
Abstract
We investigated by immunohistochemistry (IHC) the distribution of caldendrin, the founding member of a novel family of neuronal calcium-binding proteins closely related to calmodulin, in human forebrain. Caldendrin immunoreactivity was unevenly distributed, with prominent staining in the paleo- and neocortex, hippocampus, and hypothalamus. With the exception of the hypothalamus, labeling was restricted to the somatodendritic compartment of neurons. This distribution completely matches that reported in rat, indicating that the cellular function is most likely conserved among species. Therefore, one prerequisite for functional studies in rodent models aimed at elucidation of mechanisms with relevance for humans can be based on the present findings.
T
Analysis of caldendrin's cellular functions requires the use of rodent animal models. Previous studies, however, have shown astonishing species differences in the expression of CaBPs, which ultimately limit the significance of rodent studies in terms of generalization to human conditions (Heizmann and Braun 1995). We therefore undertook the effort to localize caldendin immunoreactivity (IR) in human forebrain.
All brains were obtained from pathologists with full consent of each family and in accordance with the ethics and rules outlined by German law and the local ethics commission of the University of Magdeburg. Brains of three individuals without neurological or psychiatric disorders (two men, 49 and 63 years, and one woman, 54 years) were studied. Brains were removed within 48 hr after death. The tissue preparation was performed as described earlier (Bernstein et al. 1998). Briefly, brains were fixed in toto in 8% phosphate-buffered formaldehyde (pH 7.0) for 2 months. After embedding the brains in Paraplast, serial coronal 20-μm-thick sections were cut on a microtome and mounted. Every fiftieth section was stained for morphological orientation (combined cresyl violet and myelin staining according to Nissl and Heidenhain-Woelcke).
For Western blots, frozen tissue from the dorsolateral prefrontal cortex (female, 58 years, tissue obtained 21 hr after death) was homogenized in 20 mM Tris-HCl, pH 7.3, containing a protease inhibitor cocktail (Complete; Roche, Mannheim, Germany) and centrifuged at 100,000 × g for 15 min at 4C to separate soluble proteins and the insoluble pellet. Rat prefrontal and frontal cortex was used for comparison. After SDS-PAGE on 5–20% gels, proteins were blotted onto nitrocellulose, blocked with 5% non-fat dry milk, and incubated in 1:200 diluted rabbit anti-caldendrin antiserum (purified IgG fraction/concentration 1μg/μl) overnight. Detection was performed with HRP-conjugated anti-rabbit secondary antibody and the ECL detection kit (Amersham Biosciences; Freiburg, Germany). Caldendrin-IR was detected with the same rabbit polyclonal antiserum (Seidenbecher et al. 1998) using the nickel-amplified avidin-biotin technique as described elsewhere (Bernstein et al. 1999). The antiserum was generated using a recombinant rat C-terminal fusion protein and does not recognize CaM (Laube et al. 2002). For immunohistochemical staining sections were selected from regions 1 cm rostral to the genu to ∼1.5 cm posterior to the splenium of the corpus callosum at intervals of about 1.2 cm. After preincubation of the sections with methanol/H2O2 to block endogenous peroxidases and repeated washing with PBS, the antibody was applied at a working dilution of 1:100 in PBS. Thereafter the sections were processed with the avidin-biotin method (Vectastain-peroxidase kit; Vector, Burlingame, CA) and the reaction product was visualized with 3,3′-diaminobenzidine. The color reaction was enhanced by adding 2 ml of a 0.5% (v/v) nickel ammonium sulfate solution to the diaminobenzidine (Hsu and Soban 1982). Specificity of the immunoreaction was controlled by incubation with buffer and omitting the antiserum. No specific staining was observed after this procedure.
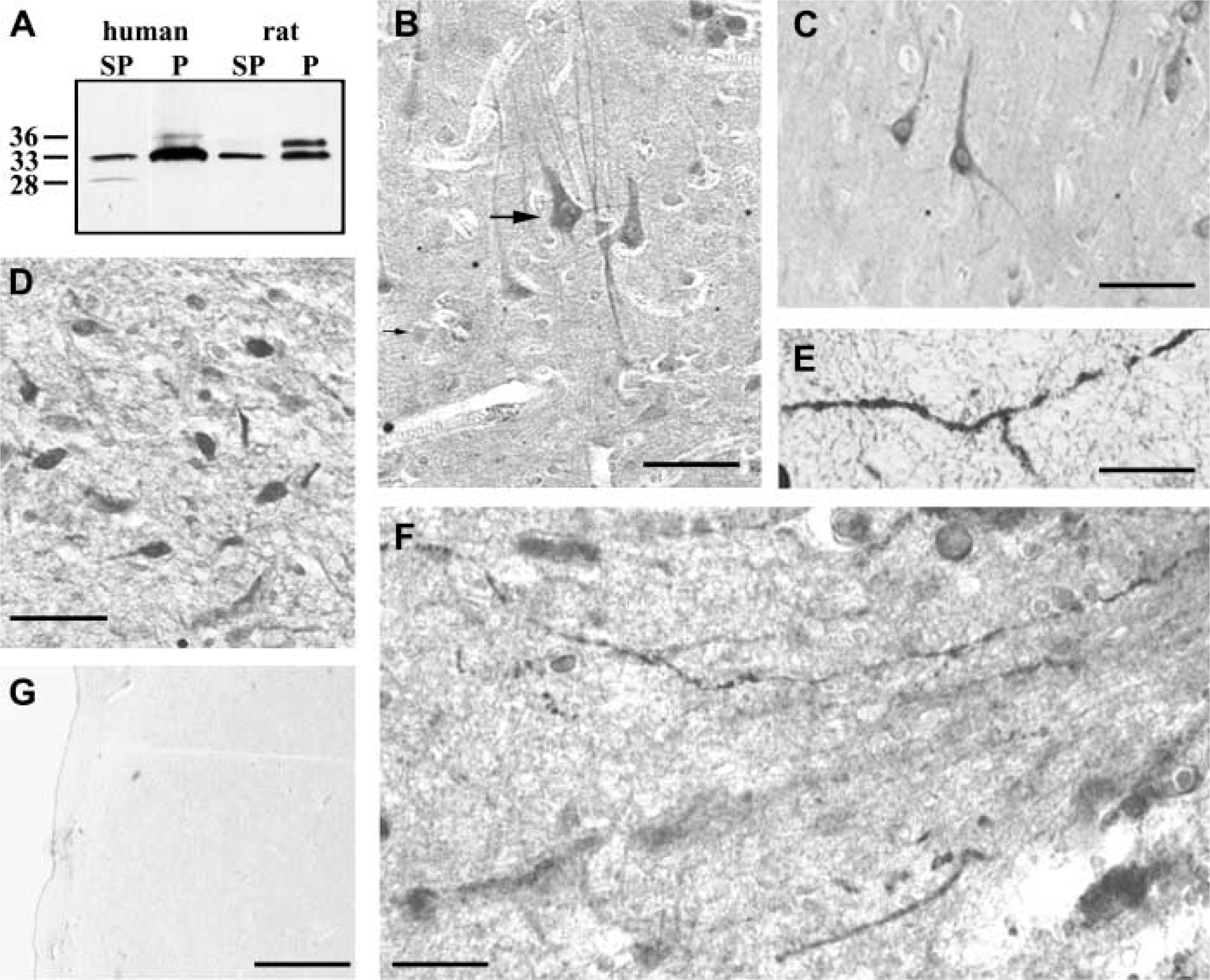
Comparison of the rat and human caldendrin cDNAs revealed a high degree of conservation among species (Laube et al. 2002). Accordingly, caldendrin-IR bands are of identical size and comparable abundance in human and rat specimens, as the Western blot in Figure 1A demonstrates. Furthermore, the partitioning of caldendrin isoforms at 33 and 36 kD into soluble and insoluble fractions is observed in the same way in both species. However, the phosphorylated 33-kD isoform (Seidenbecher et al. 1998) is more abundant in the human particulate fraction. A smaller soluble isoform at 28 kD, which is known to result from alternative N-terminal splicing of the caldendrin gene, is below the detection limit in rat cortex but is found in human material (Figure 1A).
Caldendrin showed a widespread but uneven distribution in human forebrain. No apparent gender differences in caldendrin's regional and cellular localization were found in the three brains examined. To determine to what extent postmortem time affects staining, we chose brains that were processed after different postmortem delays. There were no differences with regard to intracellular staining intensity and/or regional staining patterns between a brain with a short-term (12 hr, male brain) and two others with a long-term postmortem interval (39 and 48 hr, male and female brains).
Fourteen different paleo- and neocortical areas were studied: gyrus semilunaris, gyrus ambiens, gyrus parahippocampalis, gyrus entorhinalis, gyrus occipitotemporalis lateralis, gyri temporales inferior and medius, gyri longus and breves insulae, gyrus precentralis, gyrus postcentralis, gyri frontales medius and superior, and gyrus cinguli. Caldendrin-IR neurons were found in all cortical areas under investigation. Typically, layer III and V pyramidal neurons stood out by intense immunostaining (Figure 1B and 1C). Intracellularly, the immunoreaction was located in the perikarya and the dendrites of the nerve cells, with a staining pattern that closely resembled those of the dendritic marker MAP2 (not shown). The strongest intracellular staining intensity was observed in cingulate cortex pyramids (Figure 1C). Sometimes the neuropil showed weak to moderate immunostaining to caldendrin. A variety of interneurons were also found to express the protein. The weakest reactions were seen in entorhinal, parahippocampal, and temporal neurons.
The vast majority of pyramidal neurons showed faint to moderate immunostaining (not shown). Dentate gyrus granule cells were devoid of reaction product. Hippocampal interneurons were not immunoreactive for caldendrin. For the amygdaloid complex, the presence of some scattered neurons expressing the protein was evident (not shown).
Both basal ganglia and thalamus were characterized by multiple faintly stained neurons (not shown). No particular nuclei were caldendrin-IR.
In addition to cortical areas, the hypothalamus was the brain region with the most prominent immunolocalization of caldendrin. The protein was localized in neurons of different hypothalamic nuclei. The most pronounced immunostaining appeared in suprachiasmatic neurons near the III ventricle (Figure 1D). Remarkable immunostaining was confined to certain neurons of the paraventricular, the supraoptic (Figure 1E), the lateral, and the arcuate nuclei (not shown). Interestingly, a few strongly stained axons were detected within neurosecretory nuclei (Figure 1F) near the hypothalamic stalk (Figure 1G). This observation was somewhat surprising because it is at variance with results obtained in all other investigated rat and human brain regions. Preliminary data in rat, however, support the presence of caldendrin-IR in neurohypophyseal axons (Landwehr et al. unpublished observations). It is noteworthy that the antisera employed in this study also detects two shorter splice isoforms of caldendrin that are virtually absent from all other rat brain structures (Laube et al. 2002). The expression of caldendrin isoforms in rat hypothalamus, however, has not been analyzed to date. Therefore, we are now testing whether the present observation is due to a selective transport of caldendrin in neurophyophyseal axons and whether this reflects a differential and specific splicing event producing axonal caldendrin isoforms in hypothalamic neurons.
In human forebrain, however, the distribution of caldendrin-IR was very similar to the somato-dendritic localization of the protein in rat brain. Interestingly, as in rat always only a subpopulation of cells was caldendrin-IR. With respect to the almost identical expression pattern of caldendrin-IR, one can therefore conclude that cellular functions of caldendrin in rat and human are most likely similar. In addition, the almost identical distribution at the light microscopic level suggests that caldendrin expression in a subset of mainly principal and few interneurons has evolved due to differential requirements for somato-dendritic Ca2+ signaling in these cells. Recent functional studies have linked caldendrin signaling to voltage-dependent Ca2+ channels (Lee et al. 2002) and inositol trisphosphate receptor Ca2+ release channels (Yang et al. 2002). In addition, recent experiments from our laboratory (unpublished observations) suggest that caldendrin's protein interactions differ from those of CaM. The present work substantiates the notion that these interactions take place in the same rat and human cell types and brain regions.
Footnotes
Acknowledgements
Supported by the DFG (Kr1879/2-1, 2–2), the Fonds of the Chemische Industrie, the Land Sachsen-Anhalt (FKZ: 2508A/0086 and 3004A/0088H), and the Fritz Thyssen Stiftung.
The excellent technical assistance of S. Funke and H. Dobrowolny is gratefully acknowledged.