
Abstract
Sphingolipids serve as structural elements of cells and as lipid second messengers. They regulate cellular homeostasis, mitogenesis, and apoptosis. Sphingolipid signaling may also be important in various pathophysiologies such as vascular injury, inflammation, and cancer. Serine palmitoyltransferase (SPT) catalyzes the condensation of serine with palmitoyl-CoA, the first, rate-limiting step in de novo sphingolipid biosynthesis. This integral microsomal membrane protein consists of at least two subunits, SPT1 and SPT2. In this study we analyzed the expression of SPT1 and SPT2 in normal human tissues. Strong SPT1 and SPT2 expression was observed in pyramidal neurons in the brain, in colon epithelium, and in mucosal macrophages. However, SPT2 expression was more prominent than SPT1 in the colon mucosal macrophages, the adrenomedullary chromaffin cells and endothelium, and in the uterine endothelium. SPT2 was localized in both nuclei and cytoplasm of the adrenomedullary chromaffin cells, whereas SPT1 was primarily cytoplasmic. These observations link enhanced SPT expression to proliferating cells, such as the lung, stomach, intestinal epithelium, and renal proximal tubular epithelium, and to potentially activated cells such as neurons, chromaffin cells, and mucosal macrophages. A baseline expression of SPT, established by this study, may serve as a measure for aberrant expression in various disease states.
M
The backbone of various sphingolipids is generated from the long-chain bases sphinganine, sphingosine and, in yeast, phytosphinganine. The first unique and committed reaction to long-chain base synthesis involves the condensation of L-serine with a fatty acid acyl-CoA to generate 3-ketodihydrosphingosine by the enzyme serine palmitoyltransferase [SPT; palmitoyl-CoA; L-serine C-palmitoyltransferase (decarboxylating)] (Merrill 1983). An integral microsomal membrane protein (Mandon et al. 1992), SPT is composed of at least two subunits, SPT1 and SPT2. The catalytic subunit of SPT is believed to be SPT2, whereas the regulatory activity is believed to be the SPT1 subunit. In yeast, both LCB1 and LCB2 subunits are required for LCB activity (Nagiec et al. 1994) and a third component, Tsc3p, is essential for optimal LCB function (Gable et al. 2000).
Recently, SPT expression has been closely linked to pathophysiological conditions. Procedures such as angioplasty result in vascular injury and, in response to injury, a cascade of events collectively known as restenosis is initiated. An increase in SPT1 and SPT2 expression has been reported in proliferating vascular smooth muscle cells and fibroblasts in balloon-injured rat carotid arteries (Uhlinger et al. 2001). Upregulation of SPT subunits has also been observed in proliferating fibroblasts, transformed cell lines, and in various human tumors (unpublished observations). The association of increased SPT expression with injury makes it a potential therapeutic target.
Because SPT activity is altered by a change in the physiological state of the cell, it is imperative to determine the basal levels of this enzyme in normal tissues. The distribution of the SPT1 and SPT2 subunits may serve as a potential marker of cell activity in which high levels of the enzyme may reflect increased activity (e.g., neuronal transmission, exocytosis) or cell proliferation. The SPT1 and SPT2 levels determined in normal tissues and cell types might then be used to analyze cell types in abnormal states such as cancers, inflammation, and vascular injury. Therefore, in this investigation we examined the distribution of SPT1 and SPT2 in normal human tissues using immunohistochemistry (IHC).
Materials and Methods
Antibody Production
Rabbit polyclonal antibodies to the SPT subunits were generated using antigenic peptide sequences predicted by the algorithm of Hopp/Woods. The peptides utilized for antibody production against the human SPT1 subunit (GenBank protein accession number CAA69941) were KLQERSDLT-VKEKEEC, corresponding to residues 45–59, and KEQE-IEDQKNPRKARC, corresponding to residues 222-236. The peptides used as antigens for the human SPT2 subunit (GenBank protein accession number CAA69942) were CGK-YSRHRLVPLLDRPF, corresponding to residues 538-552, and CGDRPFDETTYEETED, corresponding to residues 549-561. A cysteine and glycine were added to the amino terminus of these peptides to allow KLH conjugation and decreased steric hindrance for the coupling. Rabbit polyclonal antibodies were raised against both peptides separately for each SPT subunit. The resulting immune sera were pooled and the mixed polyclonal antisera used as the source of antibody against the specific SPT subunit.
Western Analysis
The specificity of the rabbit anti-SPT1 or rabbit anti-SPT2 polyclonal antibody was evaluated by immunoblotting analysis. Microsomal membranes from HEK 293 cells stably transfected with both SPT1 and SPT2 were prepared as described by Williams et al. (1984). Fifteen micrograms of microsomal membrane protein was fractionated in each of the six lanes of an SDS-polyacrylamide gel. After transfer to a nitrocellulose membrane, the membrane was probed with 2 μg/ml dilution of SPT1- or SPT2-specific polyclonal serum described above. The antigen specificity of the polyclonal serum was determined by preincubating with the serum with 20 μg/ml antigenic peptides overnight at 4C before probing the immunoblot. Bound antibody was detected with an alkaline phosphatase-conjugated goat anti-rabbit IgG (Santa Cruz Biotechnologies; Santa Cruz, CA).
Immunohistochemistry
Commercial human normal and tumor checkerboard tissue slides (DAKO, Carpinteria, CA; Biomeda, Foster City, CA) were deparaffinized, hydrated, and processed for routine IHC as previously described (D'Andrea et al. 1998). Briefly, slides were microwaved in Target (DAKO), cooled, placed in PBS, pH 7.4, then placed in 3.0% H2O2. Slides were processed through an avidin-biotin blocking system according to the manufacturer's instructions (Vector Labs; Burlin-game, CA) and then placed in PBS. All reagent incubations and washes were performed at room temperature. Normal blocking serum (Vector Labs) was placed on all slides for 10 min. After brief rinsing in PBS, primary antibodies (Table 1) were placed on slides for 30 min. The slides were washed and biotinylated secondary antibodies, goat anti-rabbit (goat anti-rabbit) or horse anti-mouse (monoclonal antibodies), were placed on the tissue sections for 30 min (Vector Labs). After rinsing in PBS, the avidin-horseradish peroxidase-biotin complex reagent (ABC; Vector Labs) was added for 30 min. Slides were washed and treated with the chromogen 3,3'-diaminobenzidine (DAB; Biomeda), rinsed in dH2O, and counterstained with hematoxylin. Monoclonal antibodies to vimentin (Table 1) were used to demonstrate tissue antigenicity and reagent quality. Negative controls included replacement of the primary antibody with the same species IgG isotype nonimmune serum (Table 1). The specificity of the primary SPT antibodies was again determined by preincubating the polyclonal sera overnight with the anti-genic peptides as described above.
Double Immunohistochemical Labeling
Protocols for simultaneous double immunohistochemical labeling (IHC:IHC) have been previously published (D'Andrea et al. 1999) and are similar to those cited for single IHC labeling except that the slides were not processed for counter-staining after the second chromogen step of the first antigen detection protocol. Instead, the slides were placed in PBS and the second antigen was detected by an alkaline phosphatase-Fast Red system. The primary antibody was placed on the slides for 30 min at RT. After brief washing, the secondary biotinylated antibody was added for 30 min at RT. The slides were first washed in PBS and then the streptavidin-alkaline phosphatase reagent was placed on the slides for 30 min at RT. After washing, the Fast Red chromogen (DAKO) was placed on the slides twice for 5 min. Then the slides were processed for routine counterstaining in hematoxylin, washed, and coverslipped in a water-based mounting medium (DAKO) for viewing under a BX-50 Olympus light microscope.
Multiple controls were performed to ensure correct interpretation of the labeling on the slides. The primary antibodies were substituted with the proper species isotype to control for the detection systems. On another set of controls, the first primary was omitted and the second primary antibody was processed, and vice versa.
Results
Specificity of SPT1 and SPT2 Antibodies
Rabbit polyclonal antibodies specific for the two human SPT subunits were generated as described in Materials and Methods. The specificity of the antibodies is demonstrated in the immunoblot shown in Figure 1a. Microsomal membrane fractions obtained from SPT stably transfected HEK cells were resolved by SDS-PAGE and the Western blot was probed with either preimmune serum or the SPT-specific polyclonal antibodies in the presence or absence of competing peptides. Single immunoreactive bands of the expected molecular weights (Weiss and Stoffel 1997) were observed, specifically Mr ∼55 kD for SPT1 (Figure 1a, Lane 3) and Mr ∼65 kD for SPT2 (Figure 1a, Lane 6). Preincubating the polyclonal antibodies with the antigenic peptides before probing the immunoblot competitively inhibited detection of SPT1 (Figure 1a, Lane 4) and SPT2 (Figure 1a, Lane 7). No nonspecific binding was observed with the preimmune serum (Figure 1a, Lanes 2 and 5).
Primary antibodies
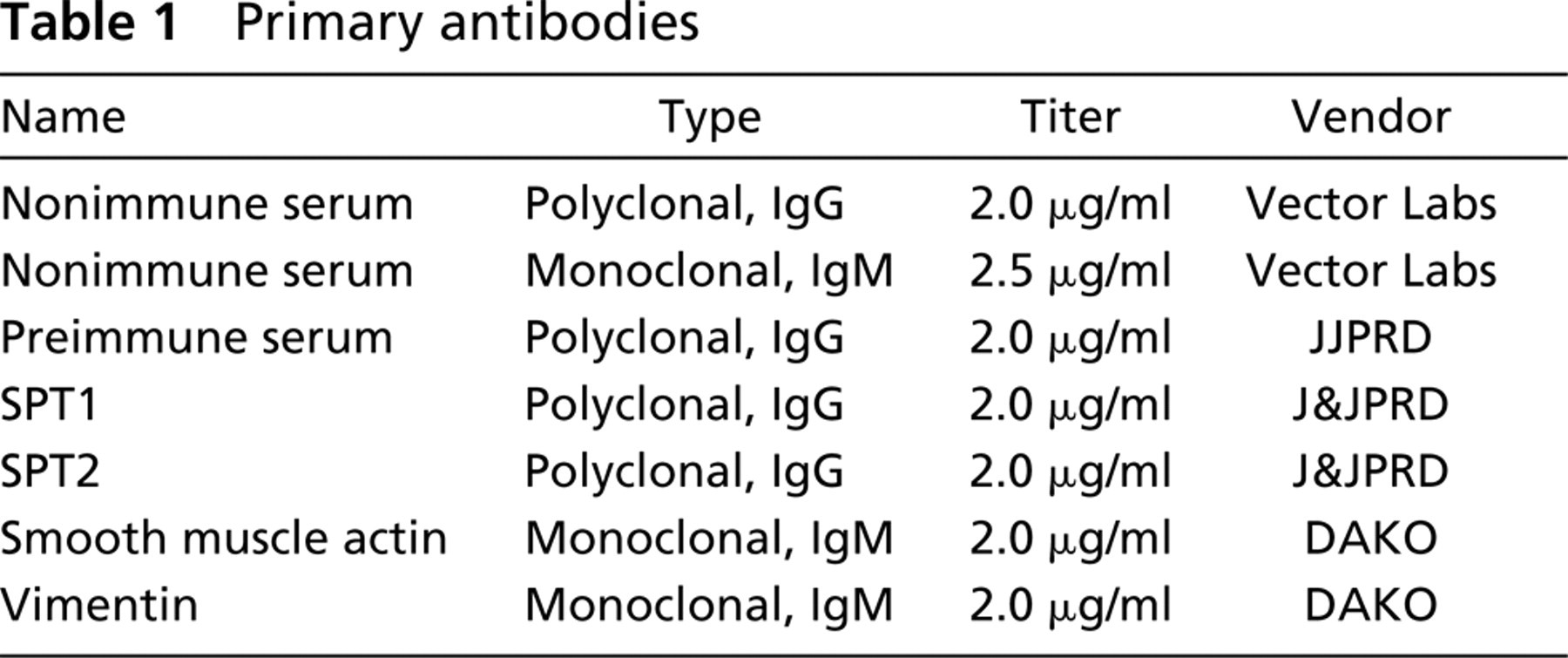
The specificity of polyclonal antibodies in IHC was also determined (Figures 1b-1e). Preincubation of the polyclonal antisera with a ten fold excess of antigenic peptides before probing the slides competitively inhibited detection of SPT1 and SPT2 (Figures 1b and 1d).
Tissue Distribution of SPT1 and SPT2
SPT1 and SPT2 protein expression in normal human tissues was analyzed using IHC. Formalin-fixed, paraffin-embedded tissues were used in a multi-tissue format to eliminate potential staining artifacts such as slide-to-slide and run-to-run variability. Table 1 lists the positive and negative controls in addition to the experimental antibodies. Positive labeling was defined by the strength of brown staining and was scored according to the following criteria: no immunoreactivity was scored as negative (N); light-brown immunoreactivity was scored as weak (W); brown immunoreactivity was scored as moderate (M); and dark-brown immunoreactivity was scored as strong (S). The negative controls did not produce observable labeling.
The distribution of SPT1 and SPT2 in human tissues is presented in Table 2. In general, the vascular endothelium and smooth muscle cells were moderately immunopositive for SPT1 and SPT2. Except for the ovarian epithelium, the epithelial cells in all tested tissues were moderate to strongly immunopositive for SPT1 and SPT2. In addition, mucosal macrophages from the colon, lung, and stomach were strongly immunopositive for SPT1 and SPT2. In the spleen, the macrophages and the polymorphonuclear cells (PMNs) stained positive for both SPT1 and SPT2, but no reactivity was observed in the lymphocytes. The colon, lung, prostate, stomach, thyroid, uterus, and vascular tissues were moderate to strongly immunopositive for SPT1 and SPT2. However, SPT1 and SPT2 were either weakly present or completely undetectable in the skin and heart tissues using the above described protocol. Figures 2–6 show some of the human tissues tested for SPT1 and SPT2 expression by IHC.
Normal brains were immunolabeled with preimmune serum (Figure 2a), SPT1 (Figure 2b)-, and SPT2 (Figure 2c)-specific antibodies. In the cerebral cortex, the pyramidal neurons (arrowheads) showed positive immunoreactivity for SPT1 and SPT2. Both SPT1 and SPT2 were localized in the neuronal cytoplasm and the expression levels of both subunits appeared similar. Purkinje cells in the human cerebellum were moderately immunopositive for both SPT1 and SPT2 (data not shown). In contrast, SPT1 and SPT2 were not detectable in other neuronal cell types, such as astro-cytes, microglia, and oligodendritic cells.
Competition assay to show specificity of the anti-SPT1 and anti-SPT2 polyclonal antibodies using immunoblotting analysis (
Immunolocalization of SPT1 and SPT2 in normal human tissues a
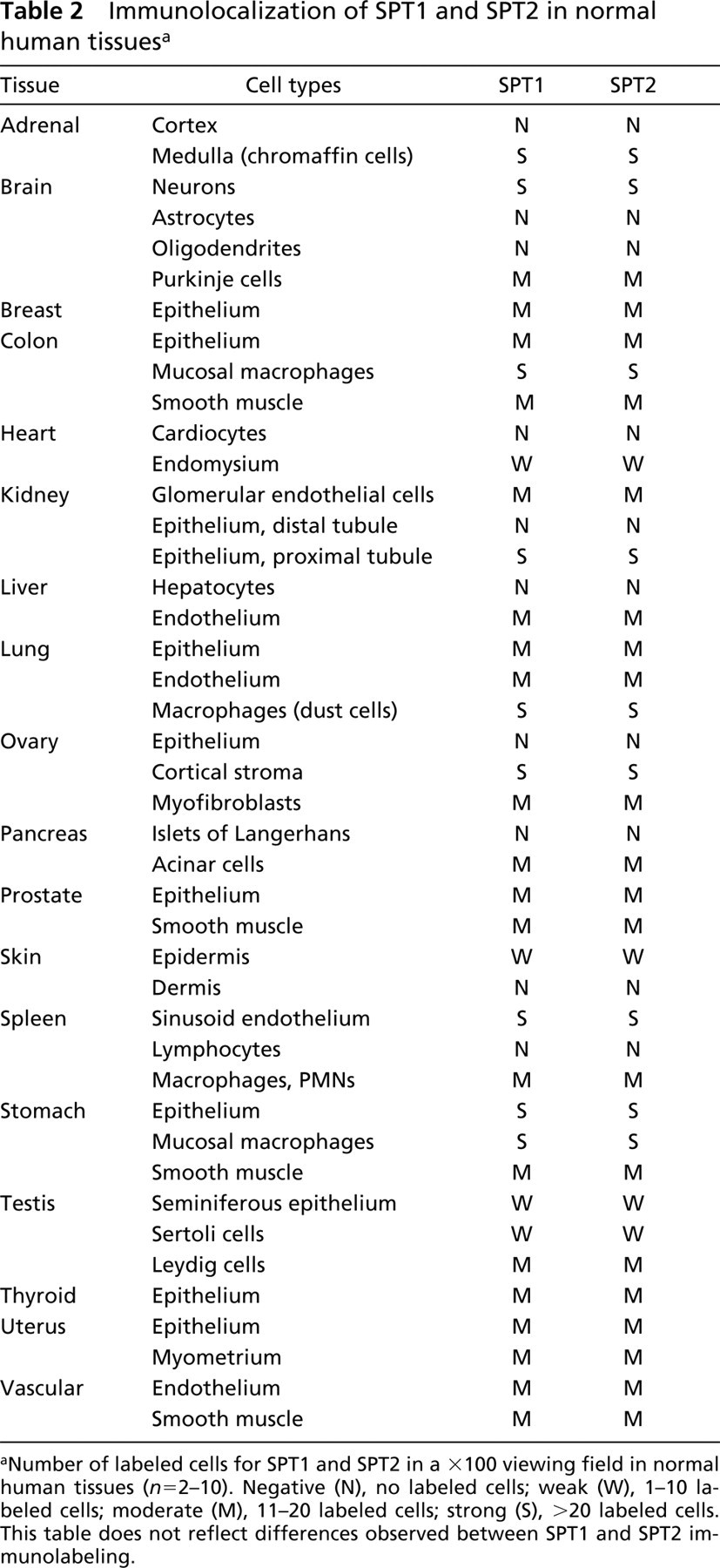
aNumber of labeled cells for SPT1 and SPT2 in a X100 viewing field in normal human tissues (n = 2-10). Negative (N), no labeled cells; weak (W), 1-10 labeled cells; moderate (M), 11-20 labeled cells; strong (S), >20 labeled cells. This table does not reflect differences observed between SPT1 and SPT2 immunolabeling.
In the human colon (Figure 3), epithelial cells (small arrowheads) and macrophages (large arrowheads) stained positive for SPT1 (Figure 3b) and SPT2 (Figure 3c). As in the neurons, expression of both SPT1 and SPT2 was mainly cytoplasmic. Compared to the moderate expression of SPT2 in the epithelial cells, the mucosal macrophages exhibited a much stronger immunoreactivity to SPT2. No immunoreactivity was observed in any cell type on staining with the preimmune serum (Figure 3a). The high expression of SPT in mucosal macrophages in the colon (Figures 3b and 3c) and stomach and dust cells (alveolar macrophages) (Table 2) may be due to the fact that these macro-phages are associated with tissues that are prone to environmental exposure and may therefore have been activated.
Figure 4 shows normal human adrenal glands immunolabeled with either preimmune serum (Figure 4a) or SPT1 (Figure 4b)- or SPT2 (Figure 4c)-specific antibody. Chromaffin cells of the adrenal medulla (large arrowheads), vascular smooth muscle cells (arrows), and endothelium (small arrowheads) showed strong positive cytoplasmic immunoreactivity for SPT1 and SPT2. SPT1 and SPT2 expression was undetectable in the adrenal cortex. SPT2 expression in the endothe-lium and the chromaffin cells appeared higher than SPT1. In addition to the cytoplasm, SPT2 expression could be clearly observed in the chromaffin cell nuclei.
Immunolabeling of normal human kidneys with preimmune serum (Figure 5a), SPT1 (Figure 5b)- or SPT2 (Figure 5c)-specific antibody showed SPT1 and SPT2 in the proximal tubules (arrowheads). Endothelial cells of the glomerulus were also SPT1- and SPT2-immunopositive (data not presented). Interestingly, the expression of SPT1 and SPT2 was again different in the proximal tubules. SPT1 expression was diffuse in the cytoplasm, whereas SPT2 immunostaining appeared more punctate and overall weaker than SPT1. No immunoreactivity was observed with the preimmune serum. Thus far, SPT activity has been localized to the cytosolic side of the endoplasmic reticulum (Mandon et al. 1992). Note the proximity of SPT2 expression to the nucleus. Because the endoplasmic reticulum is closely associated with the nucleus, the punctate appearance of SPT2 in the renal proximal tubule epithelium suggests its association with the endoplasmic reticulum.
Several normal human uteri were similarly immunolabeled with either preimmune serum (Figure 6a) or the SPT1 (Figure 6b)- or SPT2 (Figure 6c)-specific antibodies. Uterine smooth muscle cells (large arrowheads) demonstrated similar positive immunoreactivity for SPT1 and SPT2. However, the expression of SPT2 was higher than SPT1 in the endothelial cells (small arrowheads). No immunoreactivity was observed with the preimmune serum.
Our study indicated that SPT1 and SPT2 expression was particularly strong in cells, such as the adrenal chromaffin cells, that secrete epinephrine and norepinephrine on autonomic nervous stimulation and in neurons. Because it also appeared that SPT1-and SPT2-positive labeling was observed in proliferating cell types, such as the epithelial layers in the stomach, lungs (data not shown), renal proximal tubules, and colonic lumen, double IHC labeling of the human large intestines was performed. Antibodies to PCNA (in red), a marker of cell proliferation (D'Andrea et al. 1994), were used in combination with either the SPT1 (Figure 7a)- or the SPT2 (Figure 7b)-specific antibody (in brown). Large arrowheads show the co-localization of red- and brown-labeled cells, indicating that both SPT1 and SPT2 are expressed in proliferating epithelial cells.
Discussion
Our current studies characterize the distribution of the SPT subunits SPT1 and SPT2 in normal human tissues. The differences we observed in expression of SPT1 and SPT2 indicate that the localization and expression levels of SPT may be linked to the physiological state of the cell. Proliferating cells and cells that may potentially be activated, such as mucosal macro-phages in colon or chromaffin cells in the adrenal medulla, expressed higher levels of SPT. The presence of moderate to high levels of SPT1 and SPT2 in vascular tissues suggests a role for SPT in regulating signaling pathways involving messengers like S-1-P.
The differences between SPT1 and SPT2 expression in the same cell type within the same tissue suggests that there must be specific and possibly independent functions of each subunit in enabling SPT activity. How these two subunits interact and coordinate SPT activity is still unknown. Unlike yeast, overexpression of murine SPT2 alone in human HEK293 cells results in a corresponding increase in SPT activity, whereas SPT1 alone does not increase SPT activity (Weiss and Stoffel 1997). Whether an increase in SPT2 alone is sufficient for upregulation of SPT activity remains to be seen. In addition to SPT1 and SPT2, human SPT may also have additional components like the Tsc3p protein in yeast (Gable et al. 2000). Moreover, the localization of SPT2 in the nuclei suggests that SPT2 associates with another nuclear protein(s) or is modified and transported to the nucleus. Therefore, analysis of the difference in dynamics of SPT1 and SPT2 expression will help in elucidating SPT activity.
Enzymes that regulate sphingolipid metabolism are critical in maintaining cellular homeostasis, and a disruption of their activity can lead to disease. Inhibition of ceramide synthase by fumonisin mycotoxins contaminating animal feeds results in equine leukoen-cephalomalacia and porcine pulmonary edema (reviewed in Marasas 2001). Local delivery of C6-ceramide in rabbit carotid arteries after balloon angioplasty reduced neointimal hyperplasia by inhibiting extracellular signal-related kinase ERK and phosphorylation of protein kinase B (PKB/Akt) (Charles et al. 2000). Signaling pathways involving ERK, PKB/Akt, and the angiotensin II receptor are involved in vascular smooth muscle migration and growth. Activation of angiotensin II receptor induces de novo sphingolipid biosynthesis, leading to programmed cell death (Lehtonen Jukka et al. 1999). Lowering S-1-P production in TNF-α-induced endothelial cells by HDL reduces the expression of adhesion proteins and consequently increases protection against artherosclerosis (Xia et al. 1999). Therefore, enzymes regulating sphingolipid metabolism are key factors in controlling sphingolipid mediated regulation of cellular phenomena.
Because SPT is the key regulatory enzyme in de novo sphingolipid biosynthesis, it is expected that an alteration in SPT activity would affect sphingolipid-mediated regulation of cell function. In yeast, SPT has been implicated in heat and hyperosmolar stress responses (Buede et al. 1991; Patton et al. 1992; Jenkins et al. 1997). Cultured human keratinocytes, when UV-irradiated, upregulate SPT activity and show a corresponding increase in SPT2 mRNA and protein levels (Farrell et al. 1998). SPT activity is increased during apoptosis and governs de novo ceramide synthesis in cells treated with the chemotherapeutic agent etoposide (Perry et al. 2000). Inhibition of SPT activity by myriocin reverses the apoptotic and antiproliferative effects of a ceramide synthase inhibitor, fumonisin, in pig kidney cells LLCK-1 (Riley et al. 1999).
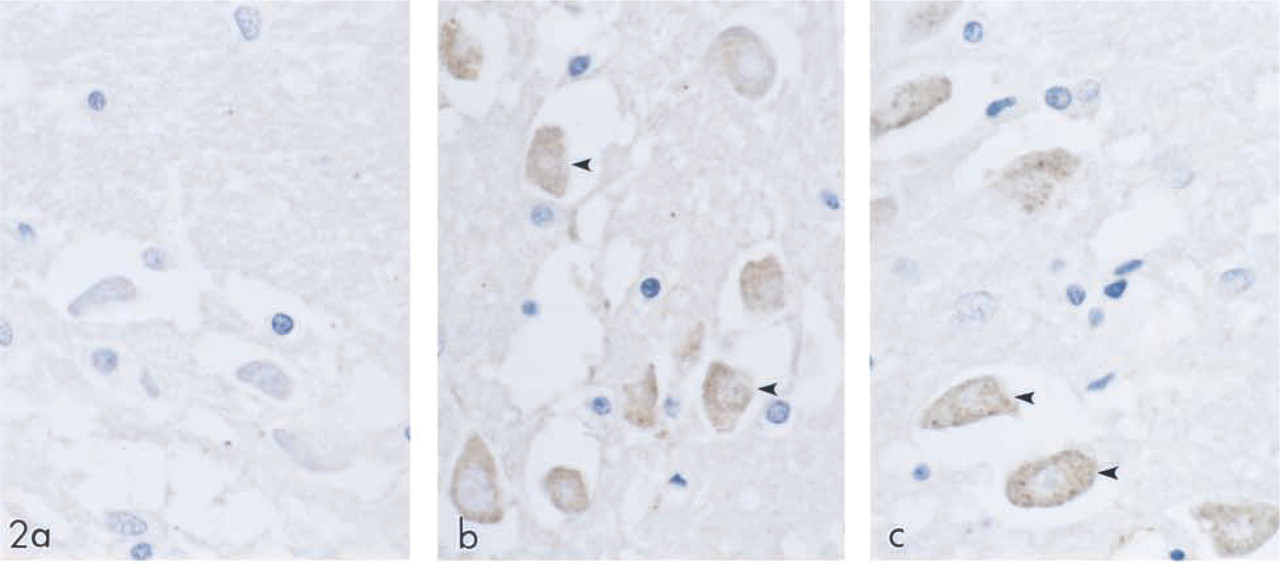
SPT expression in human brain tissue. Normal human brain was immunolabeled with preimmune serum (a), SPT1 (
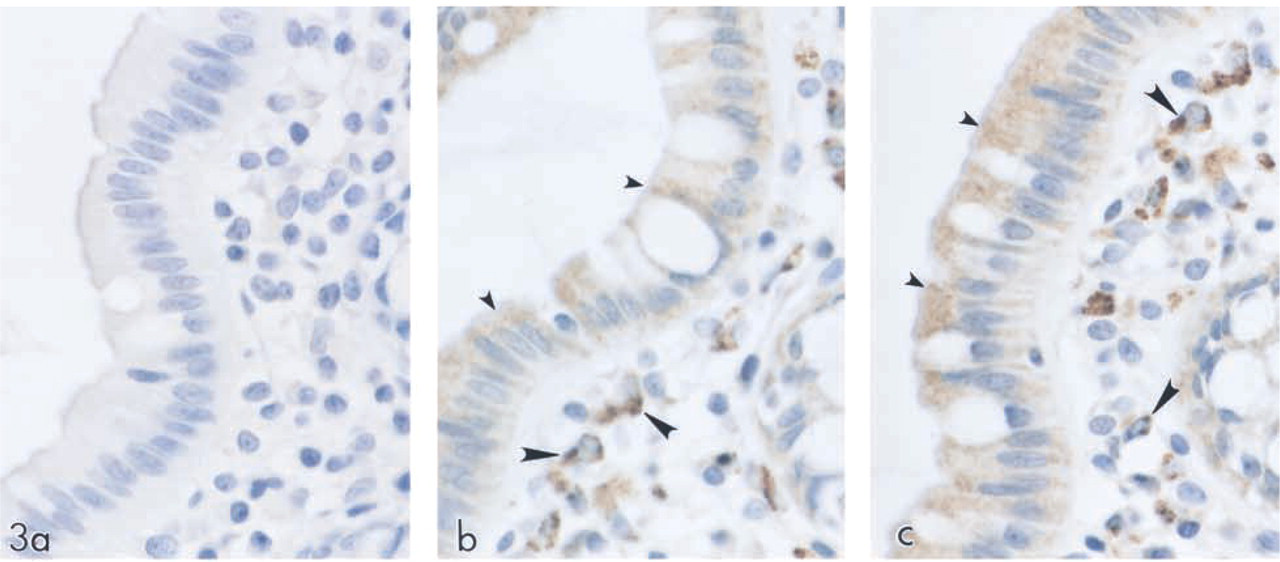
SPT expression in human colon. Normal human large intestine was immunolabeled with preimmune serum (a), SPT1 (
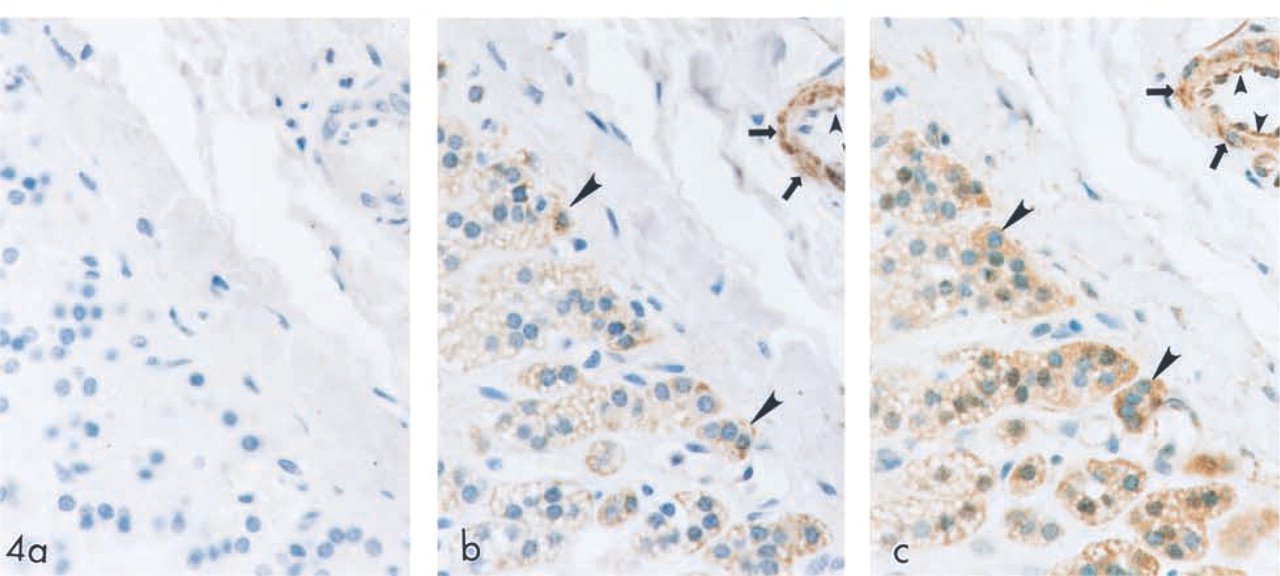
SPT expression in human adrenal tissue. Normal human adrenal gland was immunolabeled with preimmune serum (a), SPT1 (
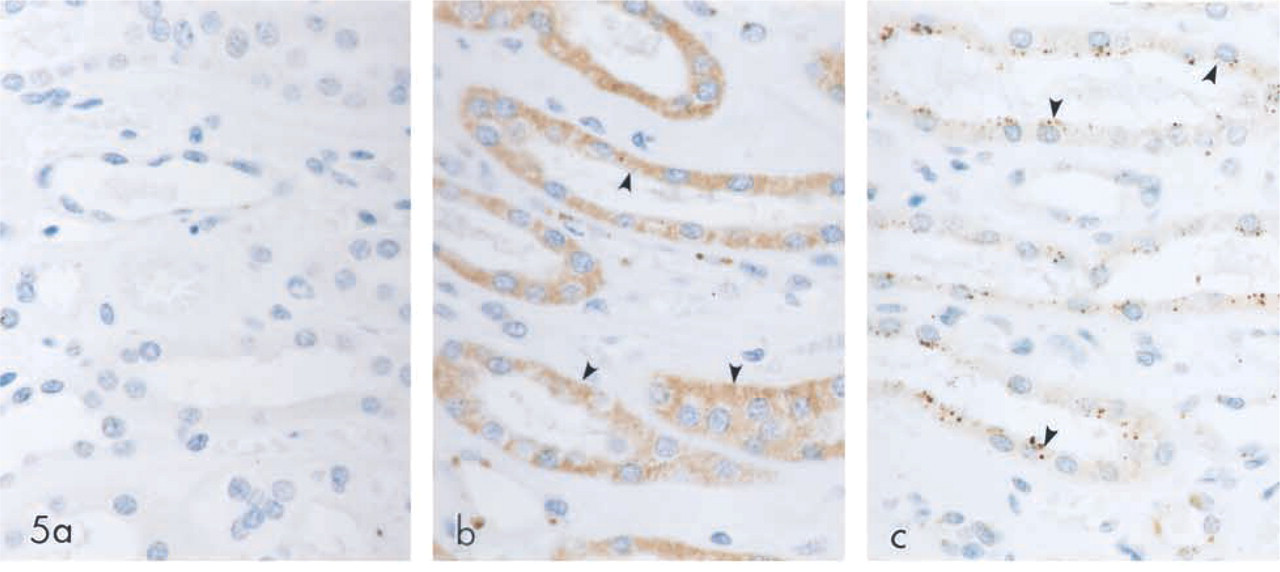
SPT expression in human kidney tissue. Normal human kidney was immunolabeled with preimmune serum (a), SPT1 (
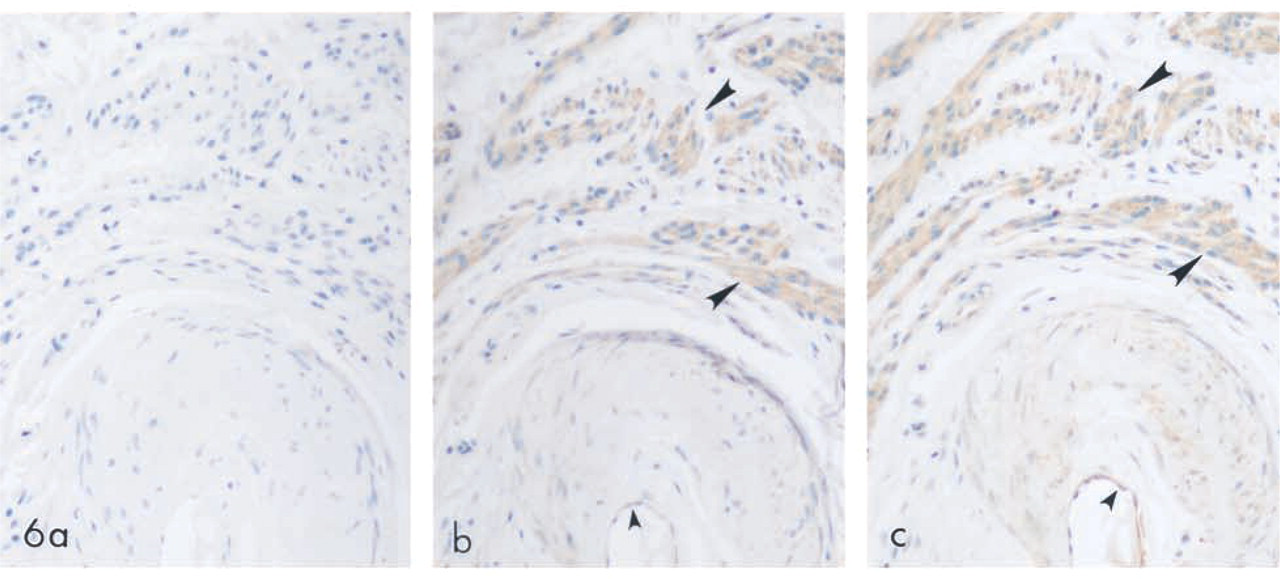
SPT expression in human uterus. Normal human uterus was immunolabeled with preimmune serum (a), SPT1 (
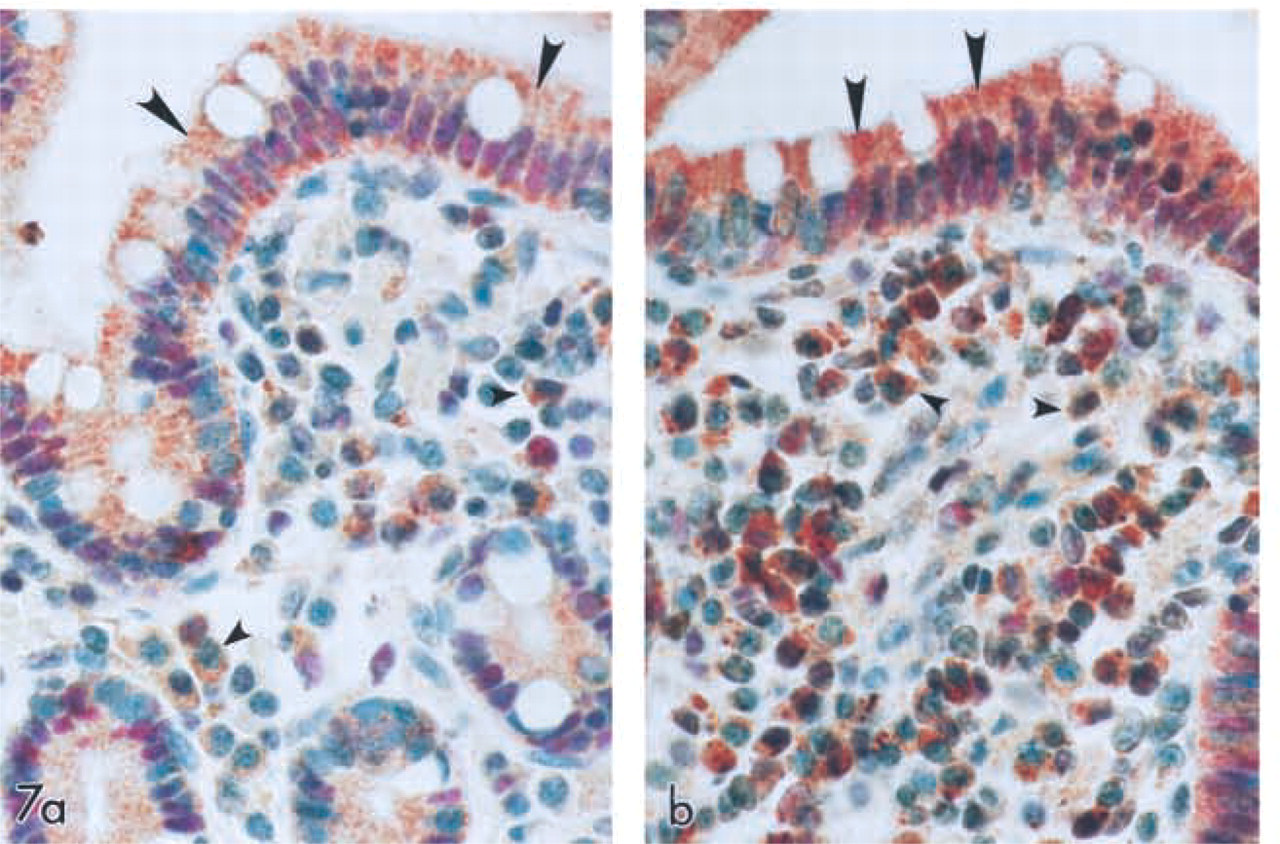
Co-expression of SPT1 and SPT2 with PCNA in the human colon. Double IHC labeling of human large intestine using antibodies to PCNA (red) and SPT1 (a) and SPT2 (brown) (
A knowledge of SPT expression in normal cells can be used to measure abnormal cellular activity in proliferative disorders such as cancers. Both the absolute level of expression of SPT and the localization of enzyme activity may be indicative of an alteration in cell physiology. The increase in SPT activity observed in pathophysiological conditions, such as vascular hyperplasia (Uhlinger et al. 2001), wound healing, and tumors (unpublished observations), suggests therapeutic potential for SPT. Inhibiting or lowering SPT activity in these conditions might affect the symptoms associated with the conditions. In porcine epithelial kidney cells, LLC-PK1, fumonisin-induced cytotoxity and antiproliferative effects were reduced on treating the cells with SPT1-specific inhibitors such as myriocin. In the same study, IP administration of myriocin to BALB/C mice reduced free sphingosine accumulation in the kidney by 60%, with no apparent clinical side effects (Riley et al. 1999). Therefore, SPT inhibitors such as myriocin may have important therapeutic potential in treatment of proliferative disorders such as cancer and may affect pathophysiologies associated with conditions such as inflammation and vascular injury.
The immunolocalization of SPT1 and SPT2 presented here is the first direct comparison of SPT1 and SPT2 expression in normal human tissues and is a critical first step towards elucidating the complexity of SPT activity in the cell. Understanding the role of these components in SPT activity is imperative in determining the regulation of the many critical sphingolipid-mediated cellular functions and responses in various disease states.
Footnotes
Acknowledgements
We express our thanks for the excellent histological and immunohistochemical expertise of Patti A. Reiser, BS, MT, HT (ASCP), Norah A. Gumula, HT (ASCP), Zabrina Thomson, MS, and Brenda M. Hertzog, BS, MT (ASCP) of the Morphometrics Department.