
Abstract
The Hedgehog (Hh) family of secreted proteins includes intercellular signaling molecules that specify cell fate and patterning during the development of many tissues. In this study we show that the different components of the Hh signaling pathway are expressed in human thymus. The three mammalian Hh proteins, Sonic (Shh), Indian (Ihh), and Desert (Dhh) hedgehog, are produced by thymic epithelial cells. Shh-expressing epithelial cells are restricted to the thymic subcapsula and medulla, whereas Ihh- and Dhh-producing epithelial cells are distributed throughout the thymus. The requisite Hh receptors, Patched 1(Ptc1) and Smoothened (Smo), and the Gli transcription factors are expressed by thymocytes and also by epithelial cells. Ptc1 is expressed in most thymocyte subsets, whereas Smo expression is mainly associated with immature thymocytes. The isoform of the Ptc receptor, Ptc2, is expressed only by intrathymic progenitor cells and epithelial cells. Other Hh-binding proteins with modulating functions, such as Hedgehog-interacting protein (Hip) and growth arrest-specific gene-1 (Gas-1), are also expressed in human thymus. Our study shows that the intrathymic expression pattern of the Hh signaling pathway components is complex and suggests that Hh proteins may regulate human thymocyte differentiation from the earliest developmental stages, as well as thymic epithelial cell function.
T
The ancient Hedgehog (Hh) family includes secreted proteins that regulate cell fate and patterning during the development of many organs. Sonic (Shh), Indian (Ihh), and Desert hedgehog (Dhh) are the mammalian Hh proteins (Ingham and McMahon 2001). They act as morphogens establishing concentration gradients, and this mechanism of cell communication is essential for organogenesis from flies to mammals. Nevertheless, growing evidence reinforces the idea that Hh proteins can also be considered as differentiating, mitogenic, survival, or apoptotic factors (Ingham and McMahon 2001; Bale 2002; Mullor et al. 2002). The developmental functions of Shh are very diverse. It orchestrates left-right asymmetry, dorsoventral patterning of the central nervous system, limb morphogenesis, and also participates in the development of many organs. Ihh and Dhh functions are more restricted. Ihh plays key functions in the development of bone and cartilage, and Dhh is involved in the development of testes and external genitalia, as well as in the formation of peripheral nerve ensheathment (Ingham and McMahon 2001; Bale 2002).
All Hh proteins share an apparently exclusive signaling pathway, in which two Patched (Ptc) isoforms, Ptc1 and Ptc2, and Smoothened (Smo) are the surface receptors. In the absence of Hh ligands, Ptc represses Smo, while the binding of Hh to Ptc releases Smo to signal into the cell. In the absence of Ptc inhibitory function Smo is permanently signaling (Ingham and McMahon 2001). The mechanism underlying the Ptc-Smo relationship remains enigmatic, although recent evidence supports the idea that Ptc does not inhibit Smo activity by direct interaction but rather through the regulation of vesicle trafficking and cytoplasmic transport of small modulating molecules (Nybakken and Perrimon 2002; Taipale et al. 2002).
The intracellular signaling cascade initiated by Smo culminates in activation of members of the Gli family of zinc finger transcription factors, Gli 1, Gli 2, and Gli 3. The Hh pathway has a dual effect on Gli proteins. It induces the release to the nucleus of Gli activating forms and simultaneously inhibits the production of Gli repressor fragments (Ruiz i Altaba 1999; Koebernick and Pieler 2002; Mullor et al. 2002). The specific functions of the different Gli proteins are still unclear, although all the activities demonstrated for Gli1 are positive, whereas Gli2 and Gli3 mainly function as positive and negative regulators of transcription, respectively (Ruiz i Altaba 1999; Aza-Blanc et al. 2000; Wang et al. 2000; Bai and Joyner 2001; Bai et al. 2002; Mullor et al. 2002).
Recently we have demonstrated the presence of components of the Hh cascade in the murine thymus and their involvement in T-cell maturation (Outram et al. 2000). Shh is produced by thymic epithelial cells, Ihh expression is restricted to the blood vessels located in the thymic medulla, and the Hh receptors Ptc and Smo are mainly expressed by immature CD25+ DN thymocytes. The treatment of murine fetal thymus organ cultures (FTOCs) with Shh arrests thymocyte differentiation at the CD25+ DN stage after TCRβ rearrangement and, conversely, the addition of anti-Shh accelerates the progression of DN cells into the DP stage. Shh is also described to regulate the function of peripheral CD4+ T-cells (Lowrey et al. 2002; Stewart et al. 2002) as well as the expansion of primitive human hematopoietic cells (Bhardwaj et al. 2001). In this study we show that the three mammalian Hh proteins, their specific receptors, and other Hh-binding proteins with modulating functions, as well as the Gli family of transcription factors, are also expressed in the human thymus, implying that Hh signaling might regulate the differentiation of human thymocytes.
Materials and Methods
Isolation of Human Thymocyte Subsets
Human thymus samples were obtained from children aged 1 month to 3 years undergoing corrective cardiovascular surgery. Thymus tissues were obtained and used according to the guidelines of the Medical Ethics Commission of the Hospitals La Zarzuela and Madrid-Montepríncipe (Madrid, Spain). Informed consent was provided according to the Helsinki declaration. To isolate thymic CD34+ progenitor cells, thymuses were first dissected free of surrounding connective tissue and then gently disrupted with a Potter homogenizer until completely disaggregated. Cell suspensions were enriched in immature thymocytes by using the sheep red blood cell rosseting technique, as previously described (Varas et al. 2000). The remaining mature T-, B-, NK, myeloid, and dendritic cells were then depleted by treatment with saturating amounts of anti-CD3, anti-CD19, anti-CD56, anti-CD14, and anti-CD11c MAbs (all from Becton Dickinson; San Jose, CA) bound to sheep anti-mouse Ig-coated magnetic beads (Dynal; Oslo, Norway). CD34+ cells were then purified by magnetic sorting using VarioMACs (Myltenyi Biotec; Bergisch Gladbach, Germany) in conjunction with CD34 Multisort Kit (Miltenyi Biotec) following the instructions from the manufacturer. Thymocyte subpopulations defined by CD4/CD8 expression were purified from the whole thymic population using CD4 Multisort Kit and CD8-Microbeads (Miltenyi Biotec). The purity of the enriched subpopulations was always greater than 98%.
Isolation of Human Epithelial Cells
Thymic fragments were cultured floating on Millipore filters (8 μm pore size) (Millipore Ibérica; Madrid, Spain) in RPMI 1640 (Invitrogen; Grand Island, NY) supplemented with 5% FCS (Harlan Sera-Lab; Leicestershire, UK), and 1.35 mM 2-deoxyguanosine (Sigma España; Madrid, Spain). After 7 days, thymic fragments were trypsinized (0.25% trypsin in 0.02% EDTA) (Sigma España) to form a single-cell suspension. Residual thymocytes, NK, B-, myeloid, and dendritic cells were depleted as described above by adding anti-CD34 to the cocktail of purified MAbs.
Cell Lines
Human postnatal thymic epithelial cell lines P1.1A3 and P1.4D6 were kindly provided by Dr. M.L. Toribio (Centro de Biología Molecular “Severo Ochoa”; UAM, Madrid). Cell lines were maintained in RPMI 1640 supplemented with 10% FCS, 2 mM
RT-PCR Analysis
RNA isolation was performed using a Strataprep Total RNA Miniprep Kit (Stratagene Cloning Systems; La Jolla, CA), including a DNase I digestion step, as recommended by the supplier, to avoid genomic DNA contamination. Total cDNA was synthesized with Superscript II RT polymerase (Invitrogen), according to the instructions of the commercial supplier and then used as target in the PCR amplifications. Primers were purchased from Amersham Biosciences (Poole, UK). Amplifications were performed using the following primer sets and annealing conditions: β-ACTIN, forward primer 5′-AGAGATGGCCACGGCTGCTT-3′, reverse primer 5′-ATTTGCGGTGGACGATGGAG-3′ at 61C with a 445-bp; GAS1, forward primer 5′-GAAAGGGAAGGTGCTGACC-3′, reverse primer 5′-CAAGGGCTCAAACTTATCCAA-3′ at 60C with a 311-bp; HIP, forward primer 5′-TGACCCAGACTCACAATGGA-3′, reverse primer 5′-CTCTGCGGATGTTTCTGTCC-3′ at 60C with a 315-bp; SHH, forward primer 5′-CGGAGCGAGGAAGGGAAAG-3′, reverse primer 5′-TTGGGGATAAACTGCTTGTAGGC-3′ at 58C with a 262-bp; IHH, forward primer 5′-CTACGCCCCGCTCACAAAG-3′, reverse primer 5′-GGCAGAGGAGATGGCAGGAG-3′ at 60C with a 376-bp; DHH, forward primer 5′-GTTGTAAGGAGCGGGTGAAC-3′, reverse primer 5′-GCCAGCAACCCATACTTGTT-3′ at 58C with a 184-bp; PTC2, forward primer 5′-CTGGCTTCGTGCTTACTTCC-3′, reverse primer 5′-CGGGTGTGAGGATGTTCTCT-3′ at 58C with a 287-bp product; GLI2, forward primer 5′-ACCAGAATCGCACCCACTCC-3′, reverse primer 5′-GCATCTCCACGCCACTGTCA-3′ at 58C with a 393-bp product; GLI3, forward primer 5′-CACTACCTCAAAGCGGGAAG-3′, reverse primer 5′-TGTTGGACTGTGTGCCATTT-3′ at 58C with a 403-bp product. Primers were designed from sequences available from the GenBank data base (accession numbers: β-ACTIN, BC016045; GAS1, NM 002048.1; HIP, NM 022475.1; SHH, NM000193.2; IHH, XM050846.2; Dhh, NM021044.1; PTC2, NM 000264.1; GLI2, NM5270.2; GLI3, NM000168.2). Primers to detect PTC1, SMO, and GLI1 were previously described (Bhardwaj et al. 2001). All PCR reactions were performed on a Mastercycler gradient machine (Eppendorf; Hamburg, Germany) using AmpliTaqGold DNA polymerase (Applied Biosytems; Foster City, CA) under the following conditions: 3 min at 94C, 40 cycles of 45 sec at 94C, 45 sec of each particular T annealing, 45 sec at 72C, followed by 10 min at 72C. PCR products were resolved on a 2% agarose gel and the measured sizes were as expected.
Histology and Immunofluorescence
Thymus cryosections (5 μm) were air-dried for 2 hr at room temperature and fixed in acetone for 10 min. Nonspecific binding of antibodies and streptavidin was blocked by incubation with diluted donkey serum (Santa Cruz Biotechnology; Santa Cruz, CA) and avidin-biotin (Vector Laboratories; Burlingame, CA). Human thymus sections were incubated with specific goat antibodies against Smo (C-17), Ptc1 (G-19), Ptc2 (N-19), C-terminal Ihh (C-15) (all from Santa Cruz Biotechnology), C-terminal Dhh, and C-terminal Shh (R&D Systems; Minneapolis, MN). Then they were stained with either biotin- or FITC-conjugated multi-adsorbed F(ab')2 fragments of donkey anti-goat IgG (Jackson ImmunoResearch Laboratories; West Grove, PA). Sections incubated with biotin-conjugated antibodies were further stained with Texas Red-conjugated streptavidin (Amersham Biosciences). Thymic epithelial cells were detected using an FITC-conjugated anti-cytokeratin antibody (Sigma España), whereas thymocytes were stained using a purified anti-CD2 MAb (Beckman Coulter, LaBrea, CA) followed by Texas Red-conjugated multi-adsorbed F(ab')2 fragments of donkey anti-mouse IgG (Jackson ImmunoResearch Laboratories). Slides were mounted in Vectashield (Vector Laboratories) and examined on a Zeiss Axioplan-2 microscope (Zeiss; Oberkochen, Germany) from the Centro de Microscopía y Citometría, Complutense University of Madrid.
Flow Cytometry
To analyze the expression of Smo receptor on thymocytes, a three-color immunofluorescence staining was performed by incubating the cells in PBS containing 1% FCS and 0.1% NaN3 in the presence of saturating amounts of anti-Smo-PE (N-19), anti-CD4-FITC, and anti-CD8-CyChrome antibodies for 30 min at 4C. For the analysis of Ptc1 expression, an intracellular staining was performed because anti-Ptc1 (G-19) antibody recognizes an intracellular domain at the amino terminus of Ptc1. Thymocytes were incubated with anti-CD4-PE and anti-CD8-CyChrome MAbs, treated with an FACS permeabilizing solution according to the manufacturer's instructions (Becton Dickinson), and stained with anti-Ptc1-FITC antibody for 30 min. Cells were fixed (Becton Dickinson) and analyzed in a FACScalibur (Becton Dickinson) from the Centro de Microscopía y Citometría, Complutense University of Madrid. Purified anti-Smo (N-19) and anti-Ptc1 (G-19) antibodies were obtained from Santa Cruz, and conjugated to FITC or PE by Biogenesis (Poole, UK). Anti-CD4 and CD8 MAbs were obtained from Becton Dickinson.
Results
Hh Proteins Shh, Ihh, and Dhh Are Produced in Human Thymus
To assess the expression of RNAs encoding the mammalian Hh proteins, we performed RT-PCR on RNA obtained from human thymic tissue fragments. Shh, Ihh, and Dhh RNAs were all present in the human thymus (Figure 1). Further analysis using isolated thymocytes and thymic epithelial cells showed that Shh, Ihh, and Dhh RNAs were expressed in the thymic epithelium, although we were unable to detect them in thymocytes (Figure 1). The presence of specific transcripts for Hh proteins was also detected in the human thymic epithelial cell (TEC) lines P1.1A4 and P1.4D6 (Fernandez et al. 1994) (Figure 1).
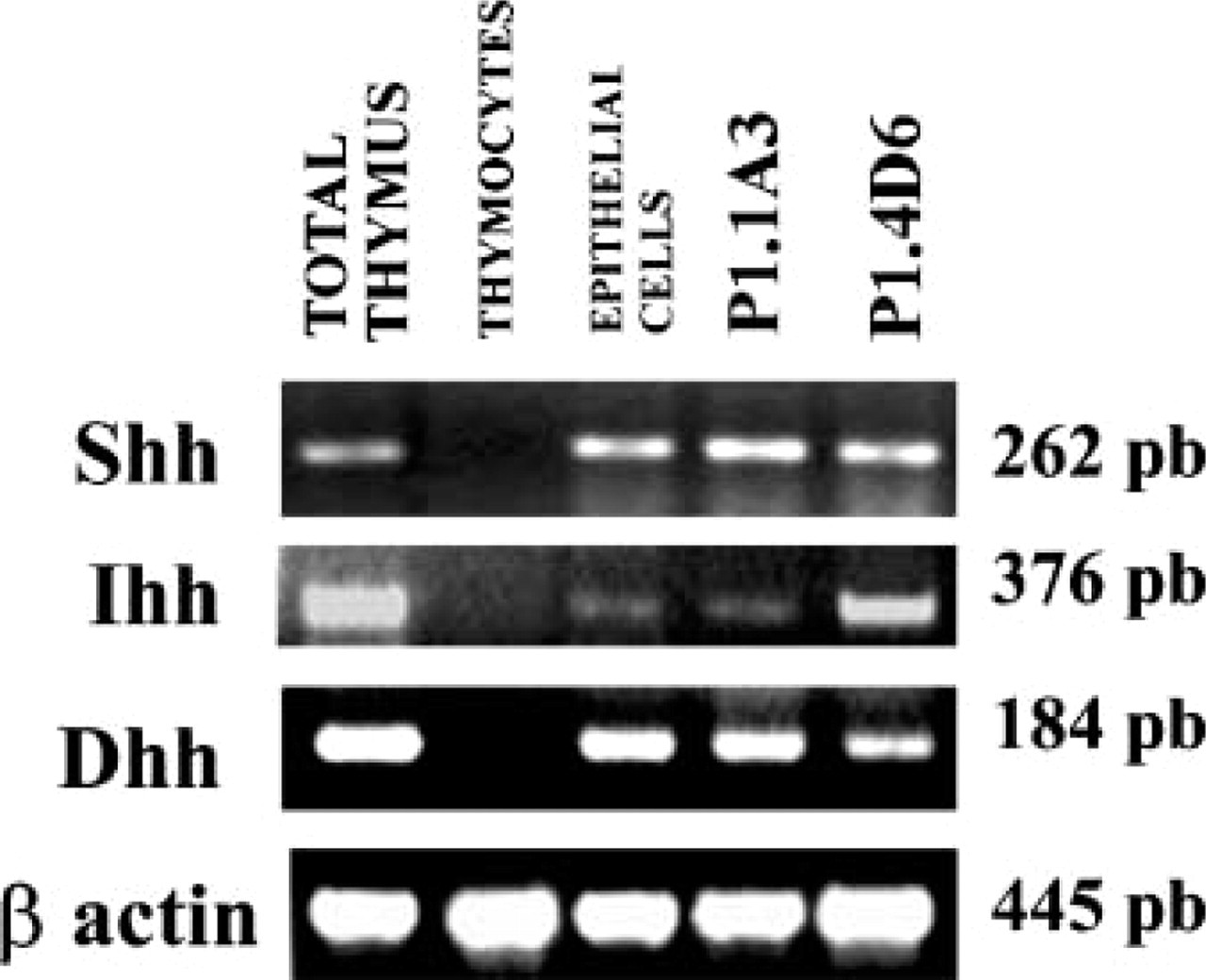
RT-PCR analysis of Hh expression in the human thymus. Primer pairs specific for Shh, Ihh, and Dhh were used to determine their presence in total thymus, isolated thymocytes, isolated thymic epithelial cells, and the TEC lines P1.1A3 and P1.4D6. β-Actin served as positive control. Band sizes are indicated.
To determine the histological localization of the epithelial cells that express the different Hh proteins, we performed double immunofluorescent stainings on human thymic tissue sections. Hh proteins are synthesized as precursor proteins that undergo an internal cleavage catalyzed by the C-terminal portion of the precursor. This process generates a C-terminal fragment that has no other known function and an N-terminal fragment that has all the known signaling activity (Lee et al. 1994). The N-terminal fragment receives two lipid modifications and, after being secreted, can tether to the membrane of producing cells or diffuse, establishing concentration gradients (Porter et al. 1996; Pepinsky et al. 1998; Lewis et al. 2001; Zeng et al. 2001). Therefore, in the immunostaining study we used antibodies that specifically recognize the C-terminal end of the human Shh, Ihh, and Dhh precursors to ensure the detection of the producing/secreting cells only. Supporting the RT-PCR results, the three proteins were expressed by cytokeratin-positive epithelial cells (Figure 2). The localization of Shh-expressing cells was restricted to the subcapsular and medullary areas (Figure 2), whereas Ihh- and Dhh-producing epithelial cells were randomly distributed throughout the thymic parenchyma, appearing as isolated cells or forming small cell clusters (Figure 2).
Human Thymocytes and Thymic Epithelial Cells Express Hh Receptors
To elucidate the putative targets for Hh signaling in the human thymus, RT-PCR was performed on different purified populations of thymocytes including CD34+ progenitor cells, immature CD4+CD8+ cells and mature CD4+CD8− and CD4−CD8+ thymocytes. We detected RNA encoding Ptc1 and Smo in all the analyzed thymocyte subpopulations, whereas Ptc2 was present only in CD34+ cells (Figure 3). Ptc 1, Ptc2, and Smo genes were also expressed in thymic epithelial cells (Figure 3). In addition, the P1.1A3 TEC line expressed all the Hh receptors, whereas we were unable to detect Ptc2 RNA on P1.4D6 cell line (Figure 3).
The proportion of Ptc1-expressing cells in the different thymocyte subpopulations was determined by flow cytometry. In agreement with the RT-PCR results, all the subpopulations defined according to CD4/CD8 expression contained Ptc1-positive cells. As shown in Figure 4, more than 85% of total thymocytes expressed the Ptc1 receptor, and similar percentages were found in all the analyzed subsets. The histological analysis confirmed that most thymocytes from the different thymic compartments expressed Ptc1, and also showed that cytokeratin-positive epithelial cells expressed high levels of the Ptc1 receptor (Figure 5).
The expression of Ptc2 was restricted to CD34+ thymic progenitor cells and cortical and medullary cytokeratin-positive epithelial cells, as demonstrated by RT-PCR and immunofluorescence (Figure 3 and 5).
On average, 35% of total thymocytes expressed the Smo receptor on their cell surface (Figure 4), a percentage similar to that described in the murine thymus (Outram et al. 2000). In the human thymus, DP and, to a lesser extent, DN thymocyte subsets contained the highest proportions of Smo+ cells (Figure 4). The double immunofluorescent analysis on thymic tissue sections, using anti-Smo antibodies combined with anti-CD2 antibodies, confirmed the expression of Smo protein in thymocytes distributed throughout the organ (Figure 5). Strikingly, the highest levels of Smo expression were associated with cell clusters composed of epithelial cells and the thymocytes interacting with them. They were located in the subcapsular, cortical, and medullary areas (Figure 5), indicating that precursor cells as well as immature and mature thymocytes may take part in them. This nested pattern of the brightest Smo expression suggests the existence of niches in which Hh signaling is stronger.
Hh-binding Molecules Are Expressed in Human Thymus
Other Hh-binding molecules, such as Hh-interacting protein (Hip) and growth arrest-specific gene-1 (Gas-1) are also expressed in the human thymus, and therefore modulate Hh signaling (Chuang and McMahon 1999; Lee et al. 2001). We found by RT-PCR Hip and Gas-1 encoding RNAs in all thymocyte subsets as well as in the thymic epithelial cell population (Figure 3). P1.1A3 and P1.4D6 TEC lines also expressed Gas-1 but not Hip (Figure 3).
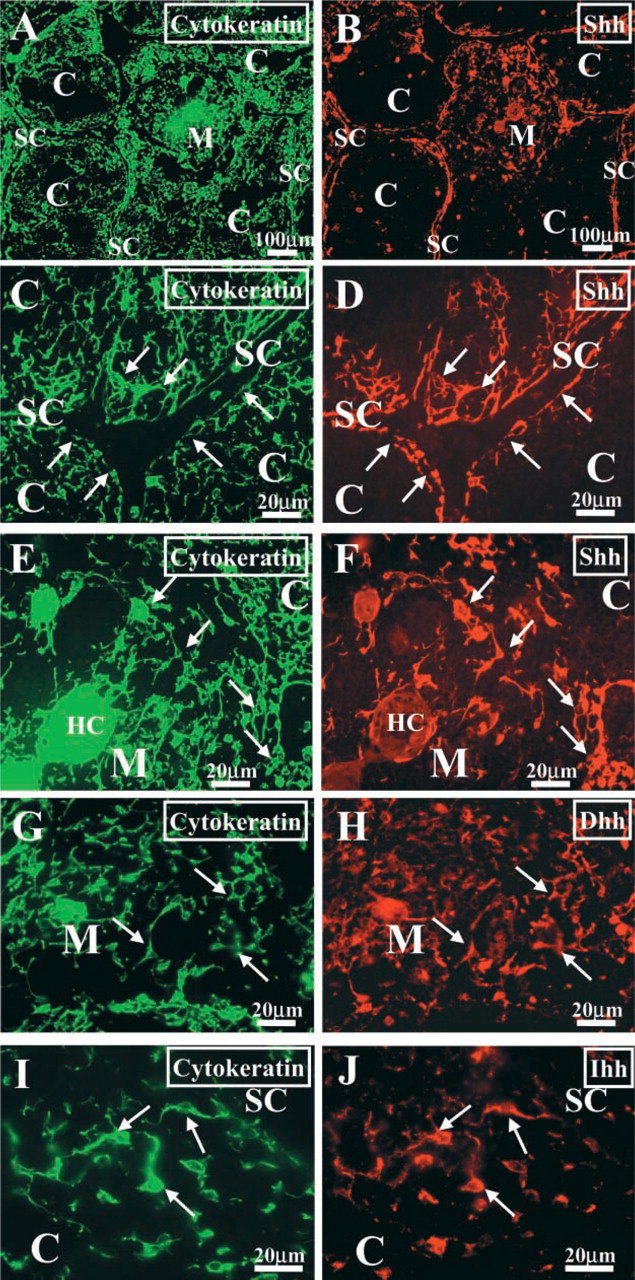
Localization of Hh proteins in the human thymus. Frozen sections of human thymus were double stained with anti-cytokeratin (green fluorescence,
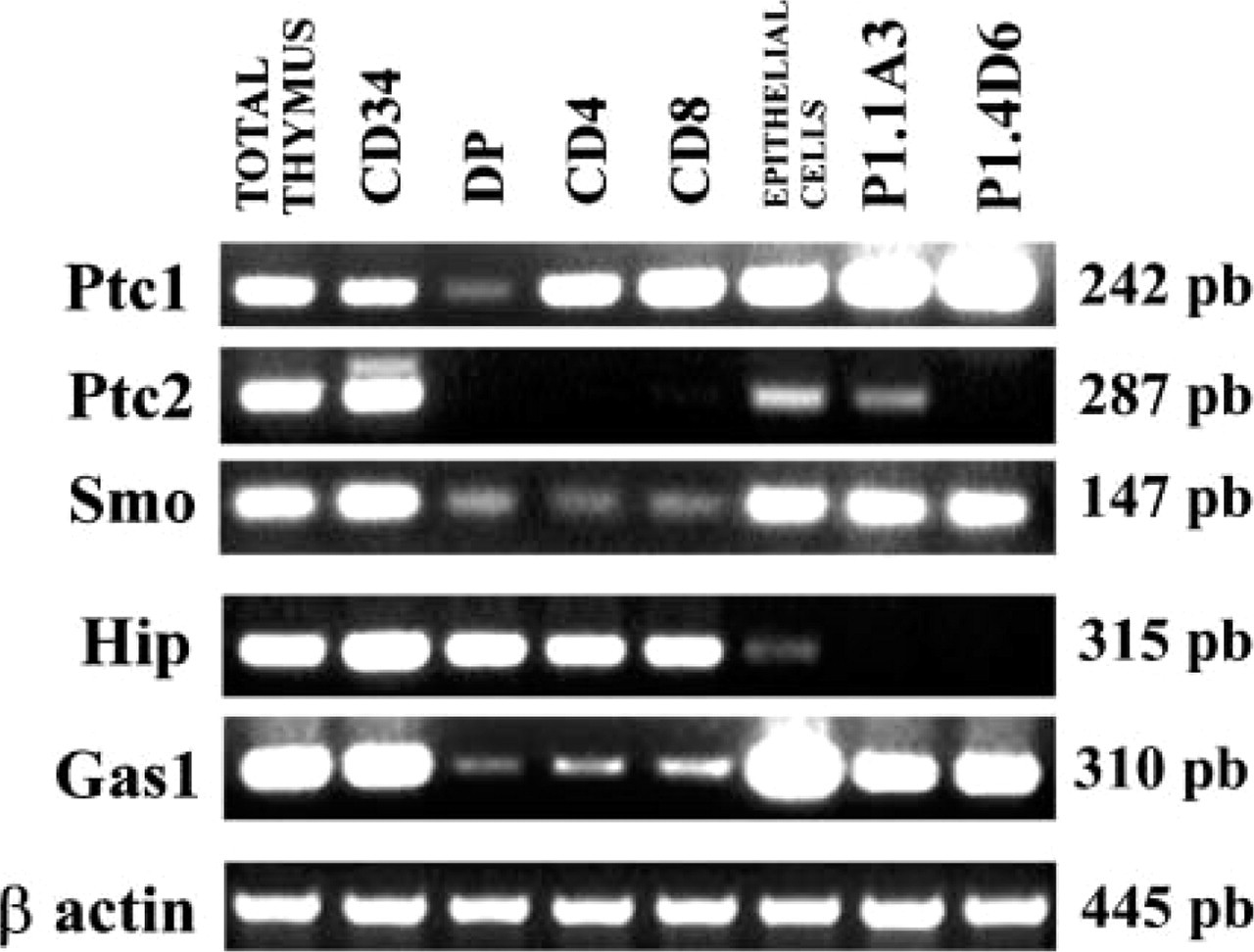
RT-PCR analysis of the expression of Hh receptors in the human thymus. Expression of Ptc1, Ptc2, Smo, Hip, and Gas-1 on whole thymus, CD34+ precursor cells, CD4+CD8+ (DP), CD4+CD8− (CD4), and CD4−CD8+ (CD8) thymocytes, epithelial cells, and P1.1A3 and P1.4D6 TEC lines. Band sizes are indicated.
Expression of Gli Transcription Factors in Human Thymus
Gli1, Gli2, and Gli3 transcription factors transduce Hh signaling to the nucleus, regulating the expression of different Hh target genes. In the human thymus, only early T-cell progenitors and the epithelium expressed the three Gli genes (Figure 6). P1.1A3 TEC line expressed Gli2 and Gli3, whereas P1.4D6 expressed only Gli3 (Figure 6).
The CD8+ SP mature thymocytes expressed Gli1 and Gli3 transcription factors, although this subpopulation expressed only low levels of the Smo receptor (Figure 4 and 6). Strikingly, we were unable to detect RNA for any of the three Gli proteins in DP and CD4+ thymocyte subpopulations (Figure 6).
Discussion
Our results show a complex intrathymic expression pattern of the molecules of the Hh cascade in human, and suggest the involvement of Hh signaling in the differentiation of human thymocytes, as previously demonstrated in the murine thymus (Outram et al. 2000). By using RT-PCR and immunofluorescence we show that human thymic epithelial cells produce the three mammalian Hh proteins, Shh, Ihh and Dhh. Thymic epithelial cells located in the subcapsulla and medulla produce the most pleiotropic of Hh proteins, Shh, whereas Ihh and Dhh production is observed in the cortex and medulla.
The distribution of Shh-secreting epithelial cells remarkably correlates with that of the epithelial cells that produce stromal cell-derived factor-1 (SDF-1) in the human thymus (Hernandez-Lopez et al. 2002). SDF-1 is a member of the chemokine CXC subfamily that we have recently demonstrated to be a survival and proliferative factor for thymic CD34+ cell precursors (Hernandez-Lopez et al. 2002). Interestingly, SDF-1 and Shh have been demonstrated to synergize during cerebellar development with SDF-1 enhancing the Shh-induced granule cell proliferation, probably via the negative regulation of PKA, a known antagonist of Hh signaling (Klein et al. 2001). The possible relationship of SDF-1 and Shh signaling pathways during the maturation of CD34+ thymic precursors could therefore be of relevant interest.
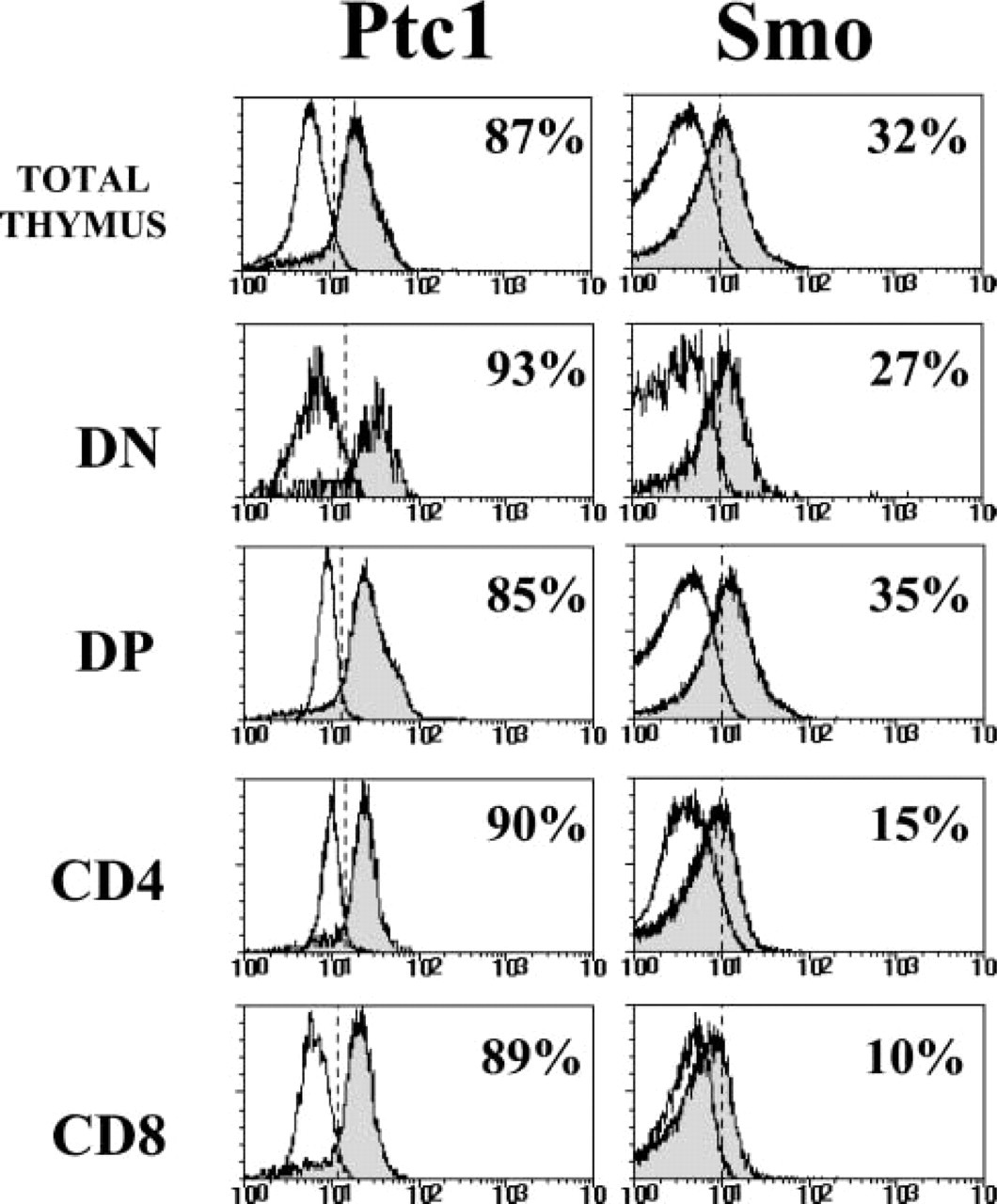
Expression of Ptc1 and Smo in human thymocytes. A three-color flow cytometry analysis was performed by staining thymocytes with anti-CD4, anti-CD8, and either anti-Ptc1 or anti-Smo antibodies. Histograms show the expression of Ptc1 and Smo (gray profiles) in whole thymus and gated CD4−CD8+ (DN), CD4+CD8+ (DP), CD4+CD8− (CD4), and CD4−CD8+ (CD8) thymocyte subsets. White profiles represent background fluorescence using irrelevant antibodies.
The murine thymus has been also described to produce Hh proteins (Outram et al. 2000). Mouse thymic epithelial cells produce only Shh, whereas Ihh is associated with medullary blood vessels. The distribution of Shh-positive cells in the murine thymus reported in our previous work (Outram et al. 2000) slightly differs from that described here for the human thymus, because we detected Shh-positive epithelial cells throughout the murine thymic parenchyma including the thymic cortex, although mainly in the outer area. The different pattern of distribution of Shh-expressing cells in the mouse thymus is caused by the use of antibodies recognizing the functionally active N-terminal fragment of Shh. Therefore, in the murine thymus we detected not only the secreting cells but also those cells that retain Shh on their membranes or have endocytosed the protein. When we stained murine thymic cryosections with antibodies that bind to the C-terminal domain of Shh, we found that the localization of the epithelial cells that produce Shh is similar to that described here in the human thymus (unpublished observations).
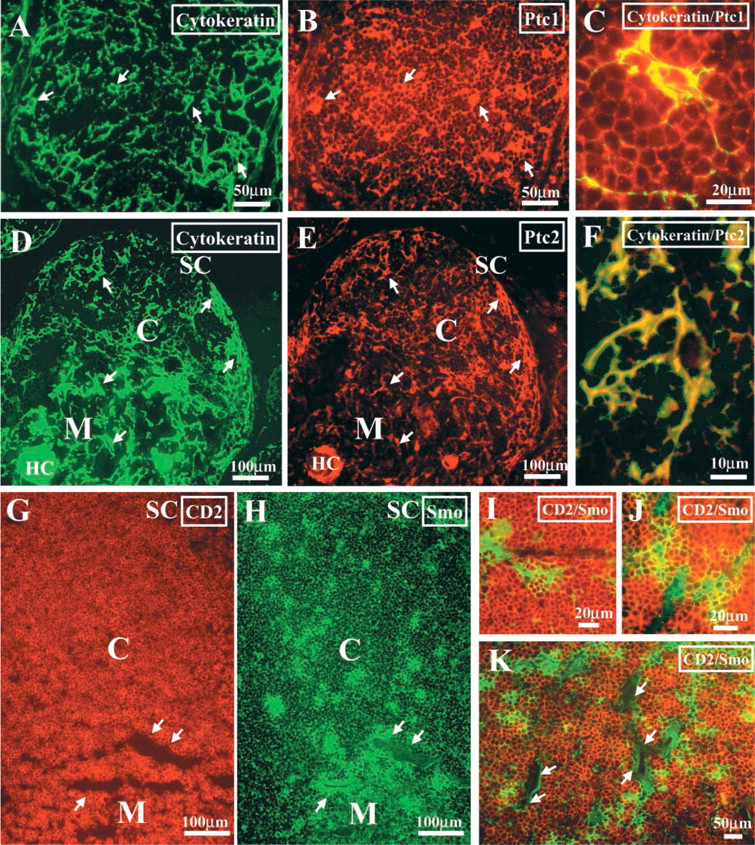
Localization of Hh receptors in the human thymus. Human thymic cryosections were double stained with anti-cytokeratin (green fluorescence,
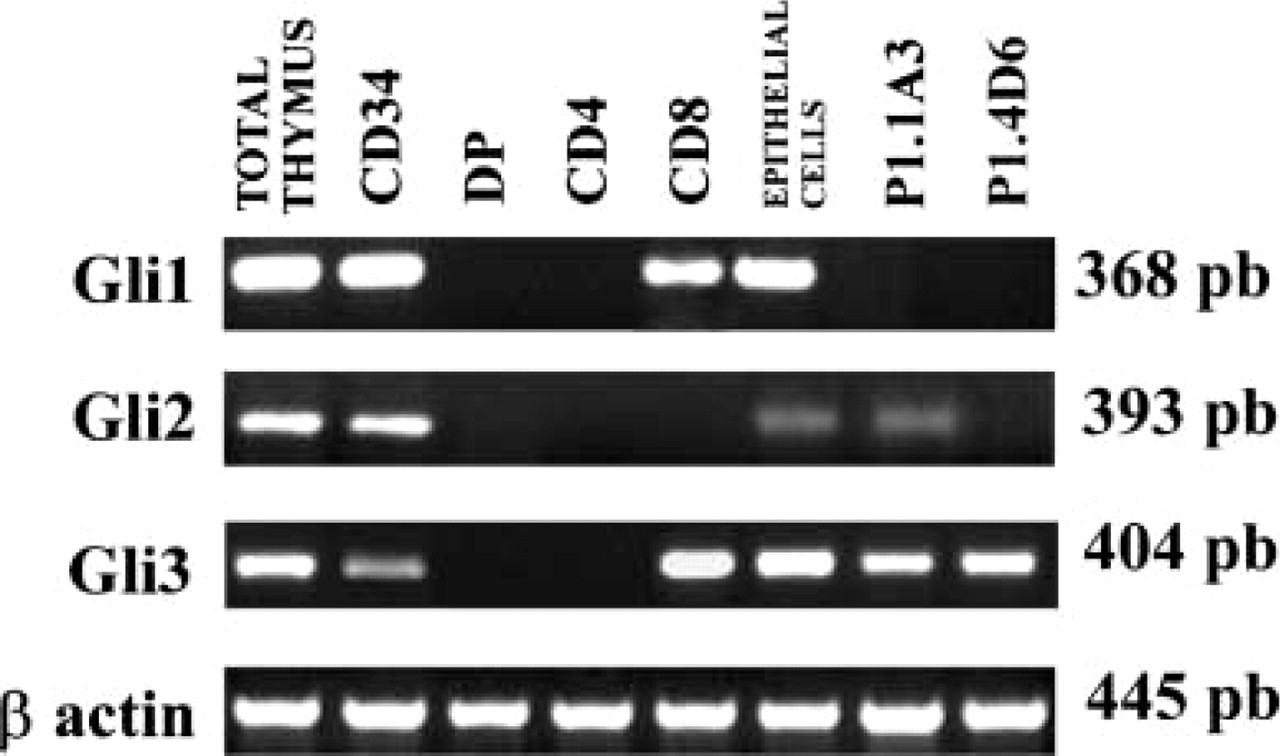
RT-PCR determination of the existence of Gli transcription factors in the human thymus. RT-PCRs were performed on cDNA from whole thymus, CD34+ precursor cells, CD4+CD8+ (DP), CD4+CD8− (CD4), and CD4−CD8+ (CD8) thymocytes, epithelial cells, and TEC lines P1.1A3 and P1.4D6 to show the expression of the transcription factors Gli1, Gli2, and Gli3. Band sizes are indicated.
The evidence of the existence of the three Hh proteins in the thymus is interesting. Although all these molecules bind to Ptc with the same affinity and share the same signaling cascade, they exhibit specific functions, at least during development, as clearly demonstrated by their distinct spatial and temporal expression patterns (Bitgood and McMahon 1995). Moreover, the signaling potencies for Shh, Ihh, and Dhh have been demonstrated to be different using several in vitro assays (Pathi et al. 2001). The expression in the same tissue of different Hh proteins has been previously described in bone, cartilage, kidney, heart, gut, and eye (Ingham and McMahon 2001). In some organs, such as heart and gut, the different expressed Hh proteins show unique or redundant activities (Ramalho-Santos et al. 2000; Zhang et al. 2001). Conversely, in growth plate chondrocytes Ihh and Shh have different and even opposite functions, and are differentially regulated by retinoic acid (Wu et al. 2002). In the human thymus, although Shh, Ihh, and Dhh could have overlapping functions, it is reasonable to think that their expression is differentially regulated, allowing the existence of specific microenvironments with different levels of Hh signaling, which would be associated with concrete processes leading to thymic homeostasis.
Human thymic epithelial cells also express the Hh receptors Ptc1, Ptc2, and Smo, as well as the Gli transcription factors, which suggests an autocrine role of Hh proteins on thymic epithelium. Furthermore, Hh expression in long-term established TEC lines indicates that this expression is independent of sustained epithelium-thymocyte cross-talk, and that Hh proteins are produced by thymic epithelial cells in a cell-autonomous manner.
Human thymocytes do not produce Hh but express the specific receptors needed for receiving and transducing Hh signals derived from the epithelium. The flow cytometry analysis shows that most thymocytes express Ptc1. As in other tissues, the broad expression of Ptc1 in the thymus may have a dual function of both repressing Smo signaling and sequestering free Hh protein. The basic role of Ptc1 in Hh signaling is the repression of Smo in the absence of Hh, which is abrogated after Hh binding, allowing Smo to signal (Ingham and McMahon 2001). However, the Hh pathway positively regulates Ptc1 expression (Goodrich et al. 1996; Marigo and Tabin 1996) and so a negative feedback loop is created because Ptc1 can also bind Hh proteins and induce their endocytosis, limiting the availability and the movement of Hh proteins from their source (Incardona et al. 2002; Nybakken and Perrimon 2002). The possibility that the endocytosis, of Ptc1/Hh complexes triggers other signaling pathways cannot be discarded (Incardona et al. 2002).
Ptc2 receptor is also expressed in the human thymus. Ptc1 and Ptc2 expression overlaps in CD34+ thymic progenitor cells and in cortical and medullary cytokeratin-positive epithelial cells. The specific role of Ptc2 in the Hh pathway remains unclear. Ptc2 has some characteristics similar to Ptc1, such as the extracellular loops that mediate the interaction with Hh proteins and the modulation of its expression by Hh signaling (Stone et al. 1996; Zaphiropoulos et al. 1999). However, Ptc2 is not able to replace Ptc1 function in cancerous cells (Zaphiropoulos et al. 1999) and during mouse development (Stone et al. 1996), implying that Ptc2 must have functions related to but also distinct from those of Ptc1.
The expression of Smo is more restricted than that of Ptc1 and is mainly associated with both DP and DN thymocytes. In mice, most Smo-expressing cells appear in the CD25+ DN thymocyte subset, which is therefore the main cell target for Shh activity (Outram et al. 2000). However, these differences in the distribution of Smo-expressing cells do not necessarily imply that Hh regulates distinct events during thymocyte differentiation in mouse and human. In fact, murine CD25+ DN thymocytes and human early DP thymocytes represent equivalent thymocyte subsets because they are the developmental stages in which the β-selection process is taking place (Carrasco et al. 2002; Spits 2002).
Interestingly, CD34+ thymic progenitors express Ptc and Smo receptors as well as the three Gli transcription factors, pointing to a role of Hh proteins from the earliest steps of human T-cell development.
In the human thymus Hh signaling is modulated by the expression, in both thymocytes and epithelial cells, of Hip and Gas1. Hip is a membrane protein positively regulated by Hh signaling. It binds to all mammalian Hh proteins with the same affinity as the Ptc1 receptor. Thus, Ptc1 and Hip compete for binding to the ligand, but whereas Ptc1 regulates Smo activity, Hip exclusively sequesters the ligand attenuating Hh signaling (Chuang and McMahon 1999; Chuang et al. 2003). Conversely, the role of Gas-1 in Hh signaling is still unclear (Mullor and Ruiz i Altaba 2002). The Gas family of proteins is known to include anti-tumor growth markers, mainly related to G0-arrested cells. However, it has recently been demonstrated that Gas-1 is a Wnt-target gene that binds to Shh, blocking its growth-stimulating activity (Lee et al. 2001). Moreover, Gas-1 expression is not restricted to non-proliferating cells during development (Lee and Fan 2001), which suggests that Gas-1 is not a mere growth-arrest factor. Its possible role in the Hh-dependent modulation of T-cell differentiation remains to be elucidated.
Gli1, Gli2, and Gli3 transcription factors are important mediators of Hh signaling. They exhibit only partially overlapping functions and, whereas Gli1 and Gli2 are Hh-dependent activators of Hh targets, Gli3 acts mainly as a repressor. Moreover, Gli2 might be a Hh-independent repressor (Ruiz i Altaba 1999; Aza-Blanc et al. 2000; Wang et al. 2000; Bai et al. 2002; Mullor et al. 2002). In the human thymus, only early T-cell progenitors and the epithelium expressed the three Gli genes. These findings could indicate the occurrence of a complex Hh signaling response in these thymic cells. Similarly, early hematopoietic cell progenitors and stromal cells from human bone marrow expressed Gli1, Gli2, and Gli3 (Bhardwaj et al. 2001). However, these results might merely reflect the intrinsic heterogeneity of these cell populations. Supporting this, P1.1A3 TEC line expressed Gli2 and Gli3, whereas P1.4D6 expressed only Gli3.
In contrast, we could not detect RNA for any of the three Gli proteins in DP and CD4+ thymocyte sub-populations. Bhardwaj et al. (2001) also showed that human CD3+ and CD19+ peripheral lymphocytes expressed Ptc1 and Smo receptors and produced Shh, but did not express RNAs for any Gli protein, in spite of the fact that human CD4+ T-cells have also been described to respond to Shh (Stewart et al. 2002). These findings raise questions about how these cells transduce the Hh signal. Several reports have indicated that Hh signaling is not restricted to the Gli family of transcription factors. The first evidence was the Shh-induced expression of chicken ovalbumin upstream promoter-transcription factor II (COUP-TFII) during the development of the neural tube, because the promoter of this gene has no Gli response element and because this induction is independent of protein synthesis (Krishnan et al. 1997). In addition, the phenotype of Gli1/Gli2 double mutant mice is less severe than that of Shh−/- mice (Chiang et al. 1996; Bai et al. 2002), and the loss of Gli1 or Gli2 in Ptc1−/- mice does not affect or does not completely rescue, respectively, the altered phenotype associated with Hh signaling (Bai et al. 2002). Although this might suggest the importance of Gli2 and Gli3 repressor activities, some of the alterations of Gli 2/Ptc double mutant mice are not found in Gli 3−/- mice (Bai et al. 2002). Likewise, Gallet et al. (2000) have demonstrated during Drosophila embryogenesis that Hh requires a transduction pathway independent of Cubitus interruptus, the Drosophila homologue of Gli transcription factors. A detailed study of the Hh signaling pathways functioning in DP and CD4+ SP human thymocytes could contribute to our knowledge about Hh signaling.
Footnotes
Acknowledgements
Supported by grants PM99-0060 and BMC2001-2025 from the Ministerio de Ciencia y Tecnología, and by CAM08.3/0041/2000 and CAM08.3/0018.1/2001 from the Comunidad Autónoma de Madrid. T.C. is supported by the Wellcome Trust. This work was also supported by Acciones Integradas Hispano-Británicas.
We thank Dr F. Villagrá and the Pediatric Cardiosurgery Units from Hospital La Zarzuela and Hospital Madrid-Montepríncipe for the thymus samples, and Dr M.L. Toribio for the generous gift of thymic epithelial cell lines.