
Abstract
Caveolin-1 is a scaffolding protein component of caveolae, membrane invaginations involved in endocytosis, signal transduction, trans- and intracellular trafficking, and protein sorting. In adult lung, caveolae and caveolin-1 are present in alveolar endothelium and Type I epithelial cells but rarely in Type II cells. We have analyzed patterns of caveolin-1 expression during mouse lung development. Two caveolin-1 mRNAs, full-length and a 5′ variant that will translate mainly into caveolin-1 α and -β isoforms, are detected by RT-PCR at embryonic day 12 (E12) and afterwards in the developing and adult lung. Immunostaining analysis, starting at E10, shows caveolin-1α localized in primitive blood vessels of the forming lung, in an overlapping pattern to the endothelial marker PECAM-1, and later in all blood vessels. Caveolin-1 α is not detected in fetal or neonatal lung epithelium but is detected in adult epithelial Type I cells. Caveolin-1 was previously shown to be expressed in alveolar Type I cells. These data suggest that expression of caveolin-1 isoforms is differentially regulated in endothelial and epithelial cells during lung development. Caveolin-1α is an early marker for lung vasculogenesis, primarily expressed in developing blood vessels. When the lung is fully differentiated postnatally, caveolin-1 α is also expressed in alveolar Type I cells.
Keywords
C
Although caveolae are present in most tissues, caveolin family members are differentially expressed in various cell types. Caveolin-1 mRNA is highly abundant in adult adipose tissue and lung, and is detected at lower levels in other tissues (Scherer et al. 1996). Caveolin-2 is co-expressed with caveolin-1 (Scherer et al. 1996) but may have a different subcellular localization (Mora et al. 1999), and caveolin-3 is specifically expressed in muscle (Tang et al. 1996). There are two isoforms of caveolin-1 protein (α and β) (Scherer et al. 1995). It was recently shown that in mice these isoforms are the main product of different RNAs (full-length and 5′ variant) resulting from alternative transcription initiation sites, which have a distinct first exon and also can differ in 620 bp of their 3′ UTR (Kogo and Fujimoto 2000). These two proteins (α-isoform 24 kD and β-isoform 21 kD) are produced by different translation initiation sites, 32 aa apart (Scherer et al. 1995). Using an antibody that recognizes both isoforms (α and β), caveolin-1 proteins are detected in the adult lung, mainly in alveolar epithelial Type I cells and alveolar capillary endothelial cells (Newman et al. 1999). We now show that, during lung development, the caveolin-1α isoform is expressed in the vasculature but not in the epithelium. In adult lung, expression is detected in endothelial blood vessel cells and alveolar epithelial Type I cells.
The process of vasculogenesis in the lung begins when the lung buds from the foregut on embryonic day 9–9.5 (E9–9.5) (Schachtner et al. 2000). It is presumed that primitive hemangioblasts of mesodermal origin differentiate to form a vascular plexus that will become the peripheral lung vasculature. Vasculogenesis has been suggested to be the main mechanism of proximal and distal blood vessel formation during development (Schachtner et al. 2000). In the perinatal period, as alveolar septae thin, the alveolar capillaries are remodeled to form a network closely apposed to the epithelium to facilitate gas exchange. Very little is known about the molecular regulators that control the development of the extensive alveolar capillary bed. VEGF and its receptors (flk-1, flt-1, and flt-4) are, among others, important regulators of lung vasculogenesis and angiogenesis (Gale and Yancopoulos 1999; Schachtner et al. 2000). The role of these molecules in vascularization of the lung is now under intense study.
In contrast, there is considerable information about molecules that influence lung epithelial development, including, e.g., many known growth factors and signaling molecules (Shannon and Deterding 1997). When development and cell differentiation are complete, several populations of epithelial cells with distinctive phenotypes line the airways and alveoli. Our previous studies have been focused on understanding the molecular mechanisms that control the alveolar Type I cell phenotype in the lung (Williams et al. 1996; Ramirez et al. 1999). Caveolin-1 is expressed in primary alveolar epithelial Type II cells cultured in monolayers on plastic plates (Campbell et al. 1999) following the pattern of the specific Type I cell markers T1α (Borok et al. 1998a) and aquaporin-5 (Borok et al. 1998b). These three genes are quickly upregulated after a day in culture, whereas Type II cell genes (e.g., surfactant apoproteins) are downregulated.
Previous studies of caveolin expression during mouse development were performed by Northern and Western blotting analysis using RNA or protein extracts from whole embryos starting at day E7. Although these data show that caveolin-1 is expressed very early in mouse embryos, they are uninformative about expression sites in different organs and/or cell types (Engelman et al. 1998a). In the same report, the authors show by immunohistochemistry that at E16 caveolin-1 is expressed in lung parenchyma and in bronchiolar epithelium. However, the specificity of the antibody for various caveolin species used in that experiment is not stated.
The current knowledge about transcriptional and/or post-transcriptional regulation of caveolin-1 expression is limited. Promoter studies show that two sterol regulatory elements mediate the upregulation of caveolin-1 gene transcription in response to increased levels of intracellular free cholesterol (Bist et al. 1997). It is not known what regulates expression of the α- and β-isoforms of caveolin-1. In vitro transcription/translation studies showed that full-length caveolin-1 mRNA lacking the 5′UTR could produce either α- or β-isoforms but that a short sequence of the 5′UTR (22 bp) predominantly drives translation of the α-isoform. The 5′ variant mRNA, on the other hand, only produces the β-isoform (Kogo and Fujimoto 2000). It was suggested that α- and β-isoforms can form caveolae of different shapes, but it is not clear if the shape of the caveolae could be linked to a difference in function (Fujimoto et al. 2000).
We have studied caveolin-1 expression patterns to determine how and when caveolin-1 is targeted to lung cells, particularly alveolar epithelial Type I cells. Our previous work about the regulation of T1α (Williams et al. 1996; Ramirez et al. 1999) shows that T1α is expressed early during lung development in the epithelium and is restricted to Type I cells in the adult lung. It is therefore an excellent molecular tool to follow the Type I cell phenotype in vitro and in vivo. However, to identify common regulatory events that direct the Type I cell phenotype overall, we need additional cell-specific markers.
We show that during lung development two mRNAs able to produce caveolin-1α- or β-isoforms are expressed in the lung. Caveolin-1α protein is expressed in the developing vasculature but not in the epithelium. In the adult, expression is detected in endothelial cells and alveolar epithelial Type I cells. Endothelial expression is detected on E10; epithelial expression first appears postnatally. It is likely, therefore, that the regulatory mechanisms of caveolin-1 gene transcription and/or translation are markedly different in endothelial and epithelial cells of the lung.
Materials and Methods
RNA and Protein Purification
Lungs were isolated from E12, E15, E19, postnatal Day 3, 5, 30, and 60, and adult FVB mice (Charles River; Wilmington, MA) (n = 1–3). They were housed in VFB (virus-free barrier) rooms, using methods described earlier (Ramirez et al. 1999) in conformity with the Boston University LASC guidelines for the care of laboratory animals. Total RNA was purified using TRIzol reagent as described by the manufacturer (Gibco; Gaithersburg, MD). Because of the small size of the E12 lungs, four lungs were pooled and RNA was purified using the same method. Lung proteins were extracted using RIPA buffer (150 mM NaCl, 1% NP-40, 0.5% deoxycholate, 0.1% sodium dodecyl sulfate, and 50 mM Tris-HCl, pH 8.0) as described previously (Williams et al. 1996).
RT-PCR
Total RNA (1 μg) was reverse-transcribed using AMV-reverse transcriptase (Promega; Madison, WI) following the manufacturer's protocol in a final volume of 25 μl. Oligonucleotides to amplify the 5′ end of the full-length and 5′ variant caveolin-1 mRNAs were ex2/R common reverse primer and a-start/F or 5′ V/F1 as forward primers described previously by others (Kogo and Fujimoto 2000). The PCR reaction was performed with 2 μl of the RT reaction, Q solution, and polymerase from Qiagen (Valencia, CA) (94C 1 min; 45C 1 min; 72C 2 min; 30 cycles).
Western Blots
Protein extracts from E15, E19, and adult lung (10 μg) (n=2) were electrophoresed on a 10% polyacrylamide gel by standard methods (Williams et al. 1996). Equal amounts of protein, measured by the Bradford method (Bradford 1976) and loaded in the gel, were used to normalize the data. After transferring gels to nitrocellulose membranes, blots were blocked and incubated overnight at 4C with a 1:1000 dilution of anti-caveolin-1 (N-20, sc-894; Santa Cruz Biotechnologies, Santa Cruz, CA) or 1:500 dilution of anti-PECAM-1 (M-20; Santa Cruz Biotechnologies) in Tris-buffered saline (TBS). After washing three times for 5 min with TBS containing 0.05% Tween-20 (TBST), blots were incubated for 2 hr at room temperature with alkaline phosphatase-labeled goat anti-rabbit IgG or phosphatase-labeled donkey anti-goat IgG diluted 1:6000 in TBST buffer. The blot was developed using a color-forming alkaline phosphatase substrate as described previously (Williams et al. 1996).
Immunostaining Analysis
Mouse embryos and fetal and adult lungs (n = 1–3) were fixed in 4% paraformaldehyde by immersion for 18 hr and embedded in paraffin by standard methods (Williams et al. 1996). Sections (6 μm) were placed on Superfrost/Plus slides (Fisher Scientific; Pittsburgh, PA). Deparaffinized sections were incubated in Vector unmasking solution (Vector Laboratories; Burlingame, CA), for 20 min at 90C for antigen retrieval. Endogenous peroxidase was quenched and normal serum was used as blocking protein. Sections were incubated with anti-caveolin-1 (see above) at 1:40,000 dilution, 16 hr at 4C. The N-20 antibody was raised against an N-terminal peptide of caveolin-1 (aa 2–21). Specific binding was confirmed by incubation of the sections with anti-caveolin-1α (N-20) antibody in the presence of a five- or 10-fold excess of the blocking peptide (aa 2–21) (Santa Cruz Biotechnology). Immunostaining was performed with a Vectastain Elite ABC kit (Vector Laboratories) followed by biotinyl/tyramide signal amplification (TSA-Indirect; NEN Life Science Products, Boston, MA) and visualized using DAB (diaminobenzidine) as a substrate. PECAM-1 staining was done using the same method as caveolin-1. An anti-PECAM-1 (M-20) antibody (Santa Cruz Biotechnology) was used at a 1:1000 dilution. Counterstaining was performed using hematoxylin and eosin as previously reported (Williams et al. 1996) or methyl green nuclear counter-stain (Vector) following the manufacturer's protocol.
Results
Expression of Caveolin-1 mRNAs During Mouse Lung Development
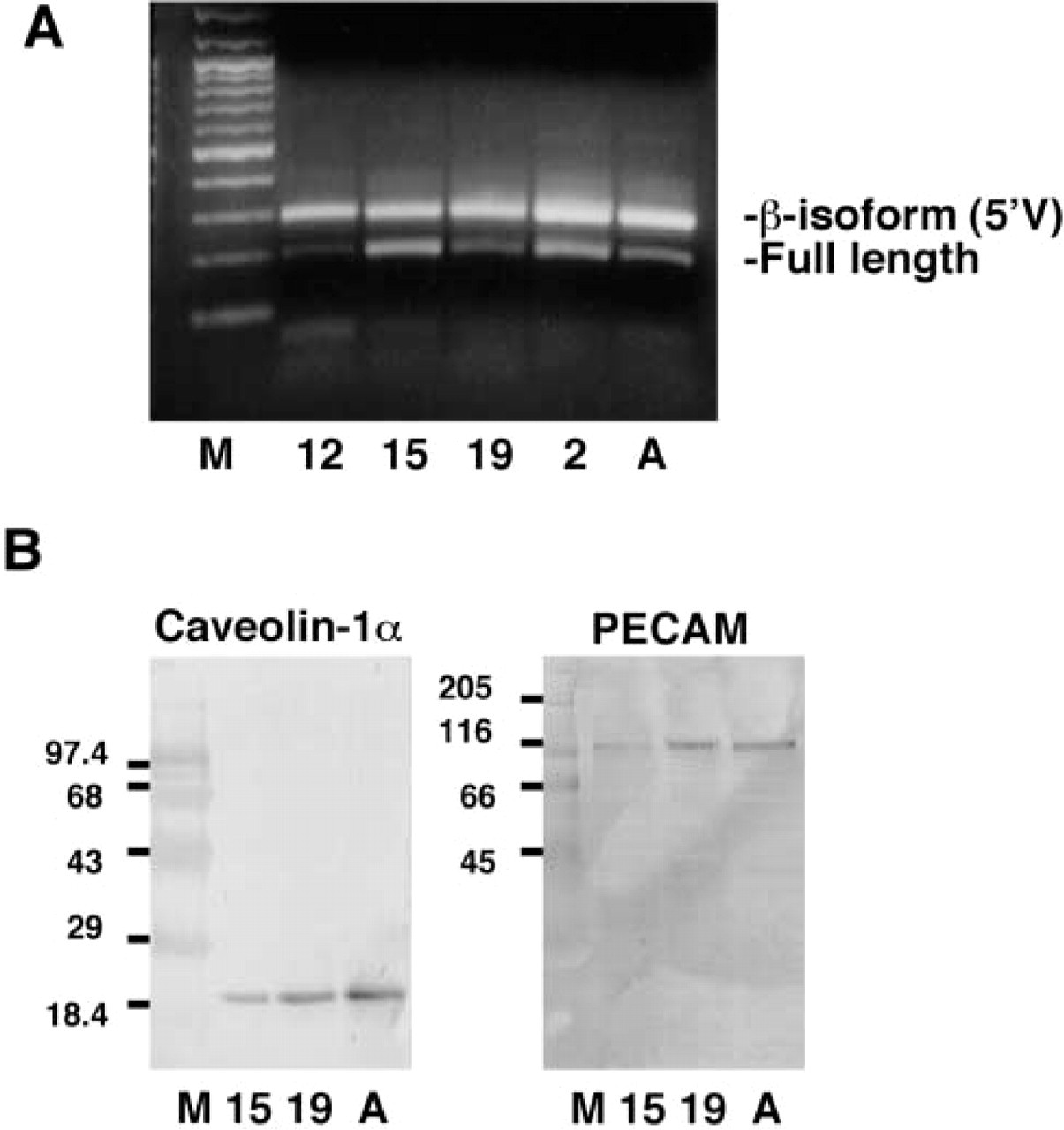
Two messages for caveolin-1 that can be translated mainly into the α- or β-isoforms have been recently described (Kogo and Fujimoto 2000). We used RT-PCR to determine the onset of expression of these mRNAs in the developing lung, starting at Day E12 (the lung bud forms in the mouse on Day E9.5) (Figure 1A). The oligonucleotides used amplify fragments of both caveolin-1 mRNAs from the 5′UTR to the end of the second exon, resulting in amplicons of 198 and 293 bp. We found that both fragments were easily detectable on day E12. Earlier studies have reported that caveolin-1 mRNA can be detected in the whole mouse embryo on day E7 (Engelman et al. 1998a). It has also been reported that adult mouse lung expresses both the full-length and 5′ variant mRNAs (Kogo and Fujimoto 2000).
Western Blotting Analysis of Caveolin-1α and PECAM-1 Protein Expression During Mouse Lung Development
Caveolin-1α protein has a unique N-terminal sequence against which the anti caveolin-1 (N-20) antibody was raised (aa 2–21). Western blots using this antibody (Figure 1B) show only one band in the lung
Caveolin-1 expression in developing and adult lung. (
at every developmental time studied; we conclude that this band is caveolin-1α (24 kD). We can detect increasing levels of caveolin-1α protein in E15, E19, and adult lung extracts. This increasing pattern of expression matches that of PECAM-1 (Figure 1B). These Western blots indicate that the antibodies used in this study are specific for the proteins of interest.
Immunohistochemical Analysis of Caveolin-1 Protein Expression in Embryonic and Adult Lung
Using the N-20 (sc-894) antibody, we determined the expression pattern of caveolin-1α protein in embryonic, fetal, postnatal, and adult lung tissue sections. Analyses were performed in sections from one to three embryos or adult lung.
Whole Embryo at E15. To obtain an overview of caveolin-1α expression in mouse fetus, we examined staining patterns of sagittal sections of an E15 mouse embryo (Figure 2). At this developmental stage, caveolin-1α is mainly, if not entirely, expressed in endothelial cells and the staining patterns clearly outline the vasculature in many organs. Along with the kidney (Figure 2A) and gut (Figures 2A and 2F), the lung appears to express the highest levels of caveolin-1α (Figures
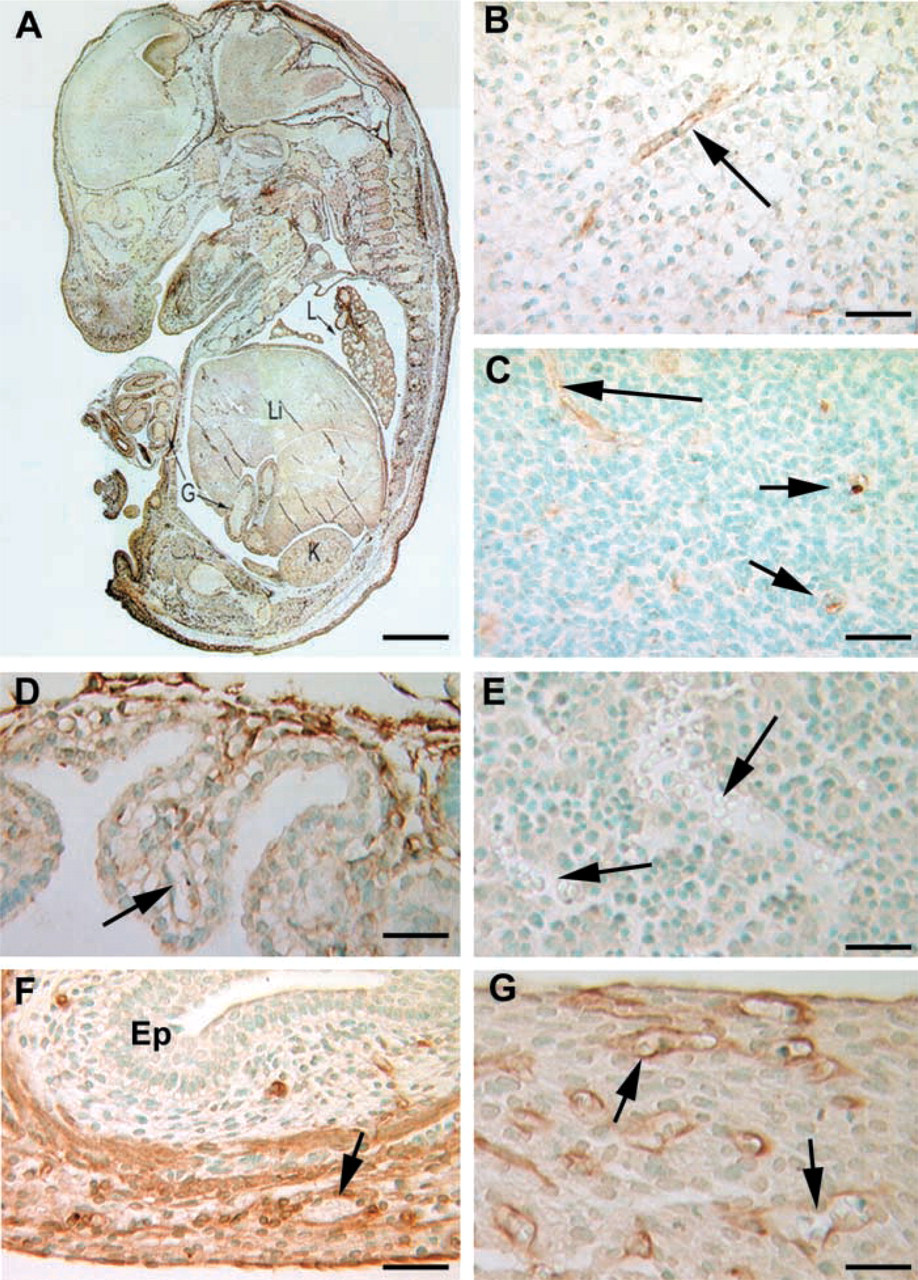
Caveolin-1α immunostaining at Day E15 of mouse development. Sagittal sections of a 15-day embryo were stained for caveolin-1α. (
2A, 4D, and 4E). Endothelial cells of the heart are also positive (Figure 2G). There is no detectable expression in liver sinusoids (Figure 2E), although caveolin-1α was shown to be expressed in adult rat liver endothelium by an electron immunogold method (Yokomori et al. 2000). In the brain, a few forming vessels in the white and gray matter express low levels of caveolin-1α (Figures 2B and 2C). The choroid plexus, rich in capillaries, does not show any stained cells (Figure 2D) at this developmental time point. Blood vessels in the forming meninges also stained positive for caveolin-1α (Figure 2A).
Lung Tissues
Embryonic Day 10. Immunohistochemical studies of the lung at this developmental stage were performed on serial sections. Caveolin-1α is expressed by clustered cells scattered throughout the lung mesenchyme (Figure 3A). Careful examination of the stained areas shows that each cluster appears to be a small vessel with a discernible lumen (Figures 3D and 3E). Staining of serial sections using anti-PECAM-1, a marker for endothelial differentiation (Schachtner et al. 2000), indicates that caveolin-1α is indeed expressed in forming blood vessels (Figure 3C). No specific staining is observed in the epithelial bud (Figures 3A and 3B). In competition assays, incubation with anti-caveolin-1 antibody in the presence of a 10-fold excess of the blocking peptide (aa 2–21) shows that staining of the forming vessels is specific (Figure 3B). Binding of the antibody to the forming vessels can be prevented by incubation with the soluble specific peptide, but staining of the pleura and some regions of the apical membrane of the pulmonary epithelium cannot be blocked, confirming that epithelial and pleural staining is nonspecific. The nonspecific nature of the pleural and epithelial staining is also confirmed by staining with secondary antibody alone (Figures 3F and 3G). The nonspecific staining of some areas of the epithelium and pleura was also observed at other developmental time points (Figures 4F and 4G, Day E15). PECAM-1 has been shown to be expressed in lung endothelial cells starting at E11 (Healy et al. 2000). Here we extend those studies and report that PECAM-1 is expressed a day earlier (E10) (Figure 3C).
Embryonic Day 12. By Day E12 (Figures 4A–4C), the expression of caveolin-1α is highly localized to blood vessels and is not detected in epithelium. Vessels of all sizes are positive, including capillaries of the presumptive peripheral lung and large vessels. There is often
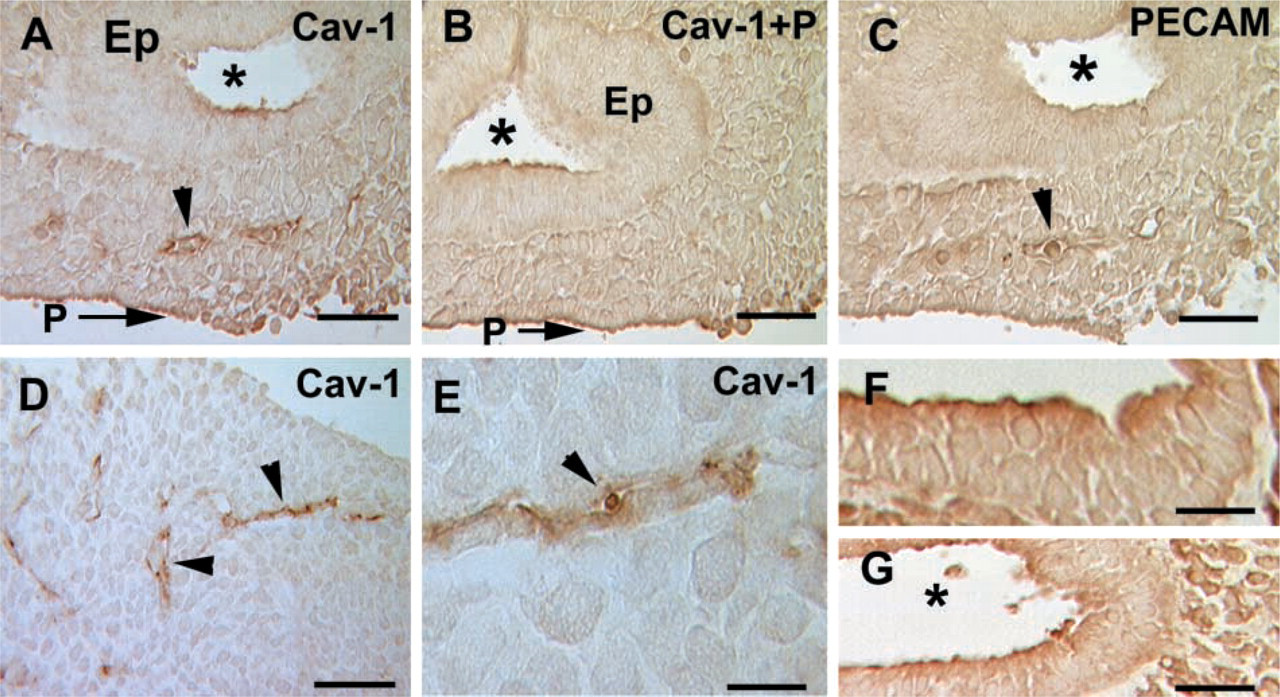
Immunostaining of caveolin-1α and PECAM-1 in E10 lung. (
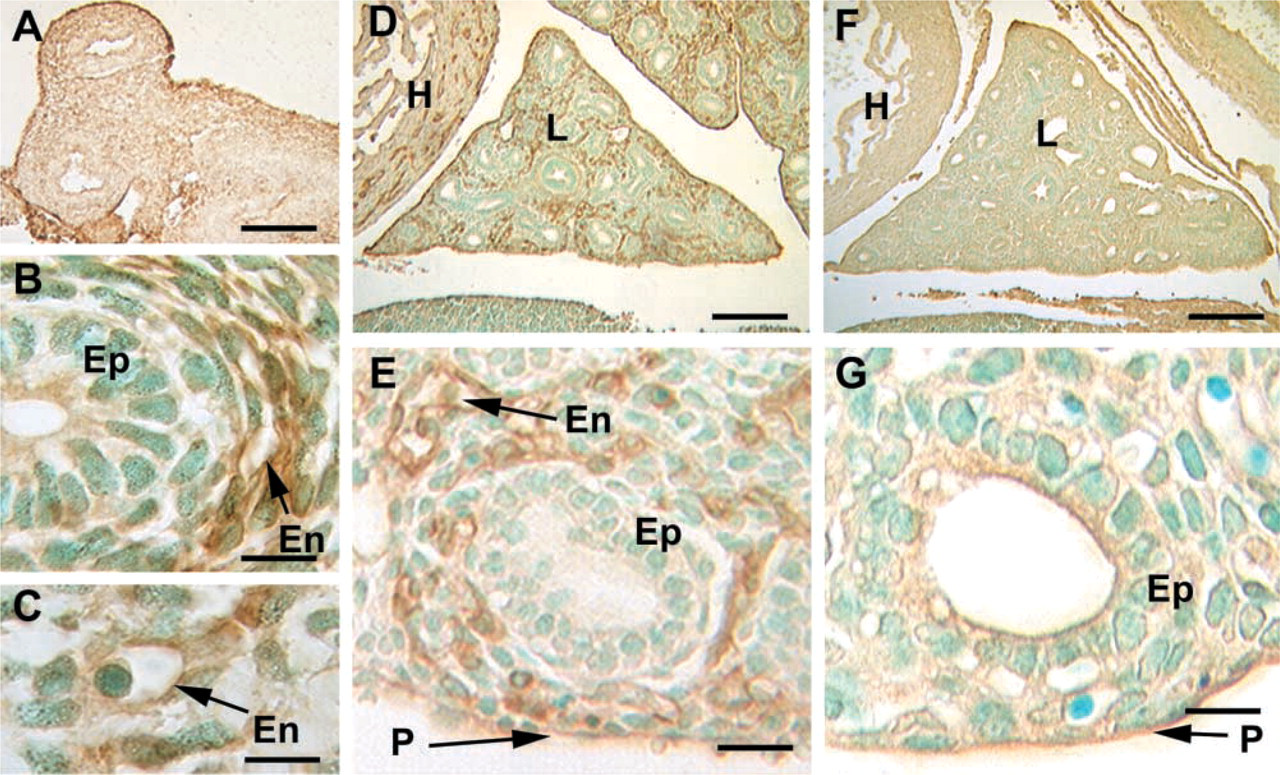
Immunostaining of caveolin-1α in E12 and E15 fetal lung. (
a well-defined ring of small vessels circumferentially arranged around the developing bronchioles (Figure 4B). Other mesenchymal cells, including smooth muscle, are negative.
Embryonic Day 15. Analysis of lung at higher magnification shows that the epithelium and mesenchyme are negative but intense staining is observed in the forming blood vessels, including capillaries (Figures 4D and 4E). Nonspecific apical staining of some areas of the epithelium and the pleura was also observed at this developmental time point (Figures 4F and 4G).
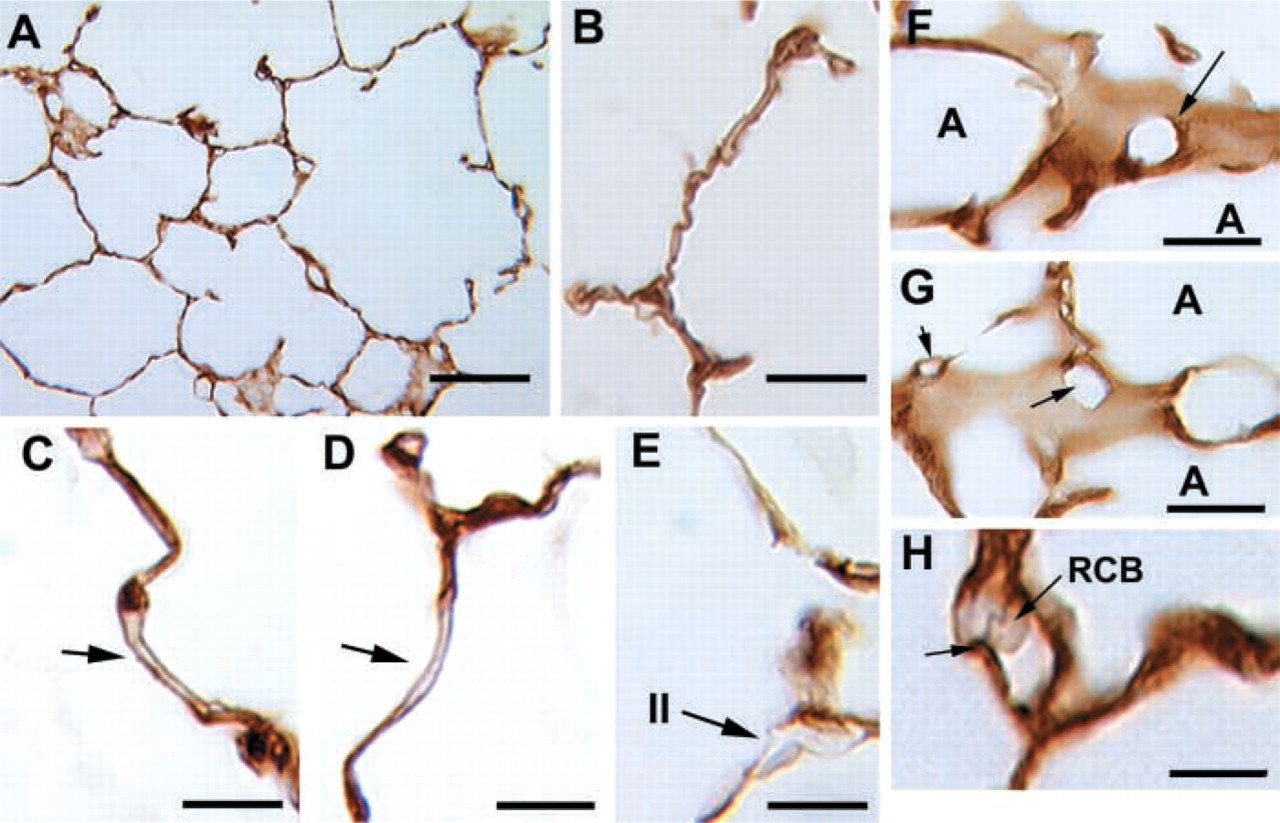
Embryonic Day 19. Towards the end of gestation (Figures 5A–5C) the peripheral capillary bed is intensely reactive for caveolin-1α. Although morphogenesis of Type I cells is ongoing and the cells express the Type I cell marker proteins T1α and Aqp-5 by this time (Ramirez et al. 1999, 2000) there is no detectable caveolin1-α expression in either bronchial or distal epithelia. Counterstaining with hematoxylin and eosin shows that only endothelial cells express caveolin-1α, whereas muscle cells surrounding either upper airways (Figure 5D) or blood vessels (Figures 5E and 5F) are negative.
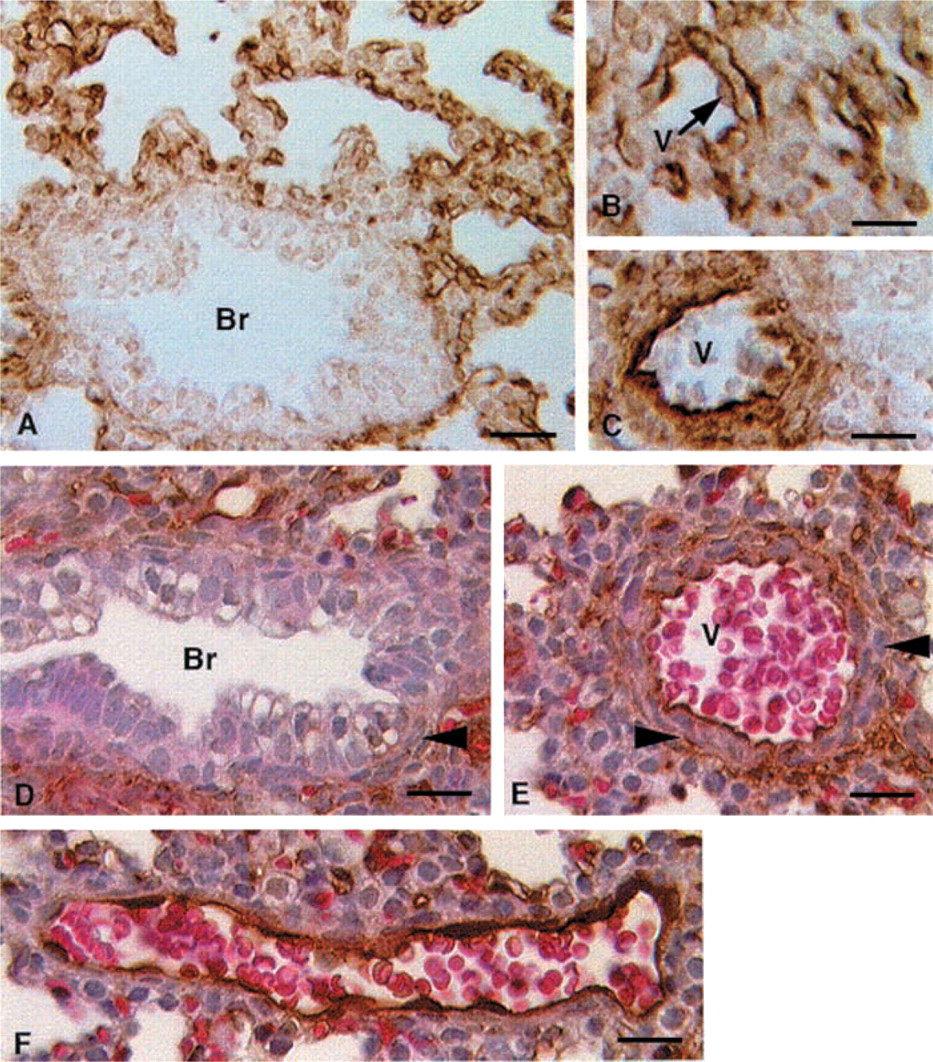
Neonatal and Adult Lung. Endothelial staining is readily detected on postnatal Days 1, 3, 5, 30, and 60 (not shown) but epithelial staining is not evident, if present. In adult lungs (Figure 6A), however, there is continuous staining of the alveolar walls in a pattern that matches the expression of the type I cell marker T1α (Williams et al. 1996), indicating that Type I cells express caveolin-1, as shown previously by others using different antibodies. Higher magnification confirms the continuous staining on both sides of the alveolar septae (Figures 6B–6D), consistent with the known location of type I cells. In contrast, Type II cells (Figure 6E) appear not to express detectable amounts of caveolin-1α, confirming the results of Campbell et al. (1999). Tangential sections of the adult lung show that blood vessels in the adult distal lung stain positive for caveolin 1α (Figures 6F–6H). Control sections using secondary antibody alone show no staining in the distal lung (data not shown).
Immunostaining of caveolin-1α in E19 fetal lung. (
Discussion
We present here studies of the developmental pattern of expression of the caveolin-1 gene in normal mouse lung, from early development until adulthood, focusing in the temporal patterns of expression of caveolin-1 protein in endothelium and epithelium. Most of the studies about caveolin-1 published to date describe the functions and distribution of this gene in adult tissues or in cell lines. There is minimal information about expression of caveolin-1 during mouse development, and the patterns of expression of the two isoforms of caveolin-1 protein during development have not been previously reported.
The lung is one of the organs that expresses the highest levels of caveolin-1 mRNA in rodents (Scherer et al. 1996). Engelman et al. (1998a) have shown expression of caveolin-1 protein in mouse lung at Day E16, the earliest time point they examined, in parenchyma and epithelium. Others reported that caveolin-1 expression is first detected in the lung on E19 (Kasper et al. 1998). Although these results appear to be discrepant, the antibodies used by these investigators may recognize different epitopes and/or there may be different sensitivities due to particular methodologies. It is important to point out that in both reports the antibodies used do not differentiate between the α- and β-isoforms of caveolin-1. Our findings show that two caveolin-1 messages, which can be translated into caveolin-1α and -β proteins, are present in the lung throughout development and that the α-isoform of caveolin-1 is expressed as soon as the lung buds only in endothelial precursors that form the initial network of pulmonary blood vessels.
Considering that the lung starts budding from the foregut endoderm at day E9.0–9.5, we show expression of endogenous endothelial markers (PECAM-1 and caveolin-1α) at the earliest developmental time point of the forming lung studied to date. Another early marker of lung vasculogenesis may be flk-1. Although the endogenous gene pattern in the budding lung was not studied (Schachtner et al. 2000), lac Z
Immunostaining of caveolin-1α in adult mouse lung. (
expression in flk-1-lacZ mice was detected in the forming lung vasculature at Day E10.5.
Caveolin-1α may therefore be involved in lung endothelial cell differentiation and/or function during vasculogenesis. Early expression of caveolin-1α in the vascular plexus suggests that the caveolin-1 gene could be upregulated by factors that trigger vasculogenesis. Vascular endothelial growth factor (VEGF) and its receptors (Flk-1, Flt-1, and Flt-4) have critical roles in vasculogenesis: Mice carrying null mutations for either Flk-1 receptor or VEGF are deficient in blood vessels and in the specification of endothelial precursors (Gale and Yancopoulos 1999). In a human endothelial cell line, caveolin-1 protein is downregulated by VEGF, and downregulation is abolished by treatment with angiogenesis inhibitors (Liu et al. 1999). This result does not necessarily rule out the possibility that VEGF upregulates caveolin-1 during vasculogenesis, when cells are differentiating from hemangioblasts. Although caveolin-1α is expressed in endothelial cells early during lung development, whether caveolae form in primitive endothelial cells is unknown. The functions of caveolae and caveolin proteins in lung development are also unknown. Downregulation of caveolin-1 expression appears to be necessary for enhanced cell proliferation, as shown by studies of both tumor and oncogene-transformed cell lines (Engelman et al. 1998b). However, the endothelial cell population expressing caveolin proliferates rapidly during lung development.
Caveolin-1α expression remains strictly endothelial in the fetal lung. Postnatally, vascular expression persists with little or no expression in the epithelium. In adult mice (>2 months) expression is clearly observed in vessels and in epithelial Type I cells, as also reported by others. Postnatal upregulation of caveolin-1α in lung epithelium appears to take place gradually. The alveolar spaces of the adult lung are lined by thin, flat epithelial Type I cells that are in close contact with capillaries, providing an extremely large surface for gas exchange (Schneeberger 1997). Recently, the phenotype of the caveolin-1 null mutant mouse has been reported (Razani et al. in press). Homozygous mice are viable, although a few homozygous mice die before 9 months of age for unknown reasons. The studies performed by Razani et al. (in press) at 4–5 months of age showed that at that time there are already major alterations in the distal lung. At about that time, we detect expression of caveolin-1α in Type I epithelial cells and in endothelial cells. Alveolar walls of homozygous caveolin-1 null mice are thicker and hyper-cellular, and there appears to be hyperproliferation of alveolar endothelial cells. The authors indicate that there is no expression of either caveolin-1α or -β in null mouse lungs and that caveolae are not formed in alveolar endothelial and Type I epithelial cells. The large number of caveolae and expression of caveolin-1 gene in normal Type I cells and capillary endothelium that form the alveolar wall (Newman et al. 1999), and the phenotype of the null mice, suggest that caveolae and caveolin proteins have a role in the maintenance of alveolar function.
Our studies suggest that α- and β-isoforms of caveolin-1 are expressed in different cell populations during lung development. Caveolin-1α is mostly, if not entirely, expressed in pulmonary endothelial cells before birth, and it does not match the epithelial T1α and AQP-5 gestational expression patterns in the forming lung. Caveolin-1β could be expressed in the lung epithelium during development because expression of caveolin-1 in the epithelium was described at day E16 using an antibody that recognizes both isoforms. It is possible, therefore, that the caveolin-1β expression pattern matches those of other Type I epithelial cell genes. The high homology of these two isoforms and the lack of antibodies, produced in species other than mouse, that recognize only the β-isoform prevent us from characterizing caveolin-1β expression patterns in mouse lung by immunohistochemistry.
There is an emerging interest in caveolae because of their role in transcytosis, endocytosis, and cell signaling. Interaction of several signaling molecules with the scaffolding domain of caveolin proteins may serve as a mechanism to sequester and inactivate signaling molecules and thus regulate their availability (Okamoto et al. 1998). The caveolin-1 null mouse phenotype highlights the importance of these proteins in the adult lung. The role of caveolin-1 in lung development is still not known. Caveolin-1 appears to play a more critical role in the function and homeostasis of the lung after birth than during development. Whether the alterations in the caveolin-1 null lung start in utero or after birth has not been reported. Our findings accentuate the importance of studying caveolin-1 gene regulation by factors that specify hemangioblast differentiation, proliferation, and vessel formation in lung vasculogenesis and to compare endothelial to epithelial caveolin-1 gene regulation. Understanding these issues could clarify whether caveolae in alveolar capillaries and epithelial cells have different roles in the maintenance of alveolar function and/or whether they cooperate to preserve a functional air-blood barrier in the lung.
Footnotes
Acknowledgements
Supported by NHLBI HL47049 and by the Parker B. Francis Families Foundation. MIR is a Parker B. Francis Fellow in Pulmonary Research.