
Abstract
Caveolin-3, a muscle-specific member of the caveolin family, is strongly localized to the neuromuscular junction (NMJ) in adult rat muscle fibers, where it co-localizes with α-bungarotoxin staining. In 24-month-old rats, less distinct staining corresponds with the normal aging changes in the NMJ. After denervation, the pattern and intensity of staining begin to break up as early as 3 days, and by 10 days little staining remains. The functional implications of this concentration of caveolin-3 at the NMJ remain obscure, but it is possible that its absence could account for some of the phenotypic characteristics of individuals with caveolin-3 mutations.
Keywords
T
Caveolin-3 interacts with dystrophin and the dystroglycan complex (Song et al. 1996), and it has recently been found that patients with certain muscular dystrophies exhibit abnormal amounts of caveolin-3 (Tateyama et al. 2002). Limb–girdle muscular dystrophy, in particular, is caused by deletion or missense mutations in the caveolin-3 gene (Minetti et al. 1998), and increased numbers of caveolae and amounts of caveolin-3 are found in Duchenne muscular dystrophy (Repetto et al. 1999). Recent research on mutants of caveolin-3 has demonstrated dystrophy-like phenotypes in mice (Galbiati et al. 2000). Mutations of caveolin-3 have also been found to be associated with rippling muscle disease, a rare condition characterized by electrically silent irritability of muscle fibers (Betz et al. 2001).
Despite the many associations of caveolin-3 with normal and abnormal cellular functions, much remains to be learned about the role of this molecule in cell physiology. While engaged in a study involving the biology of isolated normal and denervated skeletal muscle fibers, we noted that the neuromuscular junctions (NMJ) stain intensely with an antibody against caveolin-3. This report describes our findings.
Materials and Methods
Animals and Surgical Techniques
The material used in this study was obtained from male rats of the Wistar and Sprague–Dawley strains ranging in age from newborn to over 2 years. All experimentation was done according to protocols approved by the animal care and use committees of the University of Michigan and the University of Otago. All operations were carried out under ether anesthesia, and animals were sacrificed by exposure to carbon dioxide or an overdose of ether.
For short-term denervations (less than 10 days), the right sciatic nerve was surgically exposed and a 1-cm segment was removed from the upper thigh. For longer-term denervations (2 weeks to 2 months), the sciatic nerve in the upper thigh was tightly ligated in two places with 6-0 silk suture and then transected between the two sutures. Each cut end of the nerve was reflected away from the suture site and embedded within a muscle. This technique allows permanent denervation in adult rats (Carlson and Faulkner 1988). All animals were 4 months old at the time of denervation. For all cases reported here, the extensor digitorum longus (EDL) muscle was used for analysis of single fibers and either the EDL or tibialis anterior was used for the analysis of sections. Muscles from 75 rats were used in this investigation.
Muscle Fiber Isolation
On removal from the animals, the muscle was maintained at resting length and temporarily placed in cold (4C) 0.1 M PBS for no more than 30 min. It was then incubated in 0.2% collagenase type I (Worthington Biochemicals; Lakewood, NJ) in PBS at 37C in a gently shaking incubator for approximately 3 hr. The most effective length of incubation depends on the age of the animal and the time of denervation of the muscle because of the amount of interstitial collagen present. On removal from the incubator, the muscle was rinsed twice in cold PBS and then given an additional 10-min rinse in cold PBS to eliminate remaining collagenase activity. The muscle was then fixed for 60-90 min in cold (4C) freshly made 2% paraformaldehyde (staining disappears with fixation in 4% paraformaldehyde), quickly rinsed twice in cold PBS, and then rinsed twice for 10 min in cold 0.75% glycine/saline solution to remove remaining paraformaldehyde. The muscle was then left overnight in cold glycine/saline, after which the glycine/saline was replaced by PBS. Individual muscle fibers were then mechanically separated under a dissecting microscope with sharpened tungsten needles and placed in drops of water on microscope slides. Muscle fibers will not stick to the slides if they are placed in saline. The muscle fibers were then allowed to dry on the slides and were stored for up to several months in slide boxes at −20C.
Immunostaining of Isolated Muscle Fibers
The isolated muscle fibers dried on slides were rehydrated in PBS and incubated for 60 min in 5% normal sheep serum at room temperature (RT). The fibers were then incubated overnight at 4C with 1:500 mouse anti-rat caveolin-3 primary antibody (Sapphire Bioscience; Crows Nest, NSW, Australia). After rinsing in PBS, the fibers were then exposed for 60 min at RT to an anti-mouse biotin secondary antibody (Amersham; Piscataway, NJ). After further rinsing in PBS, the fibers were exposed to a methanol/H2O2 mixture twice at 5 min each and then rinsed in PBS. The fibers were then incubated in a tertiary peroxidase-conjugated streptavidin-biotin antibody (1:200; Jackson Laboratories, Bar Harbor, ME) for 60 min at RT, washed, bathed in either diaminobenzidine or 3-amino-9-ethyl carbazole (Sigma; St Louis, MO), washed again in water, and coverslipped. Appropriate controls, such as elimination of the primary antibody and controls for endogenous peroxidase activity, were performed.
Immunohistochemistry of Tissue Sections and Immunofluorescence Microscopy
For analysis of sectioned muscle, the muscles were fixed in freshly prepared 2% paraformaldehyde in 0.1 M PBS for 24 hr at 4C, washed overnight in PBS, cryoprotected in a graded sucrose series, placed in specimen molds containing TBS/Tissue Freezing Medium (Triangle Biomedical Sciences; Durham, NC), and oriented for longitudinal sectioning. The samples were quickly frozen in isopentane that had been pre-cooled by dry ice. Longitudinal 9.0-μm sections were cut with a cryostat and mounted on glass slides. Before staining, the sections were fixed in 100% methanol at −20C for 10 min. The slides were allowed to air-dry and then the sections were re-hydrated in PBS, treated with 1% Triton X-100 in PBS for 10 min, washed in PBS, and incubated with 10% normal goat serum for 20 min at RT. The sections were labeled with a mouse anti-rat caveolin-3 primary antibody (5 μg/mL; Transduction Laboratories, Lexington, KY; this is the same antibody as that obtained from Sapphire Bioscience for staining isolated fibers) for 3 hr at 37C. After staining the sections were washed in PBS, incubated with 10% normal goat serum for 10 min at RT, and labeled with an FITC-conjugated goat anti-mouse secondary antibody (Jackson ImmunoResearch Laboratories; West Grove, PA) for 45 min at RT. For control, omissions as well as substitution with normal goat serum of a primary antibody were used. To indicate the area of the postsynaptic membrane in an NMJ, an α-bungarotoxin tetramethylrhodamine conjugate (Molecular Probes; Eugene, OR) was used during incubation with the secondary antibody. After staining with a secondary antibody the sections were rinsed in PBS, mounted in Vectashield mounting medium for fluorescence with DAPI (Vector Laboratories; Burlingame, CA) to counterstain nuclei, and were cover-slipped. Observation of the sections was done with a Zeiss Axiophot-2 Universal Microscope (Carl Zeiss; Oberkocher, Germany) equipped with Cy3, FITC, and UV filters as well as with differential interference contrast (DIC). Fluorescence and DIC images from the same optical fields were captured into a computer using a Zeiss Axiocam digital camera. Figures were prepared from electronic images using Adobe Photoshop software (Adobe Systems, San Jose, CA).
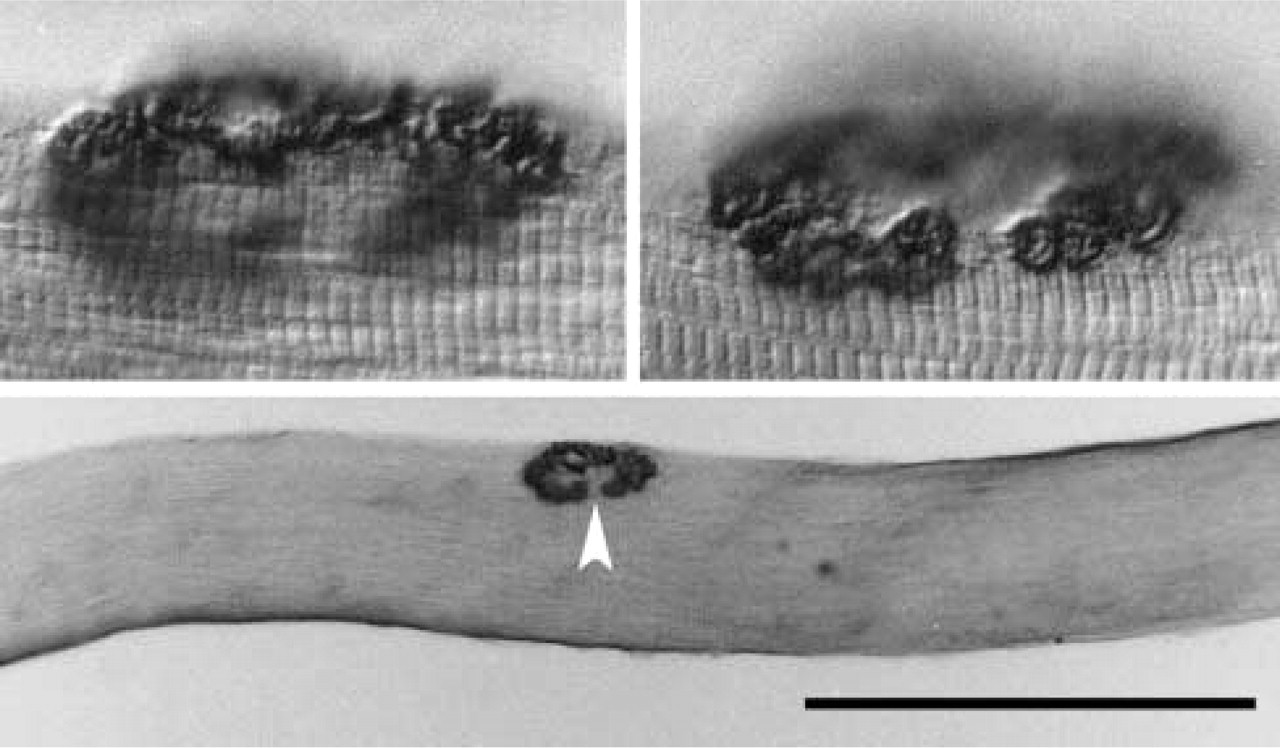
Localization of caveolin-3 staining at the NMJ (arrowhead) of an isolated fiber from the EDL muscle of a 4-month-old rat. The upper photos are higher-power views at different focal planes of the same NMJ. Bar = 500 μm.
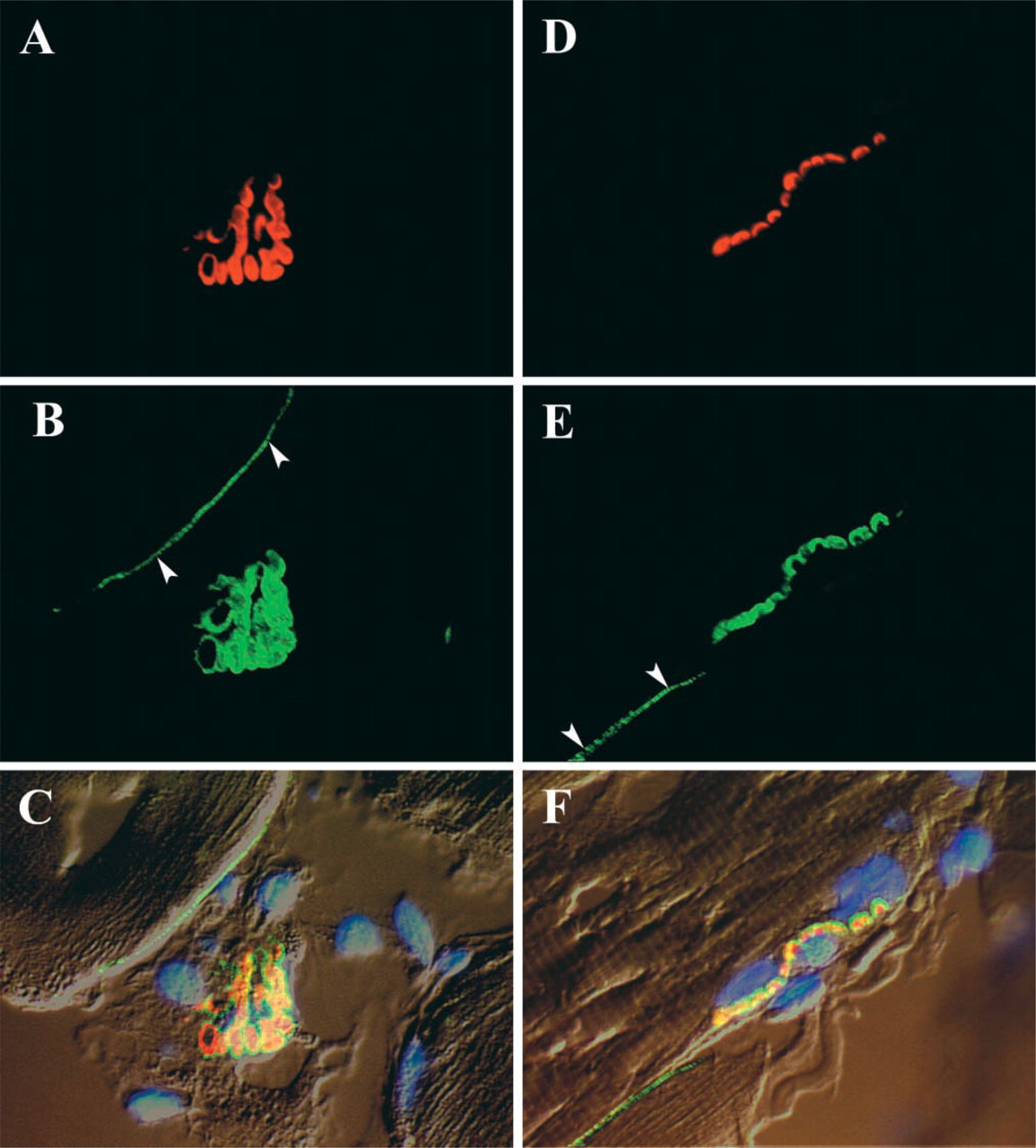
En face (
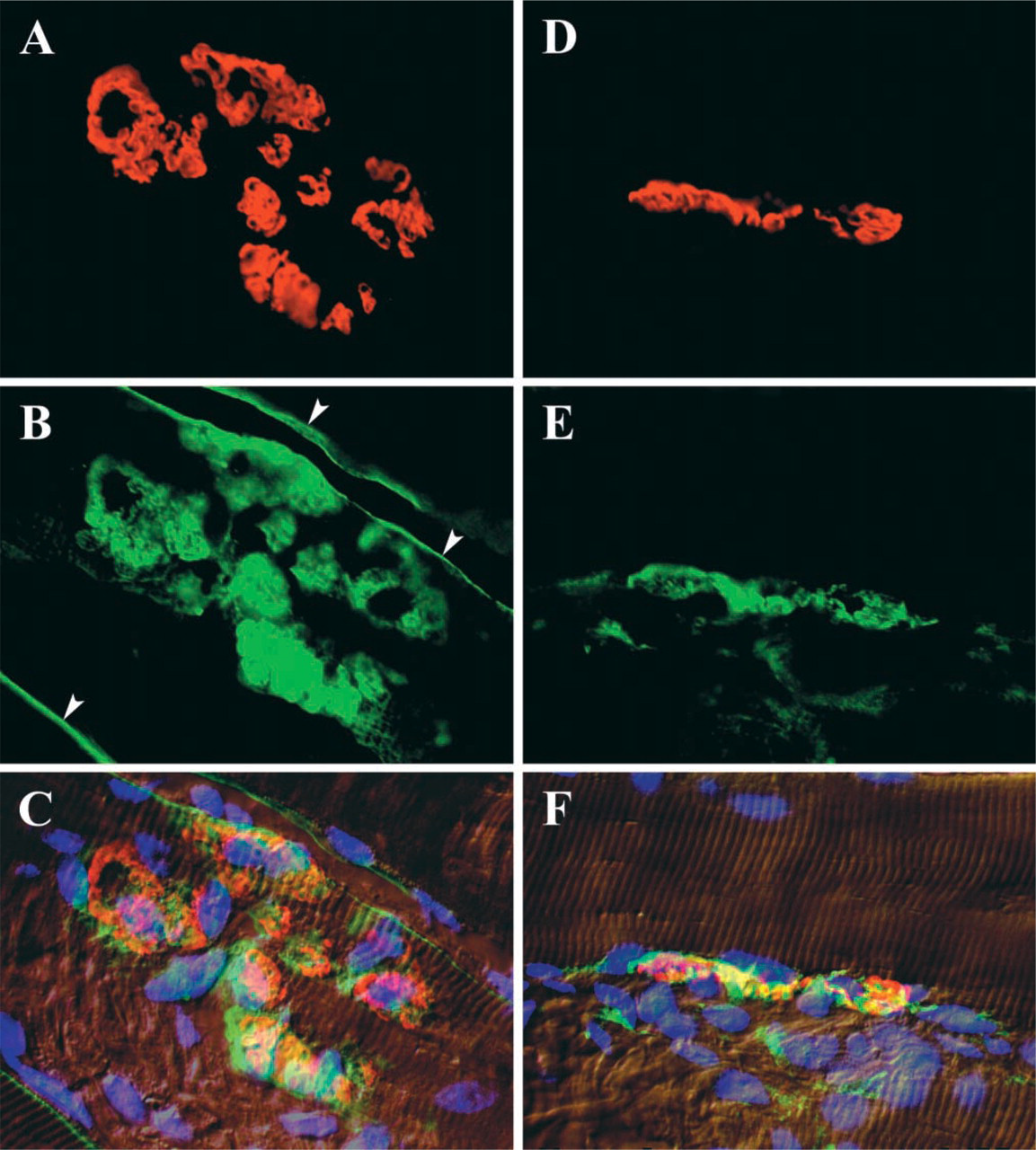
En face (
Results
As shown in Figure 1, caveolin-3 is strongly localized to the NMJ of normal adult muscle fibers. In sectioned material (Figure 2), the concentrated staining at the NMJ stands out in sharp contrast to that of the remainder of the plasma membrane, in which caveolin-3 staining is limited to a fine line in the position of the sectioned sarcolemma. Caveolin-3 staining co-localizes with the distribution of α-bungarotoxin, although it always occupies an area slightly greater than that occupied by the acetylcholine receptors, as indicated by the bungarotoxin staining. During aging, the outlines of the NMJ are not as distinct as those in young adult animals and there is less exact correspondence between the regions stained by anti-caveolin-3 and those stained by α-bungarotoxin (Figure 3).
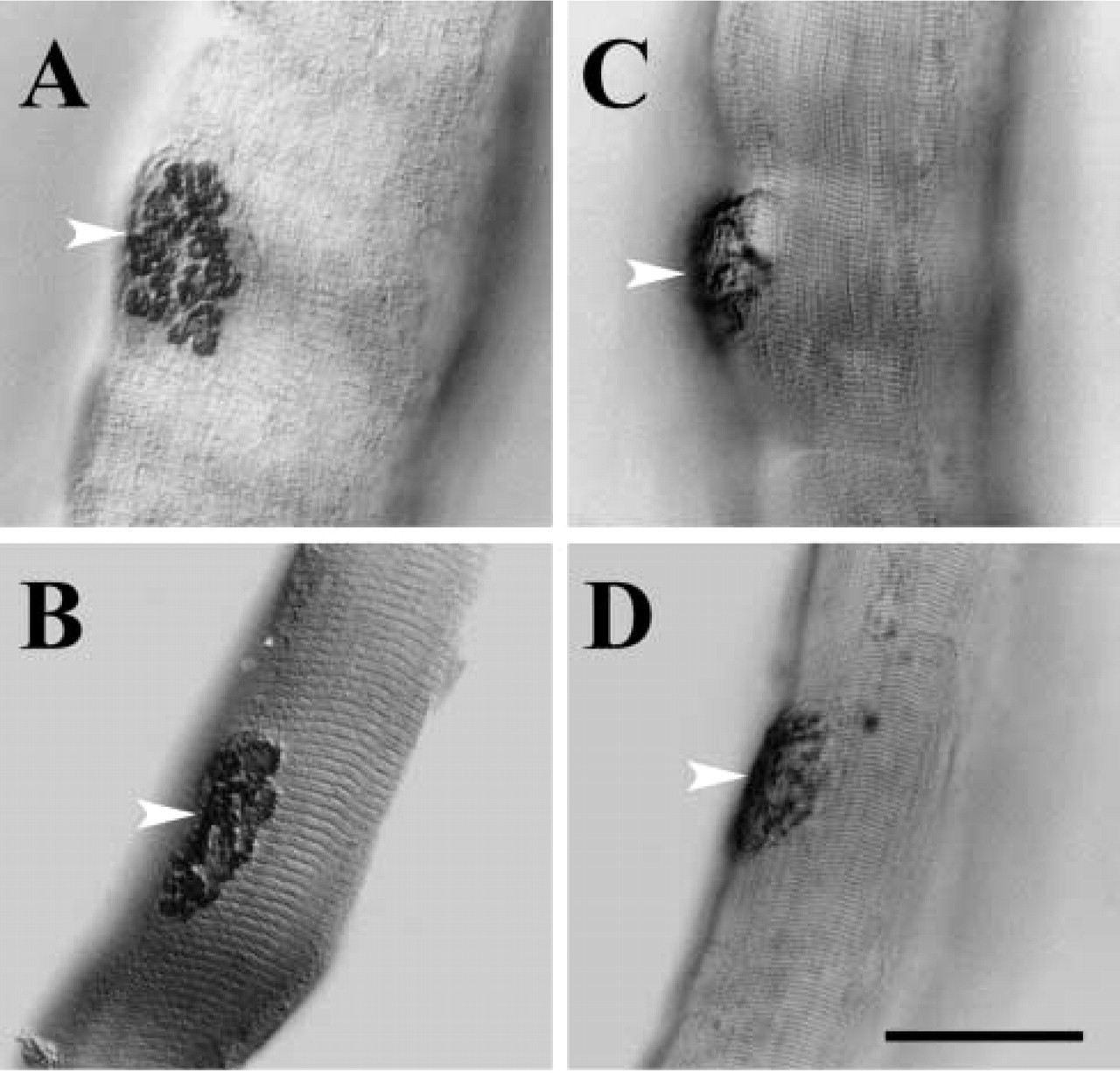
After denervation of a muscle, the caveolin-3 staining of the NMJ rapidly becomes less distinct. By 3 days (Figure 4) the overall outlines and the outlines of the individual gutters in the NMJ have already begun to break up, and by 10 days the denervated NMJ is represented only by isolated patches of staining.
Discussion
Although the existence of vesicles similar in appearance to caveolae has been illustrated in electron micrographs of the NMJ (e.g., Engel 1994), little attention has been paid to them in the scientific literature. In contrast, the presence of caveolae and caveolin-3 in the remainder of the plasma membrane of the muscle fiber has received intensive analysis (Voldstedlund et al. 2001; Ockelford et al. 2002). The intense staining for caveolin-3 at the NMJ demonstrated in the present study points to a significant concentration of this molecule at the NMJ, but at the light microscopic level the basis for this cannot be determined. Although high-power electron micrographs do not suggest an unusually high concentration of caveolae in relation to surface membrane area, the great structural complexity of the postsynaptic folds in normal rat muscle (Matsuda et al. 1988; Ogata 1988; Oki et al. 1990) could increase the total membrane surface in the area of the NMJ to the point where it would be reflected as an intense concentration at the light microscopic level. In the absence of immunoelectron microscopy it is not possible to determine if caveolin-3 is associated with other subcellular structures in the NMJ.
Anti-caveolin-3-stained NMJs of normal (
The loss of detail in the localization of caveolin-3 in the aging NMJ reflects the overall aging changes in that structure (Balice–Gordon 1997; Cardasis and LaFontaine 1987; Oki et al. 1999), where in old rats considerable structural remodeling of the junction takes place. Kawabe et al. (2001) noted a downregulation of caveolin-3 expression in muscle from young adults to older adults, as well as upregulation from neonates to young adults. This roughly corresponds to the patterns seen in staining of the NMJs, because NMJ staining for caveolin-3 in neonates is considerably less intense than in adults (unpublished observations). Ratajczak et al. (2003) reported some dissociation of caveolin-3 from sarcolemmal caveolae in aging cardiac muscle.
Similarly, after denervation the axon terminals degenerate within a day or two, and by 3 days the postsynaptic apparatus already shows significant degenerative changes, in particular a reduction in the height and complexity of the postsynaptic folds (Brown et al. 1982; Lentz 1972; Pulliam and April 1979). In these earlier studies, no comment was made on the effect of denervation on the caveolar content of the postsynaptic membrane.
In relation to the many possible functions that have been ascribed to the caveolae in skeletal muscle, it would be instructive to investigate whether or not any of the phenotypic effects that attend disruptions of caveolae or caveolin-3 synthesis in muscle might have a basis in the concentration of caveolin-3 that is found in the normal NMJ. For example, in rippling muscle disease, which is associated with caveolin-3 mutations (Betz et al. 2001), increased inducibility of nNOS (nitric oxide synthetase) due to the absence of the inhibitory effect of caveolin-3 has been suggested as a possible factor that might co-determine the phenotypic outcome of the disorder. Because nNOS is known to be localized at the NMJ and probably at the postsynaptic surface (Kusner and Kaminski 1996), it may be profitable in this and some of the dystrophies linked to caveolin-3 to investigate whether the concentration of caveolin-3 at the NMJ might play a role in the symptoms.
Footnotes
Acknowledgements
Supported by NIH grant PO1 AG-10821 and an NIH Fogarty International Fellowship (FO6 TW0-2320) to BMC, and by the Health Research Council of New Zealand to IM.