
Abstract
Adequate contrast of ultrathin cryosections is crucial for evaluating morphological detail to assess immunocytochemical localization at the electron microscopic level. We have developed a positive staining method for achieving contrast in ultrathin cryosections, from tissue fixed only in paraformaldehyde, that provides excellent contrast at the electron microscopic level.
U
Although many factors contribute to the appearance of ultrathin cryosections, one important factor relates to the fixation conditions. Typically, cells or tissues have been fixed in paraformaldehyde (PFA) (2–4%) and glutaraldehyde (GLA) (0.1–0.2%) in a buffered solution to obtain good morphological detail in immunocytochemical experiments. However, in some situations the antigen-antibody interaction may be inhibited by the inclusion of small amounts of GLA in the fixation solution (see below). In situations where small amounts of GLA in the fixative were not feasible, achievement of adequate morphological detail in ultrathin cryosections may be more difficult. Here we describe a method to provide positive contrast to ultrathin cryosections, resulting in excellent ultrastructural appearance in tissue that was fixed only in PFA. Furthermore, this method is compatible with the use of colloidal gold immunoprobes.
Materials and Methods
Reagents
Alexa Fluor 488 and 594 goat anti-chicken IgG and Pro-Long antifade kit were obtained from Molecular Probes (Eugene, OR). Mouse anti-p230 trans-Golgi monoclonal antibody was from BD Transduction Laboratories (San Diego, CA). Affinity-purified goat anti-chicken IgY 4-nm colloidal gold particles and affinity-purified, biotinylated donkey anti-mouse F(ab′)2, horseradish peroxidase (HRP)-conjugated streptavidin were purchased from Jackson ImmunoResearch (West Grove, PA). Goat anti-HRP 5-nm colloidal gold particles were from British Biocell International (Cardiff, UK). Fetal bovine serum was obtained from Invitrogen (Carlsbad, CA). All immunological reagents were handled in accordance with the manufacturers’ recommendations and used within the expiration date for each product. In addition, an anti-peptide antibody specific for caveolin-1α (CAV-1α) was generated in chickens. The immunogen was amino acids 3–14 from the N-terminal region of the CAV-1α molecule with the amino acid sequence GGKYVDSEGHLY. Immunocytochemical characterization of this antibody has been reported (Robinson and Vandré 2001; Lyden et al. 2002).
Processing of Ultrathin Cryosection for Immunocytochemistry
Human full-term placenta was used as the model tissue in these studies. Tissue was obtained from the Ohio State University tissue procurement service according to a protocol approved by the Ohio State University Human Subjects Institutional Review Board. Tissue samples from Cesarean deliveries were processed for fixation as rapidly as possible (typically within 20 min). Four different placentas were used in this study.
Tissue was cut into small pieces and put into fixative. Three aldehyde fixation conditions were employed: (a) 4% PFA in 0.1 M sodium cacodylate buffer, pH 7.4, containing 5% sucrose; (b) 4% PFA plus 0.1% GLA in the same buffer; and (c) 4% PFA plus 0.2% GLA in the same buffer. Tissue samples were fixed for 2 hr at room temperature (RT). The samples were washed and then embedded in 10% gelatin in the same buffer. The solidified gelatin was cut into small pieces and then infiltrated with 2.3 M sucrose in 0.1 M sodium cacodylate (pH 7.4) overnight at 4C. After sucrose infiltration, the samples were mounted on specimen pins designed to fit the Reichert cryoultramicrotome, frozen in liquid nitrogen, and stored in liquid nitrogen until they were sectioned.
Ultrathin cryosections were cut with a Reichert Ultracut E equipped with an FC 4D cryounit. Sections (100-nm thickness or less) were cut on a Cryo P diamond knife (Diatome-US; Fort Washington, PA) and were collected on droplets of 0.75% gelatin–2.0 M sucrose (Takizawa and Robinson 1994) or 1% methylcellulose–1.15 M sucrose (Liou et al. 1996), and then transferred to 0.25% poly-
For immunofluorescence microscopy, ultrathin cryosections mounted on cover slips were immersed in PBS for 15 min at 37C, washed three times in PBS, and incubated in a solution containing 5% fetal bovine serum in PBS (FBS-PBS) to block nonspecific protein-binding sites; 0.05% sodium azide was also present. The cryosections were then incubated with chicken anti-human caveolin-1α (diluted 1:500) in FBS-PBS for 30 min at 37C. The cryosections were rinsed in PBS four times over 15 min, immersed in FBS-PBS, and then incubated with Alexa Fluor 488-labeled goat anti-chicken IgG (diluted 1:200) in FBS-PBS for 30 min at 37C. After immunostaining, the immunofluorescence preparations were washed extensively in PBS and then mounted on glass microscope slides in ProLong antifade medium. Control sections received the same treatment except for omission of the primary antibody or substitution of pre-immune chicken antibody for the primary antibody.
For electron microscopy, ultrathin cryosections mounted on EM grids were incubated with anti-CAV-1α (diluted 1:50) following the same schedule as used for the immunofluorescence procedure. Detection of the primary antibody binding sites was then achieved with a 30-min incubation in FBS-PBS containing goat anti-chicken 4-nm colloidal gold particles (diluted 1:10) at 37C. Other sections were incubated with anti-p230, a trans-Golgi marker (25 μg/ml), incubated with donkey biotinylated anti-mouse F(ab)'2 (15 μg/ml), and then incubated with HRP-streptavidin (10 μg/ml). Each of these steps was for 30 min at 37C with PBS washes in between. Detection of the binding sites of p230 trans-Golgi was achieved with a 30-min incubation in FBS-PBS containing goat anti-HRP 5-nm colloidal gold particles (diluted 1:10) at 37C. The sections were washed five times in PBS to remove unbound colloidal gold. The sections were then fixed with 2% glutaraldehyde-PBS for 15 min at RT and then washed three times in distilled water.
Visualization and preservation of ultrastructure of cryosections was achieved in four different ways. (a) Ultrathin cryosections from tissue fixed in 4% PFA and 0.1% GLA were subjected to contrast enhancement with 0.3% uranyl acetate in 1.85% methyl celluose (UA-MC) (Griffiths et al. 1984). (b) Ultrathin cryosections from tissue fixed in 4% PFA were also subjected to the UA-MC method. (c) Ultrathin cryosections from tissue fixed in 4% PFA were post-fixed in 1–2% osmium tetroxide in 0.1 M cacodylate buffer, pH 7.4, for 15 min at RT and then washed three times in distilled water. The cryosections were then processed with modifications of the polyvinyl alcohol (PVA)-embedding method (Tokuyasu 1989). The postfixed cryosections were incubated with 0.8% uranyl acetate–1.6% PVA (UA-PVA) for 15 min and then washed briefly in PVA. The cryosections were then stained with a mixture of 0.0015% lead citrate and 2% PVA (LC-PVA) for 15 min at RT (both UA-PVA and LC-PVA were passed through a filter with 0.2-μm pore size before use). The EM grid on a drop of LC-PVA was collected with a wire loop and the excess LC-PVA fluid was removed with a small piece of filter paper. The grid was then ready for examination. Alternatively, a small square of thin formvar film (slightly wider than the diameter of the EM grid) was floated on a drop of LC-PVA. Simultaneously, an EM grid containing cryosections was incubated in LC-PVA for 15 min. The formvar film was then picked up on an EM grid with a wire loop and the excess LC-PVA fluid between the cryosection and the overlying formvar film was removed with filter paper. (d) Ultrathin cryosections from tissue fixed in 4% PFA were postfixed in reduced osmium (2% osmium tetroxide–1.6% potassium ferrocyanide in 0.1 M cacodylate buffer, pH 7.4) for 15 minutes. The cryosections were then contrast-enhanced with the same procedure used in method c above. Cryosections were observed with a Philips CM-12 electron microscope operated at 60 kV.
Processing of Cultured Cells for Immunocytochemistry
Cultured human umbilical vein endothelial cells (HUVECs) were also used to compare primary fixation conditions. HUVECs were obtained from Clonetics (Clonetic/Cambrex; Walkersville, MD) and grown in endothelial growth medium (Clonetics). Cells were cultured on 12-mm glass coverslips in 24-well culture dishes. Cultured cells were fixed under the same three conditions used for placental tissue. Fixed cells were washed in PBS at least six times. Cells were then permeabilized with 0.2% Triton X-100 in PBS for 5 min at RT. Cells were washed three times in PBS and subsequently incubated in FBS-PBS for 1 hr at RT to block nonspecific protein binding sites. Cells were then incubated with the same concentration of anti-CAV-1α and under the same conditions as used for the ultrathin cryosections. Similarly, secondary antibodies were used under identical conditions as employed with the ultrathin cryosections (in HUVECs some coverslips were incubated with Alexa 488-labeled goat anti-chicken IgG and others were incubated with Alexa 594-labeled goat anti-chicken IgG).
Image Analysis
The fluorescence signals from immunolabeled ultrathin cryosections of human placenta and HUVECs were collected with a Nikon Optiphot microscope equipped with a Photometrics Cool Snap fx digital camera (Roper Scientific; Tucson, AZ). Fluorescence images were analyzed with the Metamorph image analysis software system (Universal Imaging; Downingtown, PA). The fluorescence images of the ultrathin cryosections from each fixation procedure were collected under identical conditions. Likewise, fluorescence images of HUVECs from each fixation procedure were collected under identical conditions. This permits comparisons of the fluorescence signals from each type of fixation. Differential interference contrast (DIC) images of sections and cells were also collected so that their morphology could be determined. Figures shown in this report were compiled in Photoshop 7.0 (Adobe Systems; Mountain View, CA).
Fluorescence intensity measurements of individual caveolae in HUVECs were obtained through Metamorph. A total of 100 individual caveolae were measured for each fixation condition (20 caveolae from five different cells per fixation condition in a single experiment). Cells were selected at random using DIC optics before collection of the fluorescence signal. In an attempt to select caveolae at random, a rectangle of uniform size was drawn in the selected cells with Metamorph. Five caveolae near each of the corners of the rectangle were selected for measurement of fluorescence. Fluorescence intensity measurements were made for each caveola using the line scan function of Metamorph. The background fluorescence adjacent to each caveola was subtracted from the peak fluorescence to obtain a fluorescence intensity measurement for each caveola. The mean fluorescence for each fixation condition ± SD was then plotted in a histogram.
Results
The fluorescence signal for immunolabeling of CAV-1α in ultrathin cryosections of terminal villi of human placenta was compared in tissue prepared with three different fixation conditions. A robust signal was obtained, even in ultrathin cryosections, when tissue was fixed in 4% PFA (Figure 1A). In terminal villi, the fluorescence signal was primarily in the endothelial cells, with little if any labeling of the syncytiotrophoblast. However, inclusion of GLA (0.1% or 0.2%) in the 4% PFA fixative essentially eliminated the detection of CAV-1α in this tissue (Figure 1B). Control experiments in which pre-immune antibody replaced the specific anti-CAV-1α antibody or in which primary antibody was omitted, but which did receive secondary antibody treatment, did not display fluorescence above background levels (not shown).
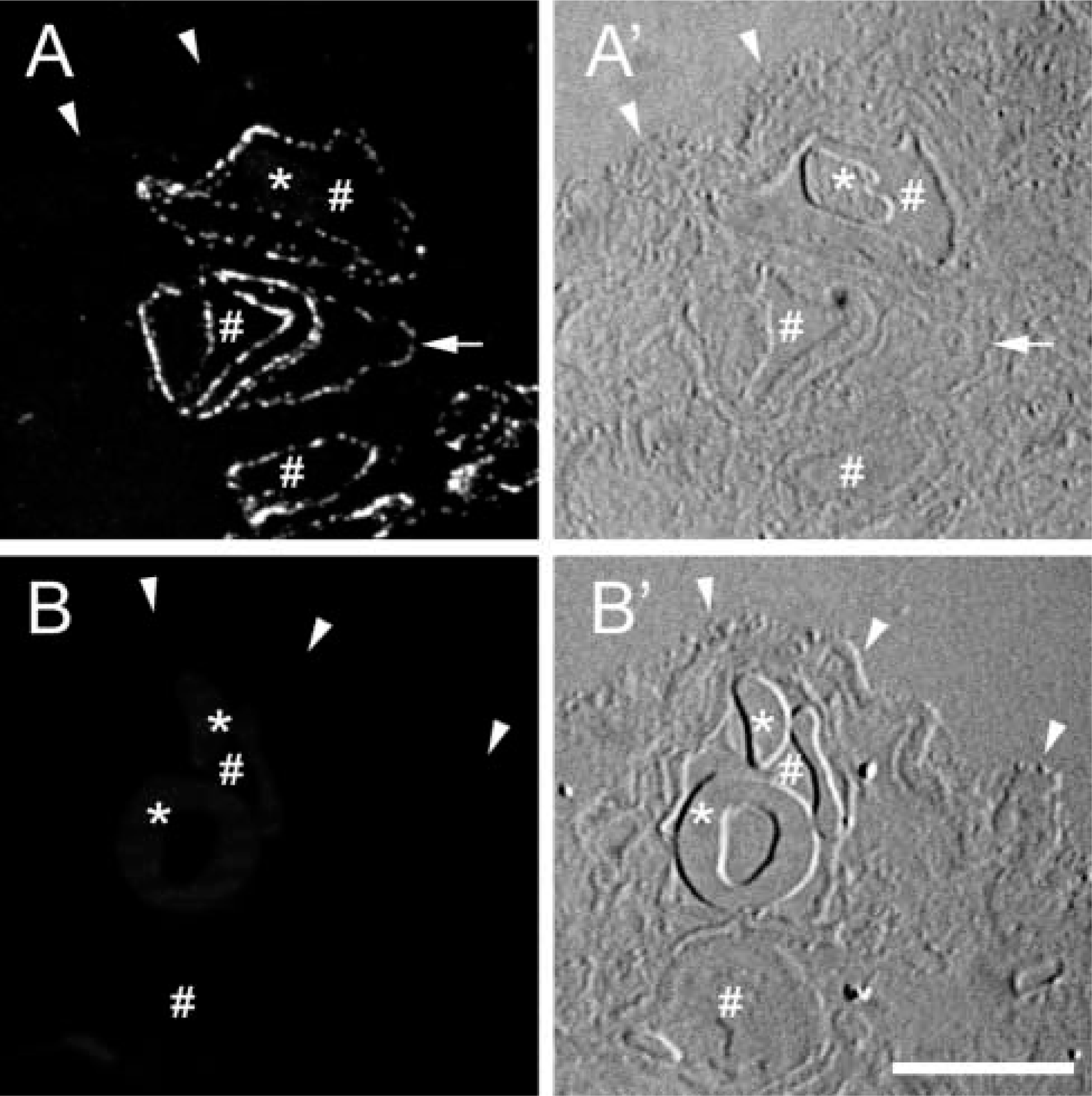
Comparison of the localization of CAV-1α in placenta fixed in 4% PFA or 4% PFA + 0.1% GLA. (
To determine whether GLA-induced inhibition of anti-CAV-1α binding was restricted to placenta, we expanded these studies to HUVECs, a caveolin-rich cell type. HUVECs were fixed and immunolabeled in the same manner as the placenta. Fluorescence images of cells fixed in the three different ways were collected in the same exact manner. Again, a robust signal was obtained from cells fixed in 4% PFA (Figure 2A). Cells fixed in 4% PFA plus 0.1% or 0.2% GLA had dramatically reduced labeling of caveolae (Figures 2B and 2C).
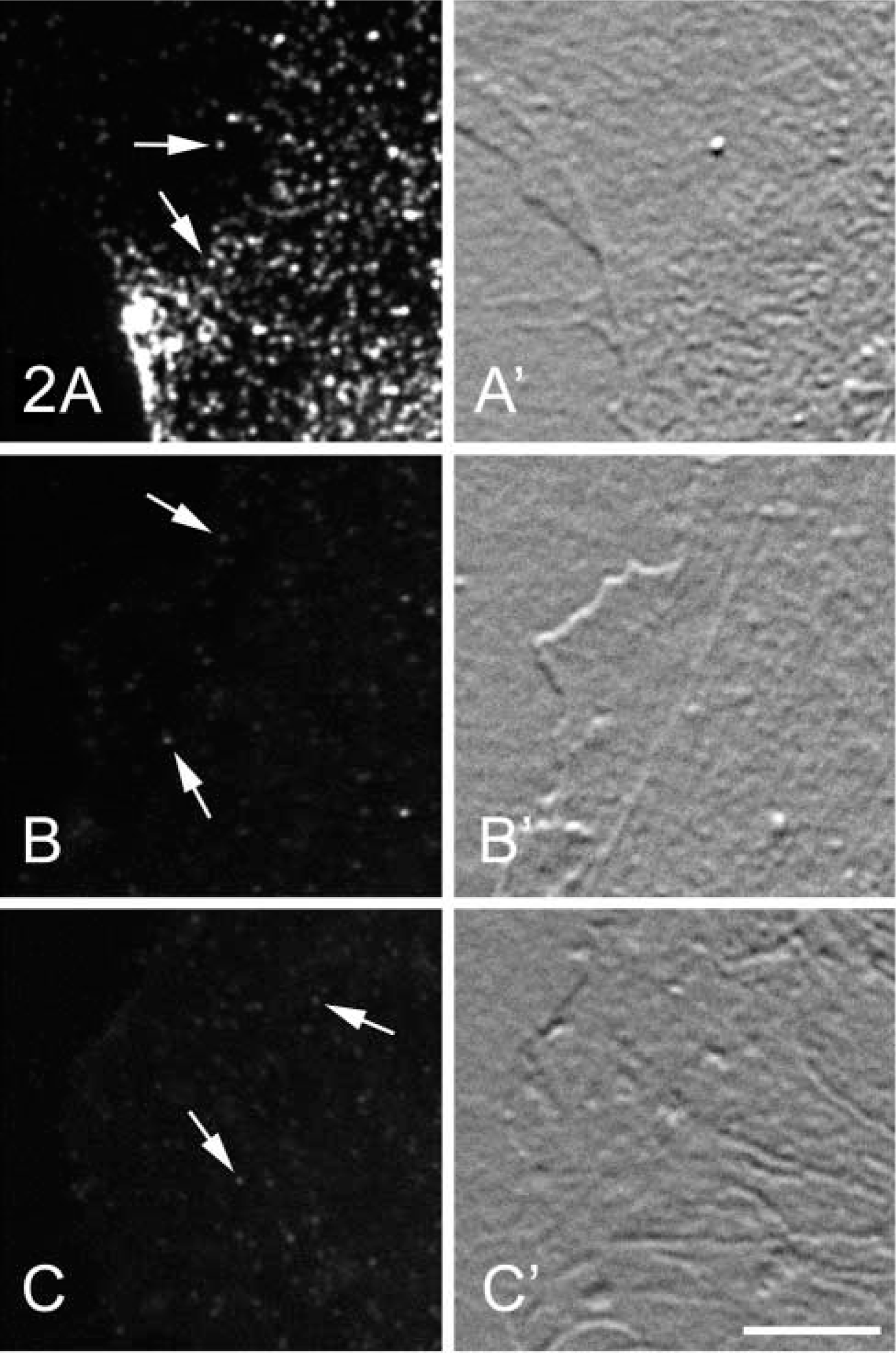
Comparison of the localization of CAV-1α in HUVECs fixed in 4% PFA or in 4% PFA + 0.1% or 0.2% GLA. (
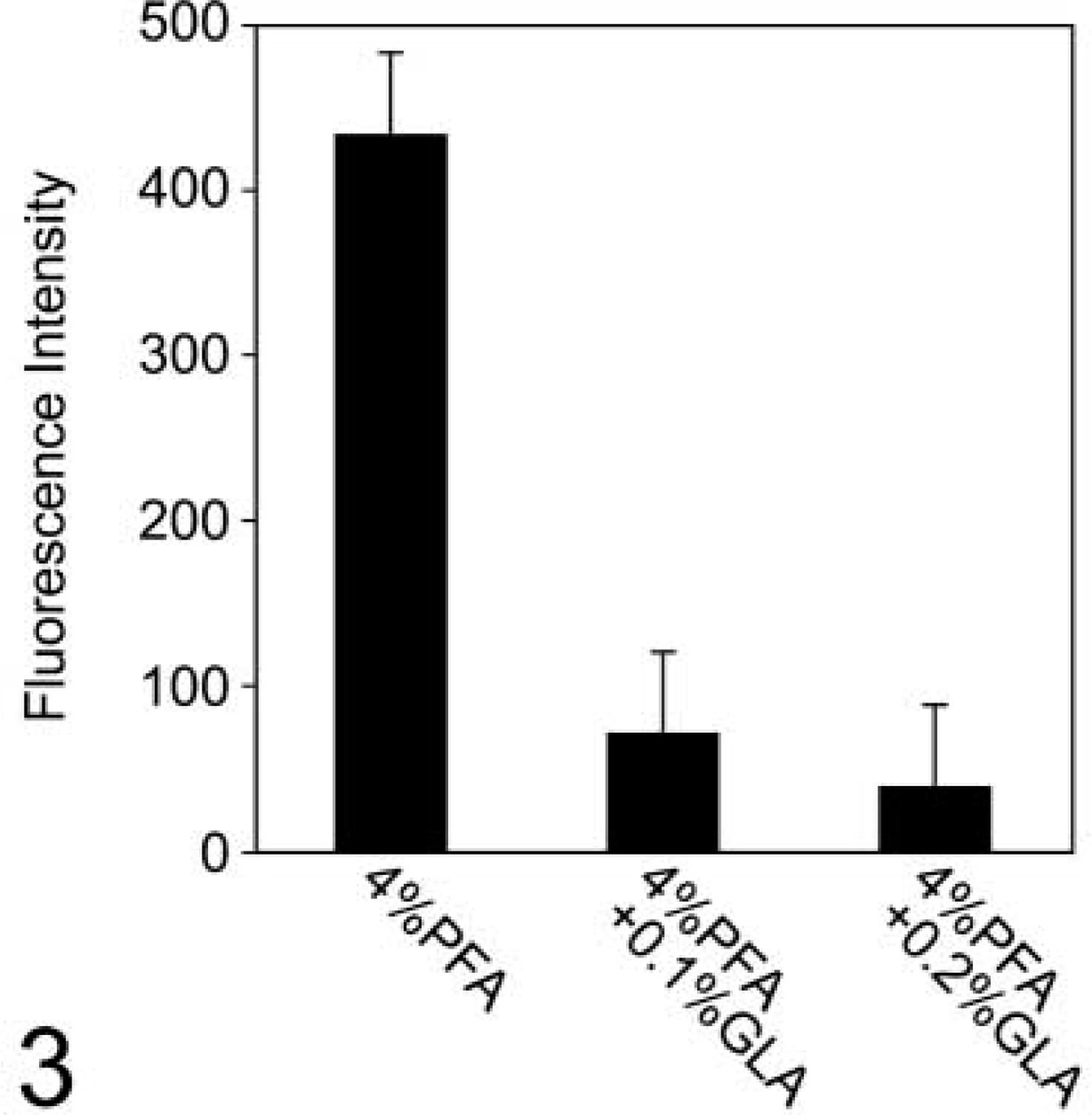
Comparison of fluorescence intensities of the CAV-1α immunofluorescence signals from caveolae in HUVECs fixed in 4% PFA or in HUVECs fixed in 4% PFA + 0.1% or 0.2% GLA. These quantitative data support the initial observation that inclusion of GLA in the PFA fixation inhibits the anti-CAV-1α - CAV-1α interaction (see Figure 2). Inclusion of 0.1% or 0.2% GLA inhibits the immunofluorescence signal by about 85% and 92%, respectively. The fluorescence intensities are the averages of 100 separate measurements ± SD for each fixation condition.
The fluorescence intensity of anti-CAV-1α immunofluorescence labeling in caveolae was measured in HUVECs fixed in the three different primary fixation protocols. Results from these fluorescence intensity measurements show that inclusion of 0.1% or 0.2% GLA in the fixative inhibits the immunofluorescence signal by about 85% and 92%, respectively (Figure 3). These values may actually be an underestimate of the inhibition induced by GLA on the immunofluorescence signal because some caveolae may be below the level of detection.
The ultrastructural appearance of ultrathin cryosections of placental tissue fixed in 4% PFA and then contrast-enhanced with the UA-MC method was of good quality and was comparable to that observed by other investigators using these preparative procedures (Figure 4). Post-fixation of ultrathin cryosections from tissue fixed in 4% PFA with ferrocyanide-reduced osmium and subsequent staining with uranyl acetate and lead citrate in PVA led to excellent fine structural appearance of these sections and provided a positive image (Figure 5). Postfixation of ultrathin cryosections from tissue fixed in 4% PFA with regular osmium tetroxide and then contrast-enhanced with uranyl acetate and lead citrate in PVA was not adequate for detailed analysis (data not shown). In addition, postfixation with ferrocyanide-reduced osmium and contrast enhancement with uranyl acetate but without lead citrate in PVA was also carried out. In this case, ultrastructural appearance was not satisfactory for proper fine structural analysis (data not shown).
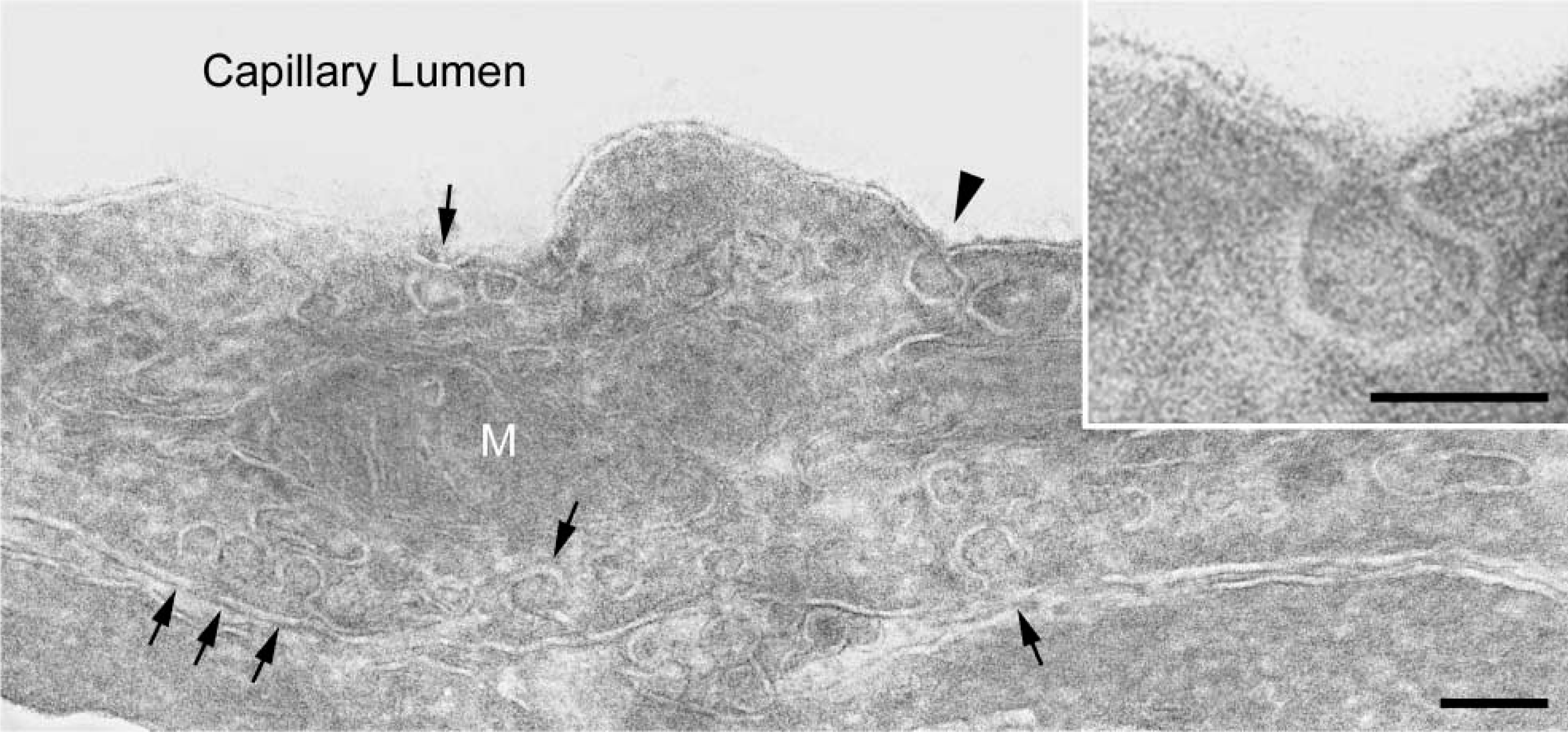
Ultrastructural localization of CAV-1α was achieved in endothelial cells from human placenta fixed in 4% PFA. Plasma membrane microdomains having the classical morphology of caveolae were decorated with 4-nm colloidal gold that was used as the secondary immunoprobe (Figure 6). Colloidal gold particles were restricted to caveola-like structures; little background labeling was detected in these preparations. There was essentially no colloidal gold labeling in ultrathin cryosections from placental tissue fixed in 4% PFA and 0.1% GLA (not shown).
We also examined the syncytiotrophoblast, a structurally complex cell type, to determine if good contrast with the new method could be achieved in an immunocytochemical preparation in this structure. We localized p230, a trans-Golgi network marker, in the syncytiotrophoblast. Immunogold particles (5 nm) indicating the distribution of p230 were restricted to vesicular and short tubular structures in the trans-Golgi network (Figure 7).
Uranyl acetate-methyl cellulose (UA-MC) method. An ultrathin cryosection of 4% PFA-fixed human placenta was negatively stained by the UA-MC method. Caveolae are well resolved in a placental endothelial cell (arrows). A mitochondrion (M) is evident. Bar = 100 nm. (
Discussion
Immunocytochemical localization of cellular antigens continues to be an important tool for experimental biologists. Here we describe an alternative to the UA-MC method for obtaining contrast and ultrastructural detail in ultrathin cryosections for immunoelectron microscopy. In our hands, this method is particularly useful for imaging samples that are fixed in PFA without any GLA, a fixation condition that may be a necessity because binding of some antibodies to their antigen is inhibited by the inclusion of small amounts of GLA in the fixation solution.
An antibody directed against the N-terminal region of CAV-1α readily detects this protein in caveolae in PFA-fixed placenta and HUVECs in immunocytochemical assays. However, inclusion of small amounts of GLA (0.1% or 0.2%) dramatically inhibits detection of CAV-1α in the same immunocytochemical procedures with this antibody (see Figures 1–3). It is known from the work of others that the interaction of some antibodies with their antigens is sensitive to the presence of GLA in the fixation solution. For example, Yokota and Okada (1997) used immunogold labeling of resin sections and particle counting methods to quantify the affect of GLA on the immunocytochemical detection of peroxisomal proteins. In that case, detection of acyl-CoA oxidase was inhibited by about 60% and detection of keto acyl-CoA thiolase was inhibited by about 70% by inclusion of 0.5% GLA in the fixative. Detection of catalase, on the other hand, was only slightly inhibited at that concentration of GLA. Those results show that even within the same organelle the binding of different antibodies to their antigens can display differential sensitivity to GLA fixation. An antigen-antibody interaction that we have studied in this report (i.e., anti-CAV-1α binding to CAV-1α) is particularly sensitive to glutaraldehyde fixation because virtually complete inhibition occurs at a GLA concentration of only 0.2%.
Typically, ultrathin cryosections are prepared from cells or tissues fixed in a mixture of PFA and GLA and then stained with UA-MC (e.g., Liou et al. 1996). We applied this method to placental tissue and observed good ultrastructural detail (data not shown). However, as noted above, this fixation method is not compatible with use of the antibody to CAV-1α that we have employed in this study, because GLA dramatically inhibits interaction of this antibody with its antigen. The UA-MC contrasting method was then applied to ultrathin cryosections of placenta fixed in 4% PFA. Good ultrastructural detail could be observed even in this case (Figure 4). However, we thought it desirable to obtain positive images of cell structure in ultrathin cryosections similar to those in standard epoxy resin sections. In an effort to achieve useful positive staining, we postfixed ultrathin cryosections with either osmium tetroxide or ferrocyanide-reduced osmium. Treatment of cryosections with regular osmium was not entirely satisfactory for critical examination of subcellular ultrastructure. However, use of ferrocyanide-reduced osmium, coupled with staining with uranyl acetate and lead citrate in polyvinyl alcohol, led to excellent fine structural detail in ultrathin cryosections from placental tissue fixed in only 4% PFA. The quality of cryosections prepared in this manner was at least equal to those prepared by the conventional method (i.e., contrast enhancement using UA-MC).
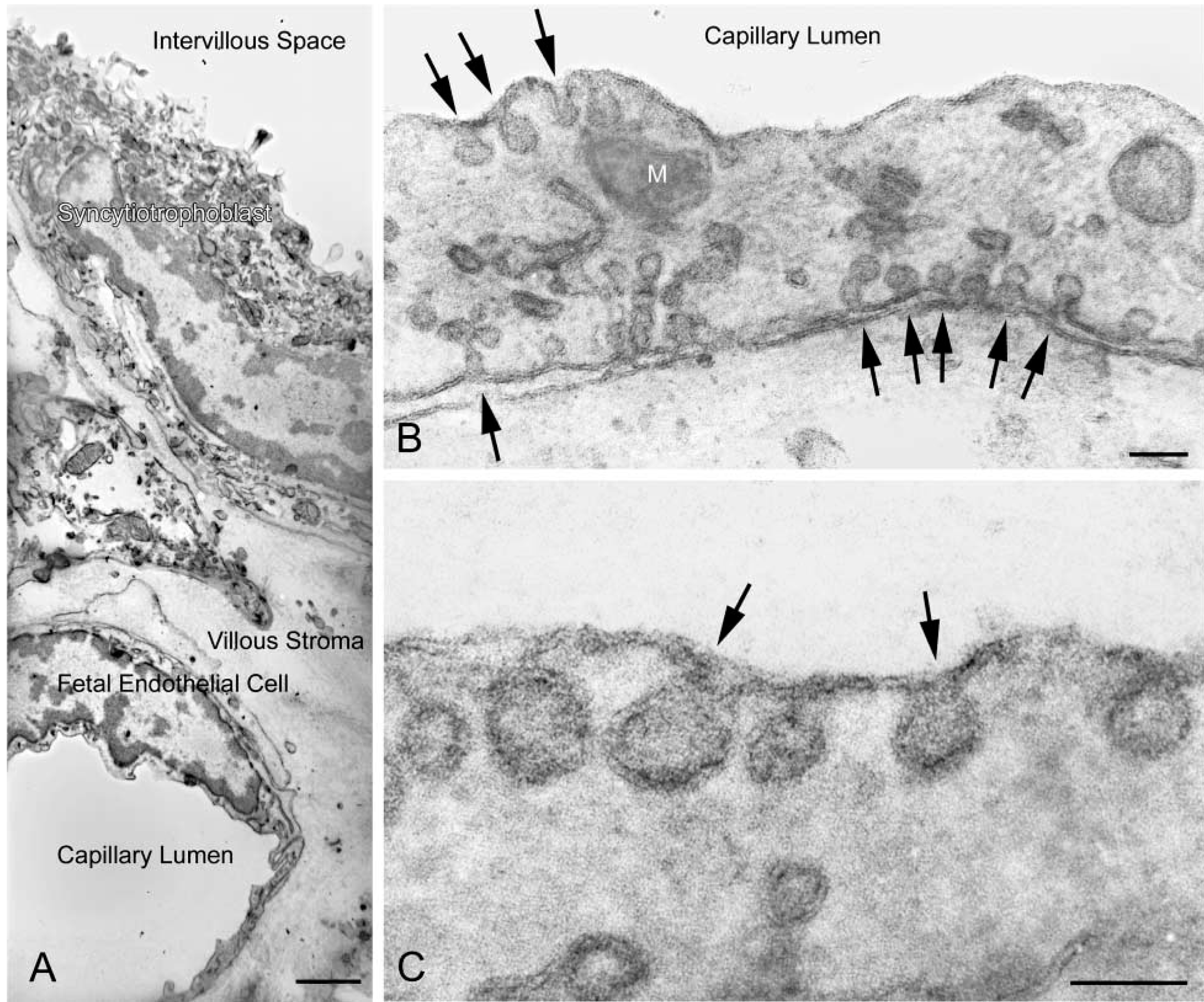
Reduced osmium method. Ultrathin cryosections of 4% PFA-fixed human full-term placenta were postfixed with ferrocyanidereduced osmium and stained with uranyl acetate and lead citrate in PVA. (
The reduced osmium method provides a positive image of membranes, whereas with the UA-MC method membranes have negative contrast. The contrast enhancement method investigators choose will probably be due to personal preferences. However, positively stained images may be more recognizable and comprehensible, especially to the non-specialist. Most importantly, this methodology is compatible with the use of colloidal gold particles as the reporter system for immunoelectron microscopy. The use of ferrocyanide-reduced osmium in electron microscopy was introduced by De Bruijn (1968) and Karnovsky (1971). This method, as a post-fixative, has been applied in a number of studies to enhance contrast of cell membranes and glycogen before embedding in resin for conventional electron microscopy (e.g., Robinson et al. 1982). To the best of our knowledge, this report is the first time that ferrocyanide-reduced osmium has been used with ultrathin cryosections. De Bruijn (1995) summarized the proposed chemical mechanism that leads to improved contrast of membranes with reduced osmium in conventional electron microscopy. We assume that enhancement of contrast in ultrathin cryosections from tissue fixed in PFA and subsequently treated with ferrocyanide-reduced osmium, and with lead citrate and uranyl acetate in PVA, occurs in a manner similar to that previously discussed (De Bruijn 1995). The approach to electron microscopic examination of immunolabeled-ultrathin cryosections described here should prove to be of value in studies using ultrathin cryosections.
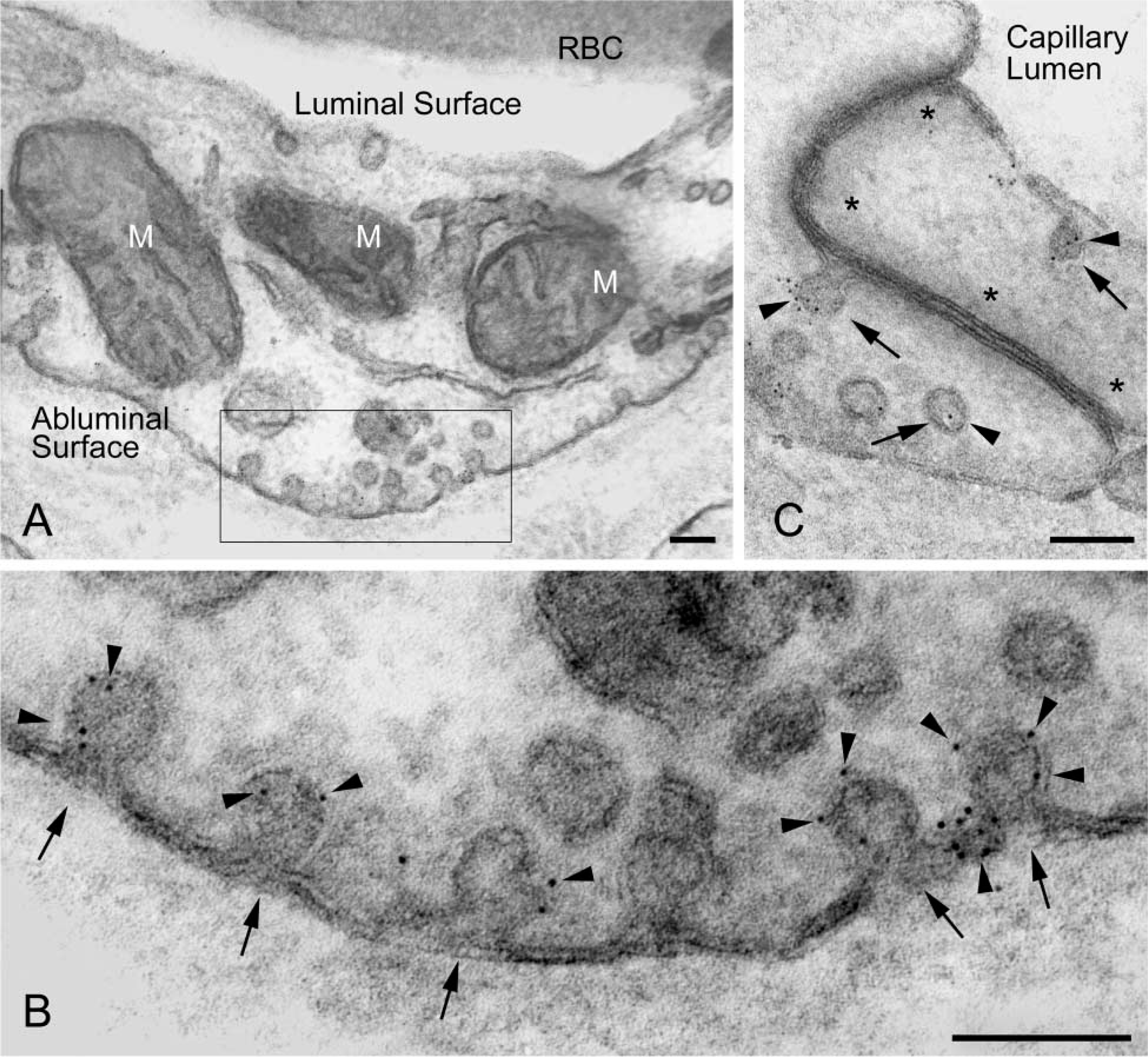
Immunoelectron microscopic localization of CAV-1α in a terminal villus of human placenta. Cryosections of tissue fixed in 4% PFA and postfixed in ferrocyanide-reduced osmium before staining with uranyl acetate and lead citrate in PVA. (
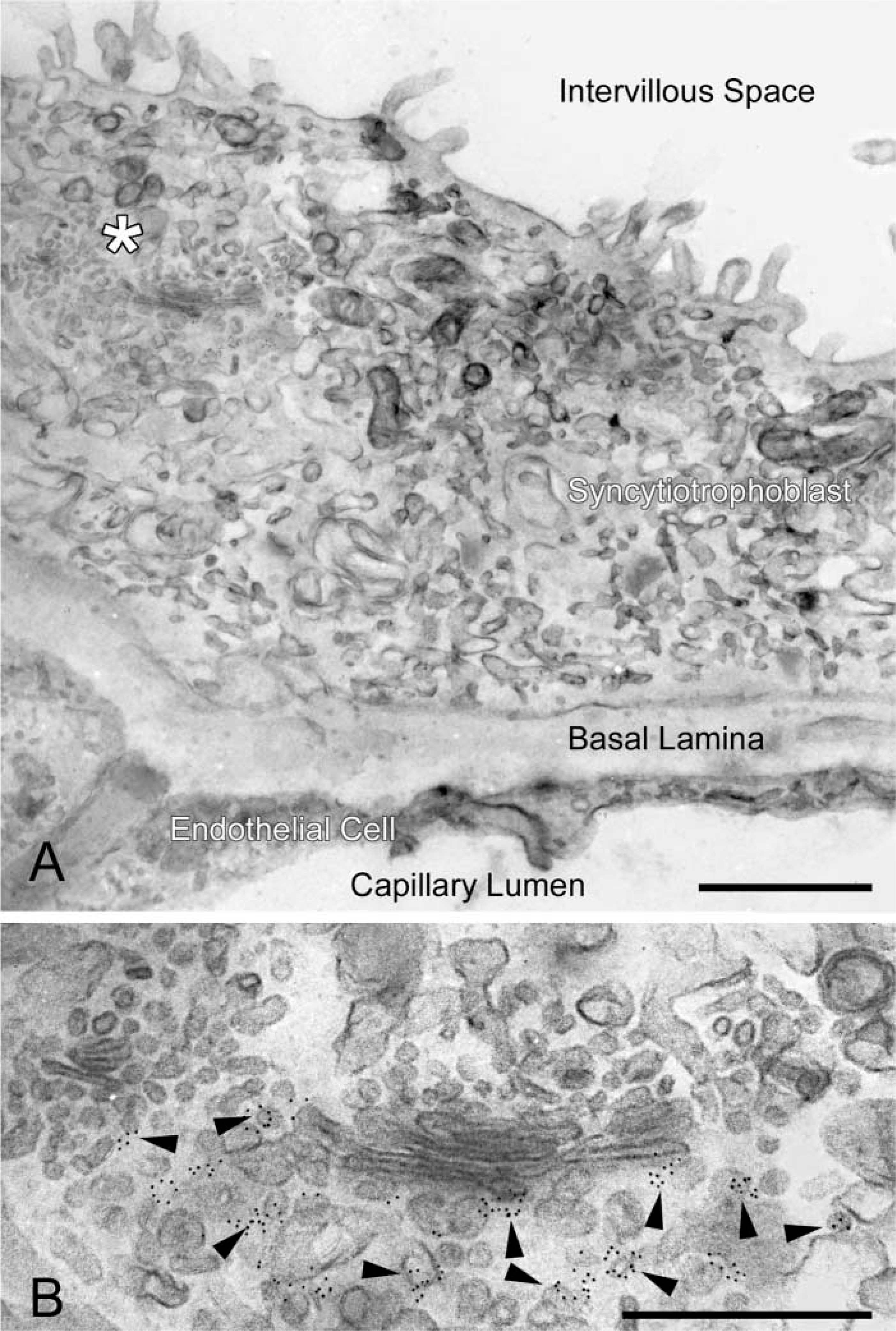
Immunoelectron microscopic localization of p230 in the trans-Golgi network in a terminal villus of human placenta. A cryosection of tissue fixed in 4% PFA and post-fixed in ferrocyanide-reduced osmium before staining with uranyl acetate and lead citrate in PVA. (
Footnotes
Acknowledgements
Supported in part by grants HD38764 (JMR) and CA88053 (CLA) from the National Institutes of Health.
We are indebted to the Campus Microscopy and Imaging Facility at OSU for assistance. We also wish to thank Heather Richard for technical assistance.