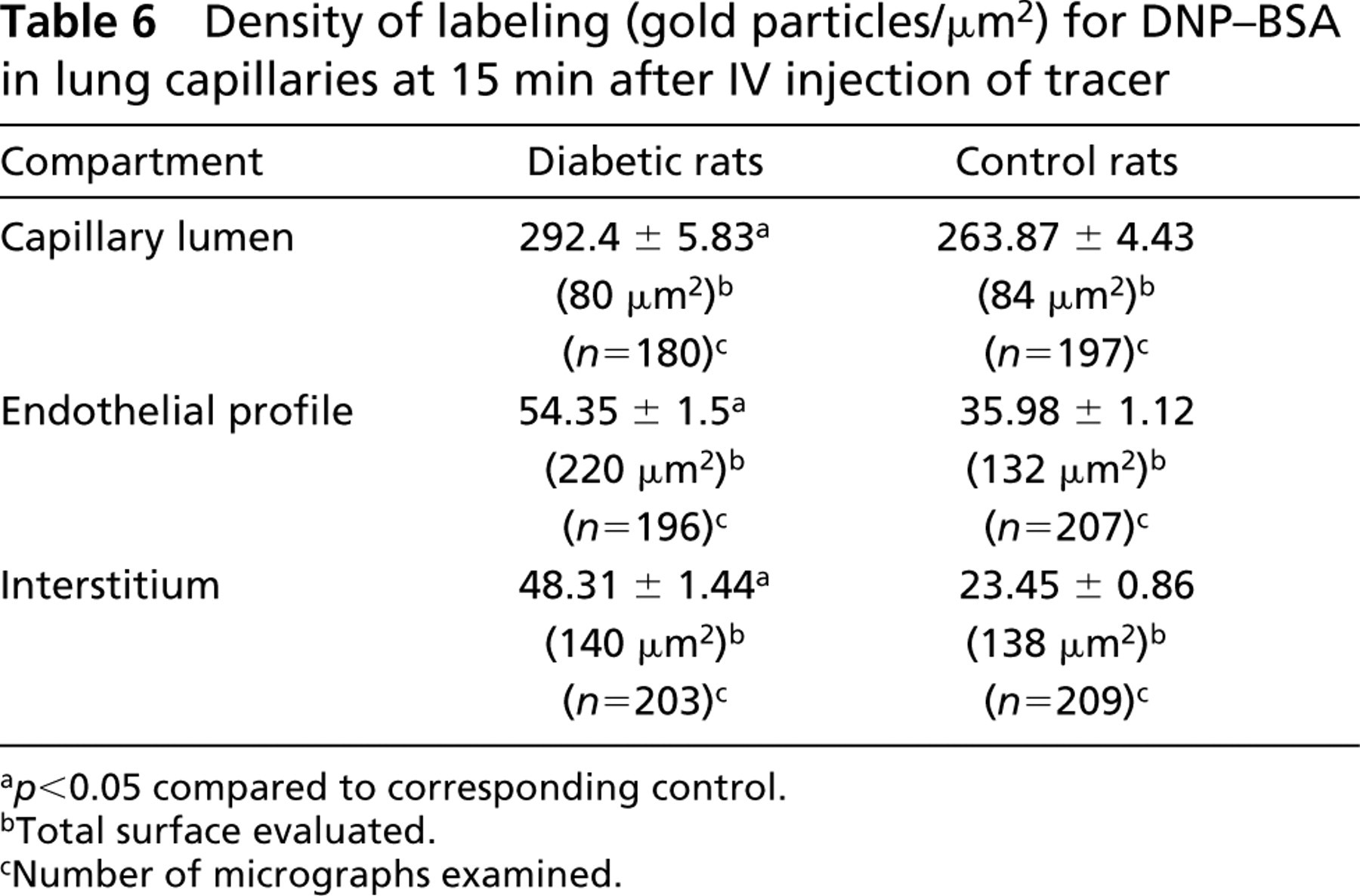
a p<0.05 compared to corresponding control.
bTotal surface evaluated.
cNumber of micrographs examined.
Because it is impossible to distinguish a borderline between the basement membranes of endothelial and type I pneumocytes in the alveolar wall, we measured the aggregated thickness of the two. We found that in normal animals this was 70 ± 0.7 nm and increased to 90 ± 0.8 nm (mean ± SEM) for the diabetic lot. The Wilcoxon and the Mann and Whitney tests indicated a confidence level of 95% for the assertion that the two distributions were different. These results show that, at 3 months after the onset of diabetes, the thickness of the basement membrane also begins to increase in the pulmonary vascular bed. However, its porosity does not appear to change. This was tested by measuring, across the basement membrane, the position of each gold particle indicating the presence of DNP-BSA molecules transported to the abluminal side of the capillary wall. We found that the spatial distribution of the DNP-BSA detected between the endothelium and the alveolar epithelium is identical for both normal and diabetic animals. The mean value of the ratio between the distance separating each gold particle from the abluminal front of the endothelium and the aggregated thickness of the endothelial and epithelial basement membranes at that place was 0.47 ± 0.007 (n = 861) and 0.48 ± 0.006 (n = 1530) for the normal and diabetic lots, respectively.
Discussion
Contrary to the situation in organs such as retina, kidney, limbs, and nerves, in which diabetic microangiopathy leads with a higher incidence to organ failure, the lung microcirculation of diabetic patients does not appear to be affected by clinically dramatic consequences. Only apparently superficial effects of the disease on this organ, such as retardation of lung development in the fetuses of diabetic mothers, alterations of several pulmonary physiological parameters, and increased susceptibility to infections, have been described. However, the present study shows that the pulmonary microvasculature is not spared by diabetes. We have observed systematic alterations in the architecture of the diabetic rat lung, particularly manifested as capillary and alveolar luminal narrowing and collapse, and an increase in extracellular matrix volume. Similar modifications, accompanied by a disorganization of the endothelial surface charges concomitant with an increase in the intravascular presence of macrophages, have been previously reported as occurring in the lungs of hamsters at 6 months after STZ induction of diabetes (Popov and Simionescu 1997). The existence of such alterations at the morphological level makes plausible the hypothesis that the physiology of the pulmonary microcirculation is also impaired in diabetic animals. Because the transendothelial macromolecular transport (transcytosis) represents one of the essential functions of the capillary endothelium, we have focused our study on this aspect. We have chosen to monitor the transendothelial distribution and egress of the most abundant plasma protein, albumin, by using quantitative ICC.
The morphometrical evaluation of the presence of endogenous albumin in the endothelial cells and in their basement membrane suggests that, at equilibrium, approximately 1.6-fold more albumin is found in the abluminal compartment of the alveolar capillaries of diabetic rats. This appears to represent not an increase in the “available space” for this protein in the alveolar wall interstitium but rather an active, intensified transcytosis, as indicated by the fact that, in the diabetic lot, the endothelial cells themselves appear to take up on average 1.4-fold more albumin molecules than in the control animals. This scenario is further strengthened by the data obtained with the exogenous, hapten-tagged variant of serum albumin (DNP-BSA) injected into the circulation. We have previously demonstrated (Ghitescu and Bendayan 1992) that such an approach preserves the normal physiological conditions in the vascular system and that the DNP-BSA is handled by the endothelium in exactly the same way as its endogenous untagged counterpart. Only 15 min after administration of the tracer, the density of labeling for DNP-BSA over the endothelial profiles amounted to 13.5% of that over the pulmonary capillary lumen in normoglycemic rats and 18.6% in diabetic animals (Table 6). The amount of DNP-BSA detected in the space between the alveolar endothelial and epithelial cells also increased rapidly at levels approximately twofold in diabetic than in normal rats. These results prove that, during the early stages of experimental diabetes, the lung capillaries, like other vascular beds endowed with a continuous type of endothelium, become hyperpermeable to plasma macromolecules.
The signal for endogenous albumin or for DNP-BSA, localized over the endothelial cell profile, was almost exclusively associated with the plasmalemmal vesicles, either open to one of the cell fronts or apparently closed, within the cytoplasm. We interpret this as a proof that the transendothelial transport of albumin takes place via the caveolar system and that, as a corollary, the diabetes-induced increase in permeability is due either to hypertrophy of this system or to an acceleration in the vesicular dynamics. As we will further discuss, our results support the first case. The involvement of caveolae in the transcytosis of macromolecules was recently questioned by the model of caveolin knockout mouse in which, despite the capillary endothelium lacking caveolin and caveolae, normal (Drab et al. 2001) or even higher concentrations of albumin were found in the interstitial fluid (Schubert et al. 2002). This challenge was only apparent, however, because the caveolin-null animals reportedly show caveolin-independent vesicles in the venular part of the microvasculature (Drab et al. 2001) and significant alterations at the level of the interendothelial junctions (Schubert et al. 2002).
The interendothelial clefts represent an alternative pathway, frequently considered, for the egress of molecules from the circulation. This is particularly attractive in relation to the so-called hemodynamic hypothesis, which regards the documented increase in capillary pressure as one of the main causes of diabetic microangiopathy (Tooke 1996). Videodensitometry has shown that capillary leakage of Na-fluorescein in the skin is significantly higher in long-term diabetic patients than in healthy ones (Oomen et al. 1999). The small (376 Da) negatively charged dye is likely to evaluate the patency of the nonspecific passageways for water and small hydrophilic molecules (small endothelial pores in the terminology of vascular physiologists), most probably the interendothelial junctions. Incubation of endothelial cells in vitro with advanced glycation end products (AGE)-modified BSA or vascular endothelial growth factor (VEGF), conditions presumed to mimic those of diabetic patient blood vessels, was found to trigger a reduction in the number of molecules responsible for the adherens and tight junctions such as VE-cadherin, its associated β- and γ-cathenins (Otero et al. 2001) and occludin, respectively (Antonnetti et al. 1998). However, we did not observe in the lung alveolar capillaries of hyperglycemic or normal rats any interendothelial junctions permeated by either endogenous albumin or DNP-BSA. This result agrees with previous studies also reporting the absence of open intercellular clefts in various vascular beds of diabetic animals (McCuskey and McCuskey 1984; Caldwell and Slapnick 1992; Yamaji et al. 1993; Vinores et al. 1998), except when diabetes was combined for the long term with aggravating conditions such as hyperlipidemia (Popov et al. 1996). Taken together, these facts rule out the possibility of explaining the diabetes-induced hyperpermeability to macromolecules through more permissive interendothelial clefts. The latter appear to account only for the abnormal transendothelial passage of small-sized molecules (Bendayan 2002; Simionescu et al. 2002).
Our data show that diabetes produces in its early stage a net expansion of the compartment of plasma-lemmal vesicles, which correlates with the documented hyperpermeability of lung alveolar endothelium to albumin. Immunoblotting assays and quantitative ICC performed on purified plasma membrane preparations have shown that, in diabetic animals, the luminal surface of the alveolar endothelium contains a significantly higher amount of caveolin 1 but not of other plasmalemma-asociated proteins, such as actin and annexin II. The existence of a direct relationship between the expression of caveolin and the number of caveolae has been previously demonstrated (Scherer et al. 1994) and confirmed by our results. Therefore, the natural conclusion is that, in diabetes, more caveolae come into direct contact and fuse with the apical plasmalemma of the pulmonary endothelium.
We are not considering at present what proportion of these organelles are connected to the plasma membrane and how many might be free in the cytoplasm as free-floating plasmalemmal vesicles. This was the reason for performing in parallel the ICC detection and quantification of caveolin in the purified plasma membrane preparations as well as in the whole endothelial cell profile fixed in situ. Both assays indicated a significant (1.73-fold and 1.24-fold, respectively) increase in the amount of caveolin 1 in diabetes.
The comparative morphometrical analysis of the caveolar compartment in the alveolar endothelium of diabetic and normoglycemic rats has shown that the overexpression of caveolin in diabetes is accompanied by a notable increase in the number of plasmalemmal vesicles and in the partial volume occupied by them in the endothelial cell. Consequently, the data presented here define these vesicles as the main endothelial structural elements involved in transendothelial transport of plasma proteins and the ones responsible for the diabetes-induced increase in endothelial permeability. It is likely that this mechanism is not limited to the alveolar capillaries but may also account for the hyperpermeability of other microvascular beds endowed with a continuous type of endothelium, such as retina and nerves, in which diabetic microangiopathy is significantly more spectacular.
An attractive hypothesis regarding the mechanism by which diabetes induces caveolin overexpression involves the altered VEGF production in the tissue surrounding the blood vessels. VEGF is a family of potent mitogens with relative specificity for endothelial cells, playing a key role in angiogenesis and also able to increase vascular permeability, hence its alternative name of vascular permeability factor. The implication of VEGF in diabetic microangiopathy draws its support from studies that have already documented that vascular complications, particularly those in the retina and kidney, are associated with transcriptional activation of the VEGF gene (Hammes et al. 1998; Cooper et al. 1999). An upsurge in the amount of VEGF (more than fourfold) was also demonstrated in the lung of spontaneously diabetic BB rats at the onset of the disease. The increase is identical to that measured in the kidneys of the same animals (Braun et al. 2001). Possible VEGF producers in the lung are the vascular and airway smooth muscle cells (Mata-Greenwood et al. 2003) and the endothelial cells themselves (Favre et al. in press). VEGF gene transcription is stimulated by a host of factors such as hypoxia, other growth factors, and cytokines, and also by several particular conditions of diabetes, such as hyperglycemia (Williams et al. 1997), advanced glycation end products (AGEs) (Treins et al. 2001; Yamagishi et al. 2002), and oxidative stress (Beinert et al. 2000; Sasaki et al. 2000). Moreover, when various types of endothelial cells were treated in vitro with VEGF, a rapid increase in the number and diameter of caveolae was evident (Vasile et al. 1999; Chen et al. 2002). However, even if proved true, this scenario of VEGF-dependent microvascular hyperpermeability in diabetes must actually be considerably more complex. Several experimental data do not entirely fit to it. For example, VEGF was proved to induce, over the long term, a change in endothelial cell phenotype from continuous to fenestrated (Roberts and Palade 1995; Chen et al. 2002). None of these fenestrations was observed in the microvasculature of our diabetic rats. In addition, the oxidative stress present in diabetes and known to activate the production of VEGF (Chua et al. 1998) was reported to inhibit caveolin palmitoylation and therefore its association with the membrane (Parat et al. 2002).
Caveolin overexpression by endothelial cells in diabetes is certainly not a singular example. Aside from the very well-demonstrated increase in the production and secretion of extracellular matrix components, several other proteins, such as endothelin and vascular cell adhesion molecule-1, key to other important endothelial functions such as the regulation of vascular tone and leukocyte diapedesis, respectively, are affected by diabetes (Ribau et al. 1999; Apostolova et al. 2001).
Our results also show that the basement membrane of alveolar endothelium, like that in other vascular beds, such as skeletal muscle (Ghitescu and Bendayan 1992) and mesentery (Arshi et al. 2000), does not represent a barrier to the blood-tissue transport of macromolecules, at least of the serum albumin size. A restricted diffusion of the endogenous albumin or DNP-BSA across the thickness of the basement membrane should manifest itself through an accumulation of the endogenous albumin or DNP-BSA in the lamina rara, the region of the basement membrane adjacent to the endothelium (Bendayan et al. 1986). No such distribution was observed. In diabetic rats the alveolar epithelial and endothelial basement membranes thicken to an extent (1.3-fold) comparable to that detected in the kidney glomerulus at the same early stage of the disease (Weynand et al. 1999; Doucet et al. 2000). However, unlike the glomerular basement membrane, which becomes leaky in diabetes, the porosity of the alveolar basement membrane does not change. The same relative distribution of the albumin molecules across the thickness of this structure is encountered in normal and diabetic rats.
Footnotes
Acknowledgements
Supported by grants from the Juvenile Diabetes Research Foundation International (no.1-2001-551) and the Canadian Institute of Health Research (no. MOP-9702).
This article represents part of the work required for fulfillment of the MSc program by M. Pascariu.
We thank Dr Irene Londoño, Ms Diane Gingras, and Ms Elisabeth Gervais for their precious help.