
Abstract
Exhaustive analysis of the location of the peripheral benzodiazepine receptor (PBR) both at the subcellular and the tissue level is warranted to gain a better understanding of its biological roles. To date, many studies have been performed in animal models, such as rat, mouse, and pig, that yielded important information. However, only a few reports were dedicated to the analysis of PBR expression in humans. To enlarge on previous studies, we investigated PBR expression in different human organs using the monoclonal antibody 8D7 that specifically recognized the human PBR. First, we performed electron microscopic analysis that for the first time unambiguously demonstrated the localization of the PBR on the outer mitochondrial membrane. Second, focusing our analysis on human tissues for which information on PBR expression is sparse (lung, stomach, small intestine, colon, thyroid, adrenal gland, pancreas, breast, prostate, ovary), we found that PBR exhibits selective localization. This characterization of PBR localization in human tissues should provide important insights for the understanding of PBR functions.
S
Materials and Methods
Tissue Samples
Samples of various normal tissues were taken from surgical specimen from patients who underwent a surgical cure for cancer at the Montpellier cancer Institute. Normal tissue was removed from the surrounding tumor and was fixed in formalin-alcohol for 24 hr, paraffin-embedded, and subsequently processed with routine techniques and IHC analysis. Normal tissue samples included tissues from the respiratory system (lung, 10 samples), glandular epithelia (colon, 12 samples; small intestine, 8 samples; stomach, 10 samples; breast, 20 samples) and endocrine tissues (thyroid, 10 samples; liver, 10 samples; pancreas, 12 samples; ovary, four samples; prostate, four samples).
Characteristics of MAb 8D7
The anti-human PBR MAb 8D7 was obtained by hybridoma fusion after mouse immunization with the human PBR C-terminal peptide (YHGWHGGRRLPE) conjugated to bovine serum albumin (Dussossoy et al. 1996). In Western blotting experiments, using either crude cell extracts or purified PBR, MAb 8D7 revealed a single band of 18 kD, which corresponds to the expected molecular weight defined by the human PBR cDNA sequence (Dussossoy et al. 1996). Its labeling specificity was previously demonstrated using the peptide competition method that abolished the labeling, by the absence of labeling in Jurkat wild-type cells that are negative for the expression of PBR, and by the specific labeling observed in yeast cells transfected to express the human PBR. No labeling was observed in the corresponding mock-transfected yeast cells (Dussossoy et al. 1996). Reinforcing the specificity of the labeling, MAb 8D7 does not crossreact with rat or mouse PBR, even though those proteins are 80% homologous. Finally, neither DBI nor protoporphyrin, which are two endogenous ligands for PBR, antagonized the antibody binding (Dussossoy et al. 1996).
Electron Microscopy
Human monocytic U937 cells were fixed with 4% formaldehyde and permeabilized with 0.05% saponin. Then the anti-PBR antibody (1:250) was added for 1 hr in 0.05% saponin/1% BSA in NaCl/Pi buffer. Goat anti-mouse IgG (Amersham, Poole, UK; diluted 10-fold) conjugated with 5-nm colloidal gold beads was used for immunogold labeling. Cells were fixed in 2.5% glutaraldehyde for 15 min. Cells were postfixed with OsO4 for 45 min and successively dehydrated with ethanol, propylene oxide and epoxypropylene. Cells were embedded in Epon resin and samples were sectioned (70 nm) with an ultramicrotome (Edelmann 2002). Micrographs were recorded with an electron microscope (JEOL 1010).
Immunohistochemical Analysis
The expression of PBR was analyzed with an IHC procedure. The antibody used was a mouse monoclonal anti-human PBR (Dussossoy et al. 1996) at dilution 1:350. In pancreas, islet cells and sparse endocrine cells were identified by IHC for chromogranin. In these structures, β-cells were identified by IHC for insulin [guinea pig polyclonal antibody (DAKO, Glostrup, Denmark; ready-to-use LSAB2 kit), α-cells by IHC for glucagons (rabbit polyclonal antibody, DAKO; ready-to-use LSAB2 kit), 8-cells by IHC for somatostatin (rabbit polyclonal antibody, DAKO; ready-to-use LSAB2 kit). Two-μm-thick parrafin-embedded sections of tissue samples were analyzed, mounted on DAKO silanized slides. All procedures were carried out at room temperature. IHC detection of the different markers was done using the streptavidin-biotin (LSAB) method (DAKO LSAB kit; Carpinteria, CA). The sections, which had been preincubated with 3% H2O2 solution for 10 min to block endogenous peroxidase, were incubated for 20 min with blocking agent and for 2 hr with the primary antibody. They were next rinsed and incubated with the secondary antibody for 10 min. They were next incubated with streptavidin conjugated to horseradish peroxidase. A positive reaction was visualized with 3-amino-9-ethylcarbazol. Before mounting, the sections were counterstained with Mayer's hematoxylin. For the negative control, the primary antibody was omitted and replaced by an irrelevant antibody (monoclonal mouse anti-human IgG; DAKO). In addition, the specificity of the labeling with PBR was systematically demonstrated by the absence of staining after preadsorbing anti-PBR antibody overnight at 4C with the immunizing peptide (data not shown). The immunoreactivity was then evaluated by two observers using a high-power lens (×400 or ×250).
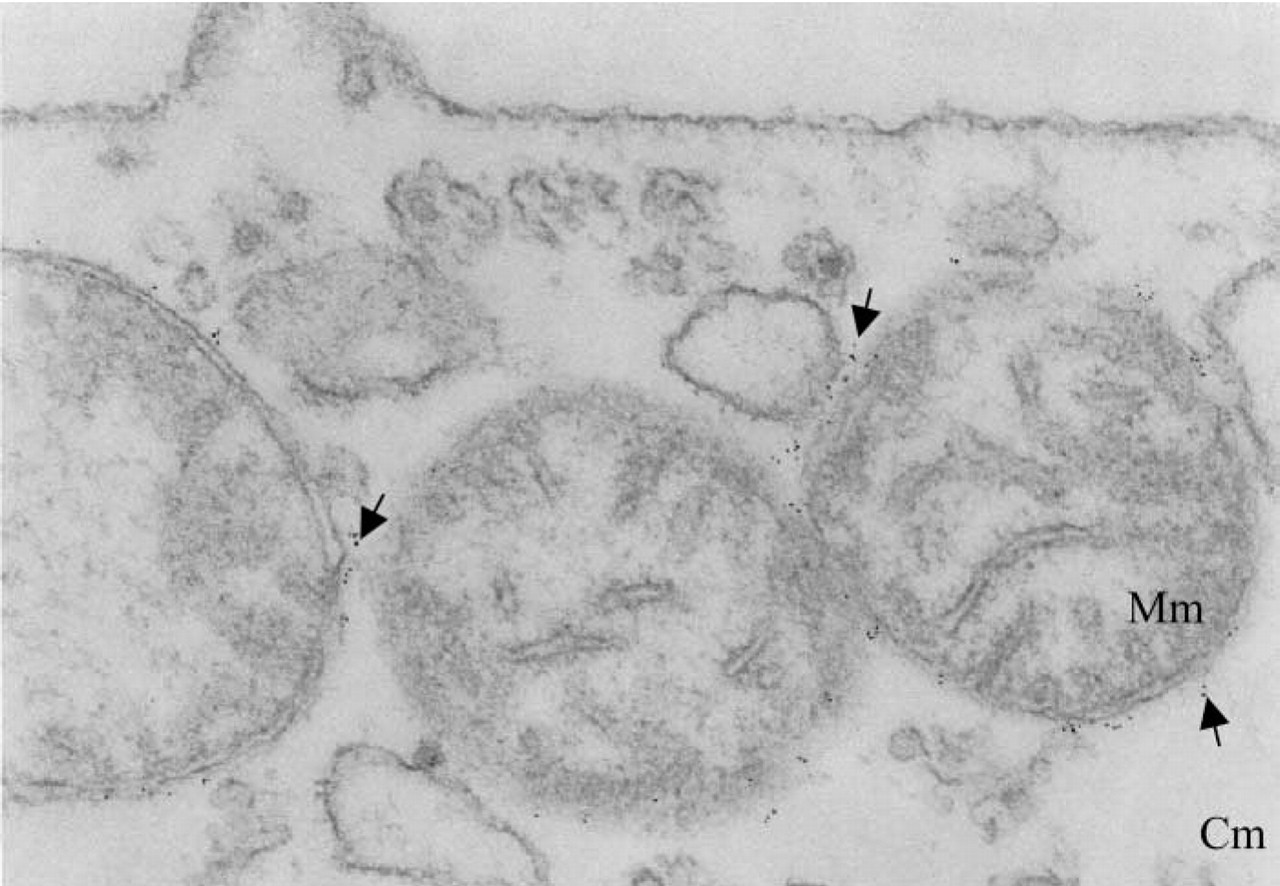
Electron microscopic analysis of localization of PBR on mitochondria in U937 cells. Ultrathin cell sections were labeled with the anti-PBR MAb 8D7, followed by goat anti-mouse IgG conjugated to 5-nm colloidal gold as described in Materials and Methods. Cm, cytoplasmic matrix; Mm, mitochondrial matrix. Arrows indicate colloidal gold particles.
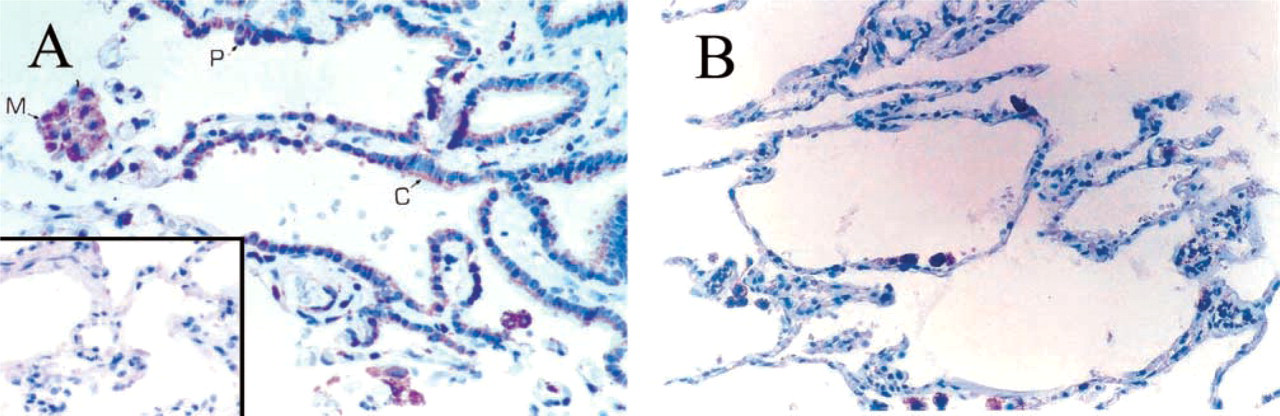
Immunocytochemical staining of human lung with anti-human PBR MAb 8D7. (
Semiquantitative Evaluation of 8D7 Staining
The labeling was evaluated using a semiquantitative method, taking into account the staining intensity and the number of stained cells in different random fields. A score of 0-9 was calculated as the product of the increase in staining intensity compared with the negative controls (0, no labeling; 1, faint staining; 2, moderate staining; 3, strong staining) and the frequency of stained cells (0, less than 10%; 1, 10-25%; 2, 25-50%; 3, more than 50%). Scores were then recorded as an index: index 0 (score 0) meant no PBR expression, index 1 (score 1-2) meant weak PBR expression, index 2 (score 3-4), meant moderate PBR expression, and index 3 (score 6-9) meant strong PBR expression. Highly positive isolated cells were evaluated separately and were not taken into account to measure the expression level of PBR in a tissue.
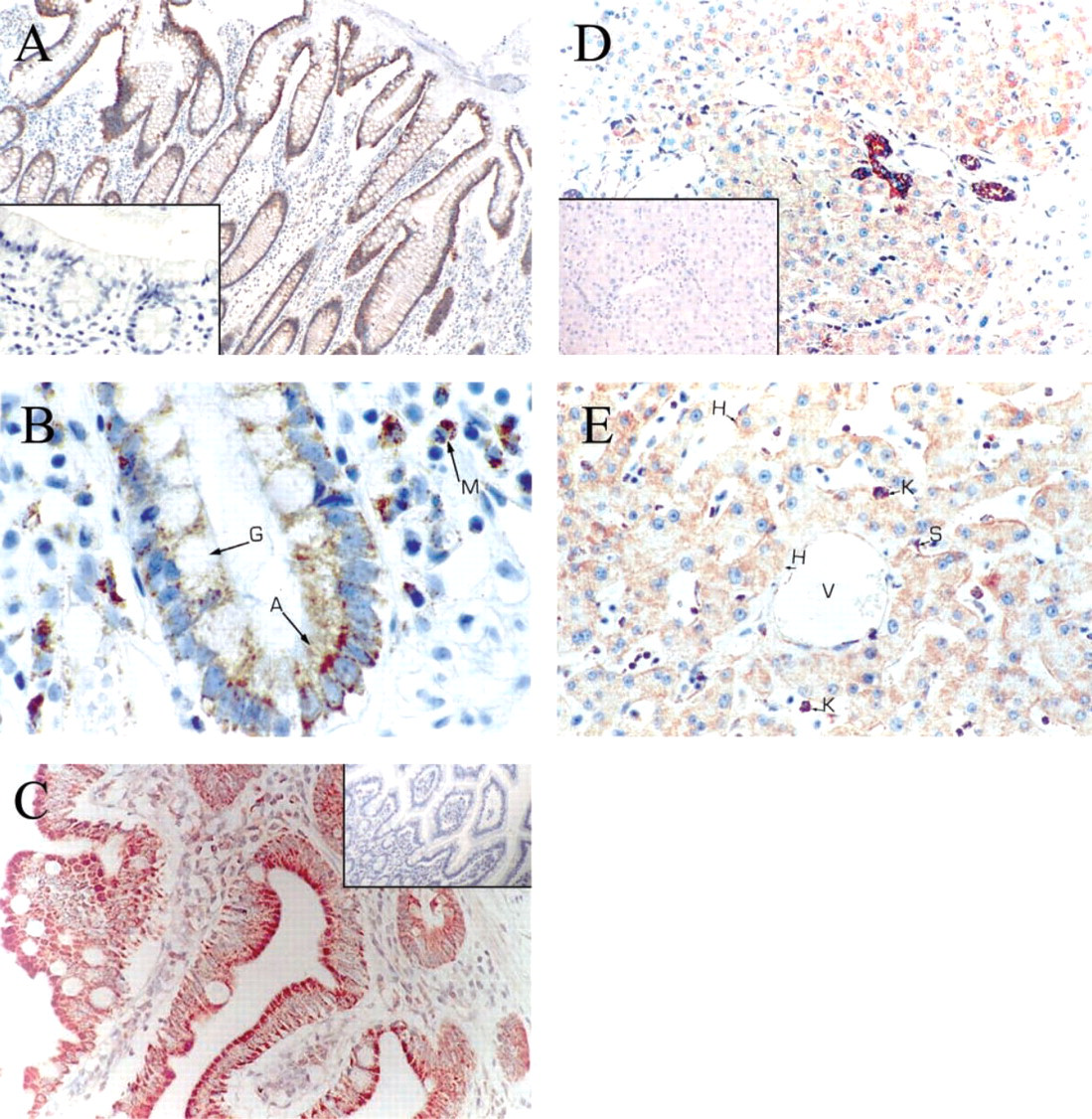
Immunocytochemical staining of human tissues from the digestive tract with the MAb 8D7 anti-human PBR. (
Immunocytochemical staining of human stomach with MAb 8D7. (
Results
Electron Microscopic Analysis of Subcellular PBR Expression
We examined the cellular distribution of PBR at the molecular level using electron microscopy on human monocytic U937 cells. Figure 1 shows representative labeling obtained with the 8D7 anti-human PBR antibody on mitochondria. At × 105,000 magnification, both the double membrane around the mitochondria and the intermembrane space can be distinguished. The gold particles are clearly restricted to the outer mitochondrial membrane. No labeling was observed on other organelles. Neither the cell plasma membrane nor the nucleus was labeled. These results not only evidence the expression of the PBR on mitochondria but also demonstrate unambiguously that the receptor is located on the mitochondrial outer membrane.
IHC Distribution of PBR Immunoreactivity in Normal Human Tissues
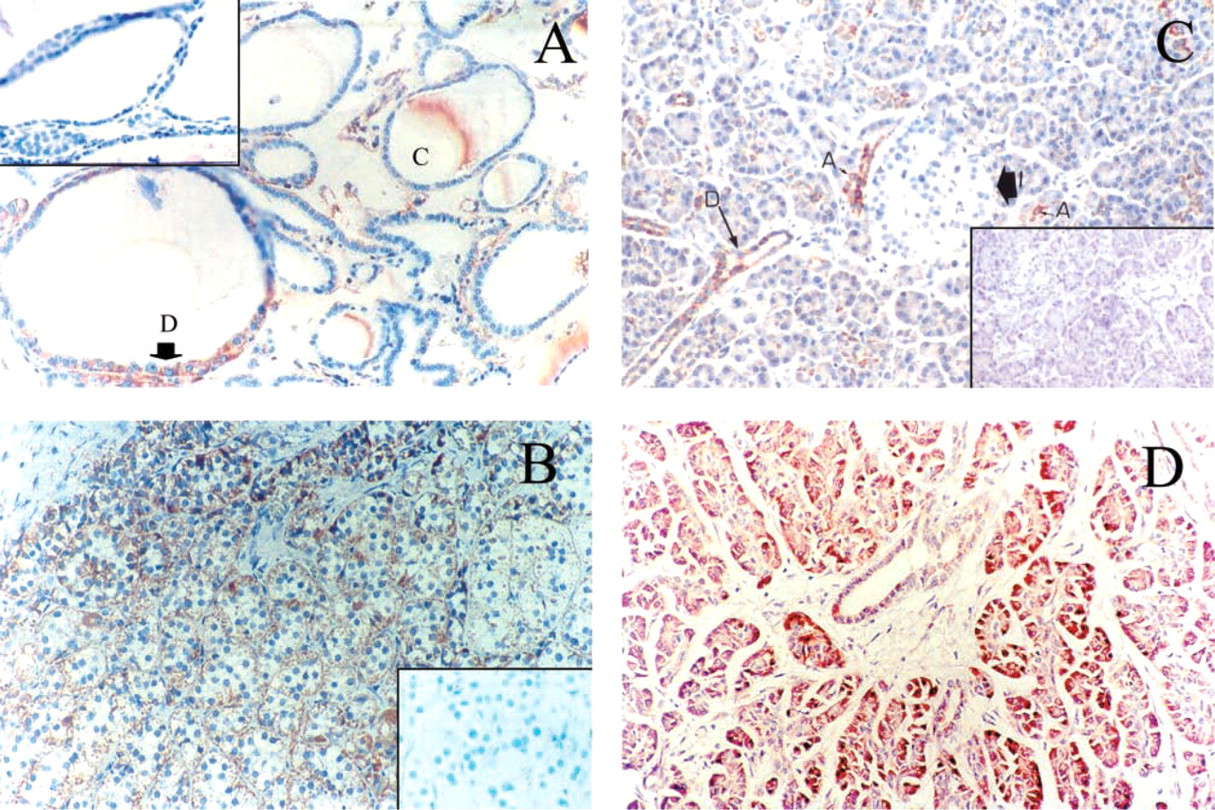
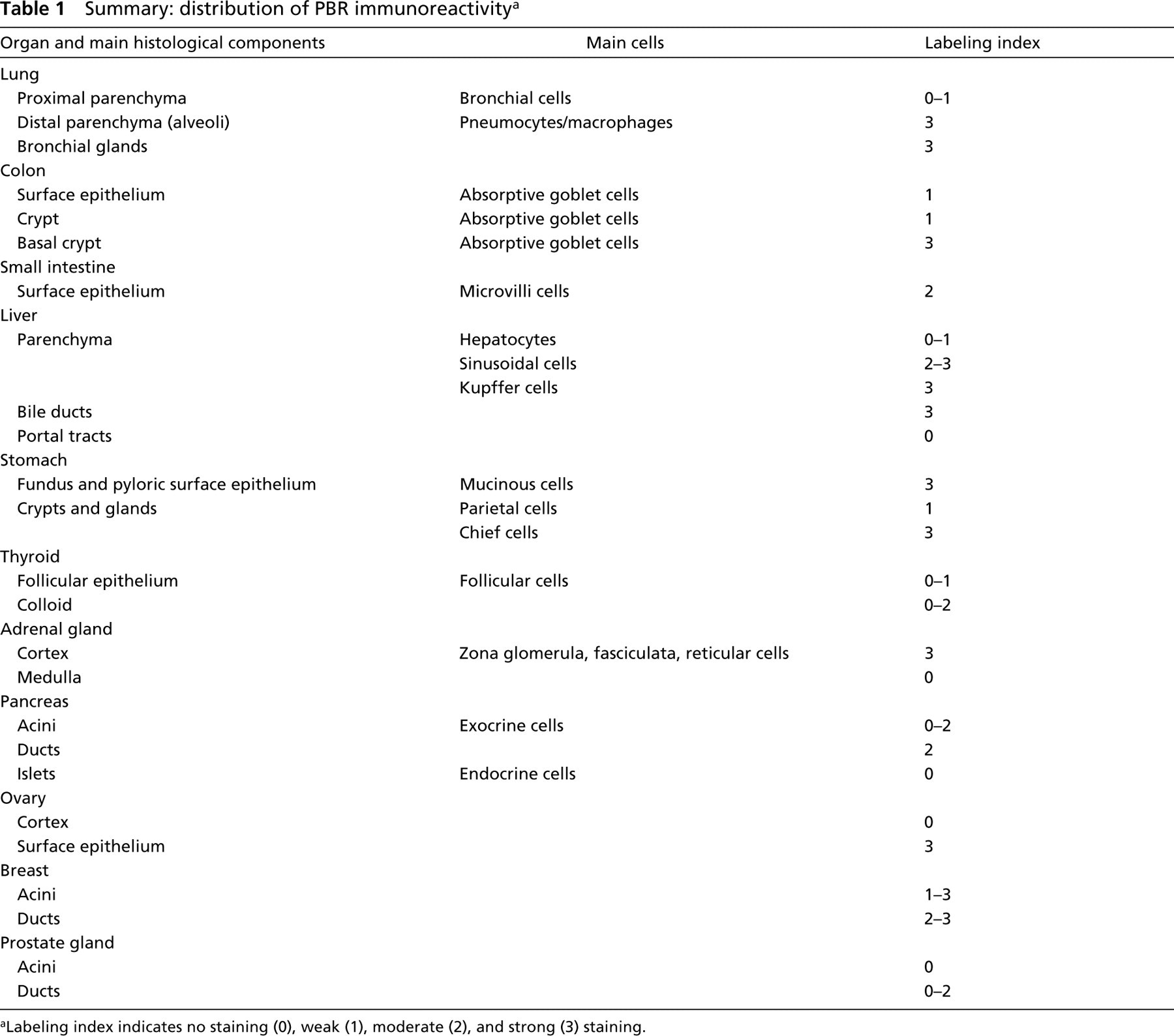
Tissue from the Respiratory System (Figure 2). In the lung, the ciliated bronchial cells showed heterogeneous staining. Most of these cells exhibited no or very weak granular staining, but some cells showed moderate staining, generally located at the apical pole of the cells (Figure 2A). In the distal lung parenchyma, strong immunostaining was observed in some flat cells lining the alveoli; those positive cells are pneumocytes (Figure 2B). Bronchial submucosal glands also exhibited strong cytoplasmic staining around the nucleus and the cytoplasmic membrane, as well as the duct cells. In addition, foamy alveolar macrophages were highly immunoreactive. No staining was seen in the bronchial cartilage.
Tissues from the Digestive System ((Figures 3 and 4). For the colon surface epithelium, both absorptive and goblet cells were weakly positive; the staining was localized on the basal pole (Figure 3A). The columnar epithelium lining the colon crypts was rather faintly stained. In contrast, basal cells at the bottom of the crypts often exhibited strong staining (Figure 3B). As in the colon, the small intestine showed very heterogeneous staining but at the cellular level the staining was regularly distributed (Figure 3C). The positive staining predominated in the villi, at the apical pole of the cells. In the liver, most hepatocytes were weakly stained, whereas a few cells showed a relatively strong staining (Figure 3D). Those cells had no specific distribution according to the hepatic lobules. The sinusoidal lining cells, the Kupffer cells, were strongly positive (Figure 3E). Bile ducts were also strongly positive and portal tracts were negative (Figure 3D). In the stomach, the gastric surface epithelium as well as gastric fundic cells (Figures 4A and 4B) and pyloric mucosa (Figure 4C) were strongly stained. Chief cells were strongly stained; the majority of staining was in the basal portion of the mucosa. Parietal cells appeared to be less positive (Figure 4B). The bottom of the glands showed no staining (Figure 4A).
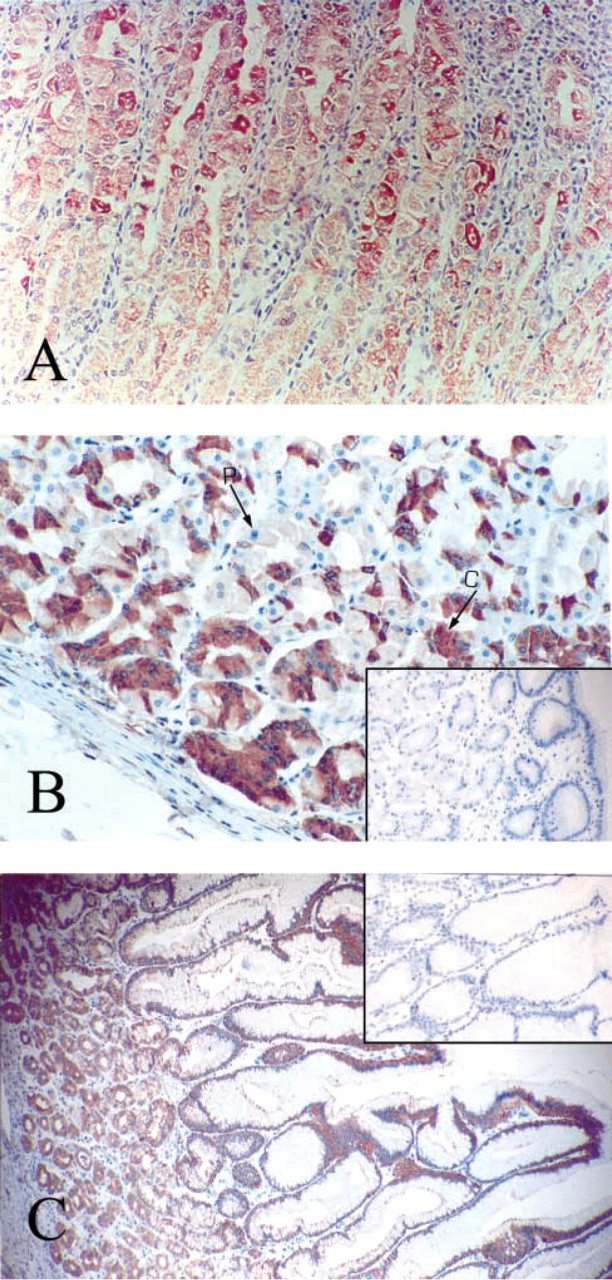
Immunocytochemical staining of human endocrine tissues with MAb 8D7. (
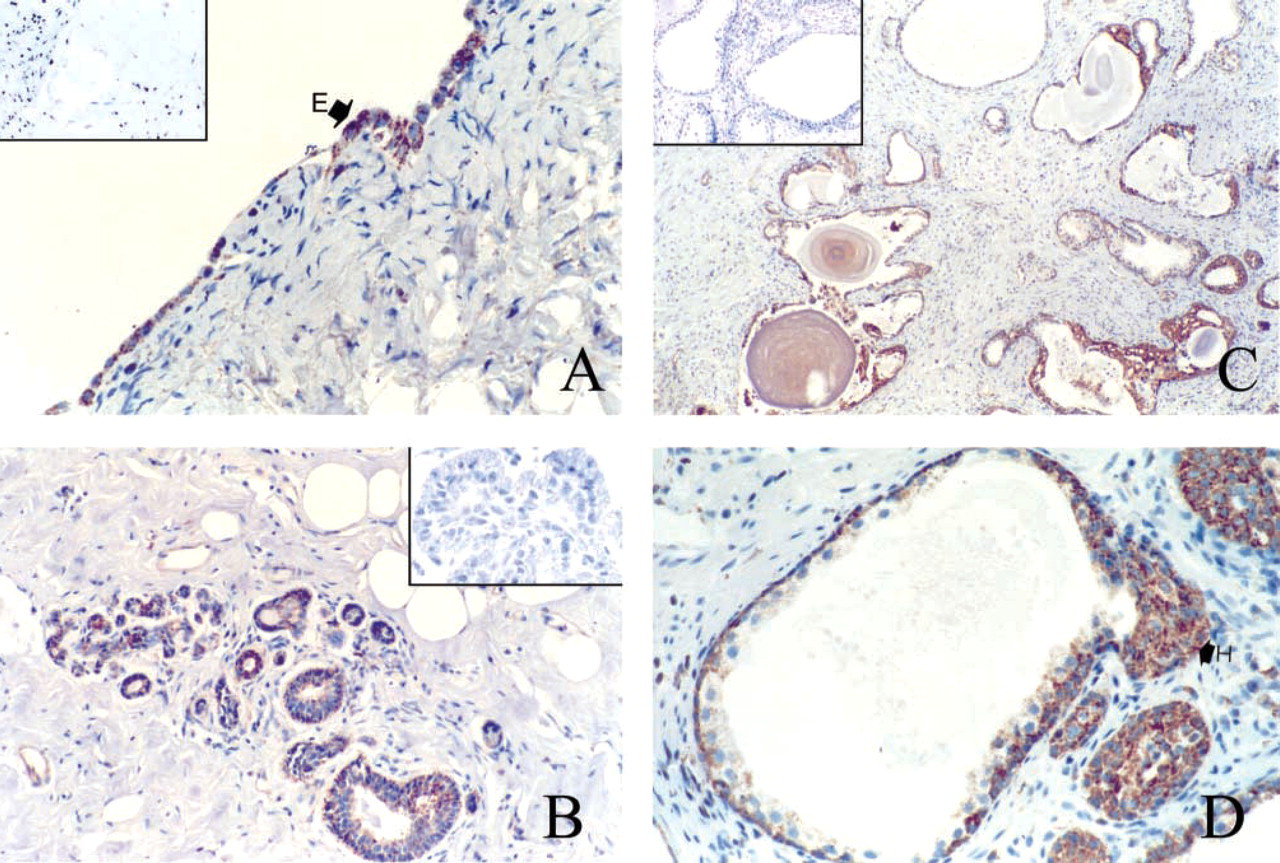
Tissues from the Endocrine System ((Figures 5 and 6). In the thyroid, follicular cells appeared to be weakly or not at all immunoreactive for PBR (Figure 5A). The positive cells were usually located in dystrophic epithelium. Most often, sparse cells, approximately less than 5% of total cell number, were weakly stained with MAb 8D7. Only one of nine samples showed 50% of follicular cells strongly positive for PBR. Colloid appeared to be moderately stained. In the adrenal gland, cells of the adrenal cortex exhibited strong immunostaining for PBR, with a rather homogeneous granular pattern of cytoplasmic and/or peripheral staining around the cytoplasmic membrane, and also in zona glomerula, zona fasciculata, and zona reticularis (Figure 5B). As expected, no staining was observed in the medulla (data not shown). In the pancreas (Figures 5C and 5D), acini, the main exocrine secretory component, showed negative to moderate granular cytoplasmic immunostaining, heterogeneously distributed in the cells of some structures only (Figures 5C). Ducts were rather homogeneously and moderately stained, with luminal staining increasing. Islets, the endocrine component of the pancreas, demonstrated distinct immunostaining patterns from one sample to another. In most pancreas samples, no immunostaining of the islets was observed or, rarely, there were very few sparse endocrine cells expressing PBR. Those samples were highly immunoreactive for insulin, glucagon, and somatostatin (data not shown). In one sample, rare cells or sparse clusters of cells exhibited very strong immunostaining. In this sample, stained granules of various sizes were often concentrated in the cytoplasm adjacent to the nucleus. Some cells appeared totally stained. In Figure 5D, the parenchyma shows strong staining. In the ovary (Figure 6A), which was of postmenopausal status with an atretic corpus albicans, sparse cells of the cortex demonstrated strong granular cytoplasmic staining. The remaining secretory cells in a degenerating corpus albicans were moderately stained. The columnar cells of the surface epithelium were strongly positive for 8D7 immunostaining. In the breast (Figure 6B), normal breast components (ductal and acinar epithelial cells) showed different labeling levels. More often they showed weak granular cytoplasmic immunostaining, with a typical mitochondrial localization, but in some cases the staining was rather strong. The staining was homogeneously distributed in the cells and also in the structures. In many ducts there was a basal or/and a luminal increase in staining. A very similar immunostaining pattern was observed in dystrophic structures. In the prostate (Figures 6C and 6D), ducts and acini of the central zone and the peripheral zone were faintly stained or negative, except for those exhibiting epithelial hyperplasia (regular or atypical). The immunohistochemical data are summarized in Table 1.
Immunocytochemical staining of human endocrine tissues with MAb 8D7. (
Discussion
The distribution of PBR has been widely described in several animal models. The goal of the present study was to characterize further the distribution of PBR in humans, focusing particularly on tissues for which information on PBR expression was rare. To this aim, we used the 8D7 monoclonal anti-PBR antibody that specifically and exclusively recognizes the human PBR (Dussossoy et al. 1996). We first addressed the subcellular localization of PBR. The localization of the PBR in mitochondria has been clearly established for 20 years, as demonstrated using fractionation approaches or confocal microscopy analyses combined with specific markers for this organelle. In mitochondria, auto-radiographic binding studies have suggested that PBR is co-localized with cytochrome oxidase and monoamine oxidase, both of which are known to be expressed on the mitochondrial outer membrane (Anholt et al. 1986). To supply additional probes, we studied PBR expression at the molecular level using electron microscopy. The use of MAb 8D7 in electron microscopic experiments provides direct evidence that the receptor is unambiguously associated with the mitochondrial outer membrane. No other subcellular expression is observed with this antibody. Such a discrepancy with the findings of other studies, which evidenced the expression of PBR in and around the nucleus in breast cancer cell lines and biopsy specimens, may be related to a difference in the C-terminal epitope of PBR (one point mutation in the MDA MB231 cells was reported; Hardwick et al. 1999) or to a difference in signal peptide, chaperones, or partner proteins involved in the regulation of PBR subcellular localization. This may be related to pathological conditions, and further studies are warranted to address this issue.
Summary: distribution of PBR immunoreactivity a
aLabeling index indicates no staining (0), weak (1), moderate (2), and strong (3) staining.
We then performed a broad analysis of PBR in several human tissues using MAb 8D7. We analyzed PBR expression in different respiratory, digestive, and endocrine tissues. In this study, we demonstrated that PBR is widely distributed in almost all normal tissues analyzed (Table 1). In those tissues, PBR expression was generally moderate and was rather homogeneously distributed, although some sparse cells or clusters of cells appeared to be intensely stained. Very often the staining in normal tissues was located at one side of the cells. This pattern of distribution of the immunostaining in differentiated cells of many glandular epithelia (especially those with brush borders and microvilli), as observed in the small intestine, colon, and stomach, appears to fit with the previously described implication of PBR in differentiation mechanisms. An upregulation of PBR expression in differentiated cells versus undifferentiated ones was previously reported in different cell lines (leukemic cells; Ishiguro et al. 1987; Canat et al. 1993b; Taketani et al. 1994), melanoma cells (Landau et al. 1998), and skin (Stoebner et al. 1999). Further supporting a role for PBR in the differentiation process, PBR ligands were shown to induce cell differentiation (Nakajima et al. 1995).
In addition, previous data have shown that PBR is highly expressed in tissues involved in steroid synthesis, and its level of expression in normal tissues is correlated with the amount of mitochondrial materials in the cell. Most likely, the sparse strongly stained cells found in otherwise weakly stained normal tissues represent cells involved in considerable synthesis, which implies large amounts of mitochondrial materials. These data are illustrated by the intense staining of some endocrine islets in weakly stained pancreatic tissue. We did not find any staining in the glandular luminal products except in the thyroid, where colloid often exhibited a moderate staining. Colloid is a storage form of thyroid hormones, which undergo continuous resorption and transport through cytoplasmic pseudopodia from follicular cells. Therefore, the staining of colloid, together with the rare positively stained follicular cells, may correlate with the transport mediating function of PBR, as already shown for cholesterol (Bernassau et al. 1993; Li and Papadopoulos 1998), porphyrins (Taketani et al. 1995), and anions (Basile et al. 1988; Kinnally et al. 1993).
Taken together, our data provide original findings on PBR expression in humans under normal conditions. These data, which may be further extended to pathological states, may be the basis for a better understanding of PBR function.
Footnotes
Acknowledgements
We wish to thank Nadine Lequeux, MicHÈle Radal, and Sylvie Roques, who provided expert technical assistance.