
Abstract
Cyclin T1 was recently identified, together with cdk9 (previously named PI-TALRE), as part of the TAK multiprotein complex, a co-factor targeted by the human immunodeficiency virus Type 1 (HIV-1) protein named Tat, suggesting a role for this complex in transcription elongation. Although studies on mRNA and protein expression have shown that cyclin T1 is ubiquitous in adult human tissues, no data have yet been reported regarding the expression of this protein in different cell lineages. Using a polyclonal antiserum raised against cyclin T1, we investigated the pattern of expression of this protein in adult human tissues by immunohistochemistry. Cyclin T1 was expressed ubiquitously, although different levels of expression were found in various organs. Some specialized tissues, such as blood, lymphoid tissues, and cells of connective tissue origin, showed high cyclin T1 expression. These specific expression patterns are only partially justified by some well-known specialized functions of cyclin T1 in certain cell types, such as its involvement in peripheral blood lymphocytes and monocyte differentiation. The high expression level found in other tissues suggests new possible roles for cyclin T1 in cell types other than those of lymphoid tissue.
Precise regulation of the cell cycle is a fundamental requirement for homeostasis of the eukaryotic cell. During the past decade, scientists successfully delved into the molecular machinery devoted to the fine regulation of the cell cycle phases and identified and characterized several genes and gene products involved. A key role in this is played by cell cycle kinases (cdk), relatively small proteins with an apparent molecular mass between 33 and 43 kD whose activity is regulated by the arrangement in a multimeric complex with larger proteins, called “cyclins” after their cyclical expression and degradation during the cell cycle. Cyclins, a family of proteins, are the regulatory subunits of the multimeric complex whose catalytic activity is provided by the cdk moiety. Different cdk/cyclin complexes, formed with clear-cut timing throughout the cell cycle, together with their phosphorylation/dephosphorylation efficiently regulate the activity of the multimeric holoenzyme. Conversely, cdk/cyclin complexes are negatively modulated by the binding of a family of small proteins called cdk inhibitors (MacLachlan et al. 1995; Peng et al. 1998). Cdk/cyclin complexes also are involved in processes clearly distinguishable from proliferative functions, such as differentiation and apoptosis (Lew et al. 1994; Tsai et al. 1994; Bagella et al. 1998). Cdk7 and cdk9 have been involved in the phosphorylation of the C-terminal domain of the RNA polymerase II, thus enhancing polymerase processing along the DNA template in a non-cell cycle-dependent manner (Bieniasz et al. 1999). Moreover, it has been found that, in mouse embryo development, cdk9 activity increases on terminal differentiation of skeletal muscles and neuronal cells in a fashion closely resembling that of cdk5 (Bagella et al. 1998).
Recent identification of cdk9 in the TAK (Tat-associated kinase) multiprotein complex, a co-factor targeted by the human immunodeficiency virus Type 1 (HIV-1) protein named Tat, suggests a role for this protein in controlling transcription elongation (for review see Yankulov and Bentley 1998). Anti-cdk9 antibodies deplete TAK activity from nuclear extracts of HeLa cells, thus indicating that cdk9 is an essential component of TAK (Gold et al. 1998; Ping and Rana 1999). The interaction between Tat and cdk9 was demonstrated not to be direct. In fact, identification of a cyclin-binding partner for cdk9, called cyclin T (after Tat), provided the link between these two proteins (Wei et al. 1998).
Human cyclin T has an amino-terminal cyclin box motif sharing 39% identity with the human cyclin and a carboxy-terminal PEST sequence (aa 709–726), which is frequently found in G1 cyclins to regulate their turnover by cellular ubiquitination and proteolysis pathways (Wei et al. 1998). Cyclin T, recently renamed cyclin T1, is the predominant cdk9-associated cyclin, although cdk9 also is found in complexes with an alternative spliced form, cyclin T2 (Peng et al. 1998; Wimmer et al. 1999). The cyclin T1 gene has been mapped to human chromosome 12 (Wei et al. 1998). Northen blotting analysis reveals a major cyclin T transcript (~8 kb) widely expressed in human tissues. A longer transcript, ~9.5 kb, has been detected in peripheral blood lymphocytes (PBLs) and a shorter transcript, ~3.0 kb, is found exclusively in the testis. In addition an abundant 3.5-kb transcript has been found only in the ovary, using a cDNA probe corresponding to the C-terminal region of the cyclin (Peng et al. 1998; Ping et al. 1998). These results indicate that cyclin T1 mRNA is expressed in all tissues and may be alternatively spliced in PBLs as well as in human germ line cells (Wei et al. 1998).
Studies on cyclin T1 mRNA and protein suggest its ubiquitous expression in human tissues. However, no data are yet available about the expression of the cyclin T1 gene product in particular cell types. We developed and characterized a specific polyclonal antiserum to describe the cyclin T1 immunohistochemical pattern of expression in several human tissues.
Materials and Methods
Normal Tissues and Cell Lines
Normal tissues from autopsy were obtained from the Department of Surgical Pathology of the Second University of Naples, Italy. Tissues were formalin-fixed and paraffin-embedded. Representative sections of each specimen were stained with hematoxylin–eosin and were examined by a pathologist to confirm the histological preservation of the microanatomic structure. For each tissue examined, at least three specimens from two different individuals were analyzed.
Human tumor cell lines SAOS-2 (osteosarcoma), T98G (malignant glioma), and MCF-7 (breast cancer) and mouse NIH-3T3 cells were obtained from the American Type Culture Collection (Manassas, VA) and were maintained in culture in Dulbecco's modified Eagle's medium (DMEM) complemented with 10% fetal calf serum at 37C in a 5% CO2-containing atmosphere. Human HaCaT immortalized keratinocytes (a gift from Dr. A. Venuti; Istituto Regina Elena, Rome, Italy) were cultured and were induced to differentiate by Ca++ addition as previously reported (Boukamp et al. 1988)
Antisera
The rabbit polyclonal immune serum against cyclin T1 was produced by immunizing rabbits with a bacterially expressed glutathione S-transferase (GST)–cyclin T1 fusion protein. Expression and purification of the fusion protein were done as previously reported (Frangioni and Neel 1993).
The production of the antiserum against cyclin T2a is described in the accompanying report (De Luca et al. 2001).
In Vitro Transcription–Translation and Immunoprecipitation Assay
One microgram of supercoiled plasmid encoding for human cyclin T1 and cyclin T2a were used to program a TnT rabbit reticulocyte lysate (Promega; Madison, WI) under control of the T7 polymerase in the presence of [35S]-methionine. Immunoprecipitation was carried out as described previously (De Luca et al. 1997).
Western Blotting
Western blotting on cell lysates or cell fractions was performed as described previously (Bagella et al. 1998). The anti-cyclin T1 antiserum was used at a 1:500 dilution. HSP70 was detected using a mouse monoclonal antibody (HSP01; Oncogene Science; Cambridge, MA).
Immunohistochemistry
Immunohistochemistry was carried out essentially as described previously (De Luca et al. 1997). Briefly, sections from each specimen were cut at 5–7 μm, mounted on glass, and dried overnight at 37C. All sections were then deparaffinized in xylene, rehydrated through a graded alcohol series, and washed in PBS. PBS was used for all subsequent washes and for antiserum dilution. Tissue sections were quenched sequentially in 0.5% hydrogen peroxide and blocked with diluted 10% normal goat anti-rabbit serum (Vector Laboratories; Burlingame, CA). Slides then were incubated at room temperature (RT) for 1 hr with the polyclonal immune serum raised against cyclin T1 at a 1:1000 dilution and then with diluted goat anti-rabbit biotinylated antibody (Vector Laboratories) for 30 min. All the slides then were processed by the ABC method (Vector Laboratories) for 30 min at RT. Diaminobenzidine was used as the final chromogen and hematoxylin was used as the nuclear counterstain. Negative controls for each tissue section were prepared by substituting the primary antiserum with the respective pre-immune serum. All samples were processed under the same conditions. Two pathologists (A. Baldi and F. Baldi) evaluated the staining pattern of the proteins separately and scored each specimen for the percentage of positive nuclei identified. An average of 50 fields was observed for each specimen. Cells were scored as reported in the legend to Table 1.
Cyclin T1 protein expression in normal adult human tissues a


a+ –, low to undetectable expression level (0–10% of positive cells); +, low expression level (11–20% of positive cells); ++, medium expression level (21–60% of positive cells); +++, high expression level (>60% of positive cells).
Results
Characterization of a New Specific Antiserum for Cyclin T1
A polyclonal antiserum was produced by immunizing rabbits with bacterially-expressed GST–cyclin T1. The specificity of this antiserum was assessed by immunoprecipitation. As shown in Figure 1A, the antiserum recognized a specific band with an apparent molecular mass of 87 kD in 35S-labeled SAOS-2 and T98G cell lysates (Figure 1, Lanes a and c). Preimmune serum from the same rabbit was used as a control (Figure 1, Lanes b and d). To prove that the antiserum was specific only for cyclin T1, the in vitro translated products of both cyclin T1 (Figure 1, Lane a) and cyclin T2a (Figure 1, Lane b) were western blotted and assayed with the antiserum anti-cyclin T1. As shown in Figure 1B, no cross-reaction was observed. An identical specificity was found for the antiserum against cyclin T2a, as shown in the accompanying article (De Luca et al. 2001).
For Western blotting analysis, protein extracts from MCF-7, SAOS-2, and T98G cells were used. Cyclin T1 expression was easily detectable in all these human cell lines by use of the specific antiserum (Figure 2). By means of a binding assay utilizing the in vitro-translated forms of the cyclin T family members, a negligible crossreactivity with cyclin T2a was observed. In addition, immune crossreactivity with murine cyclin T1 was demonstrated positively using a NIH-3T3 cell lysate (not shown).
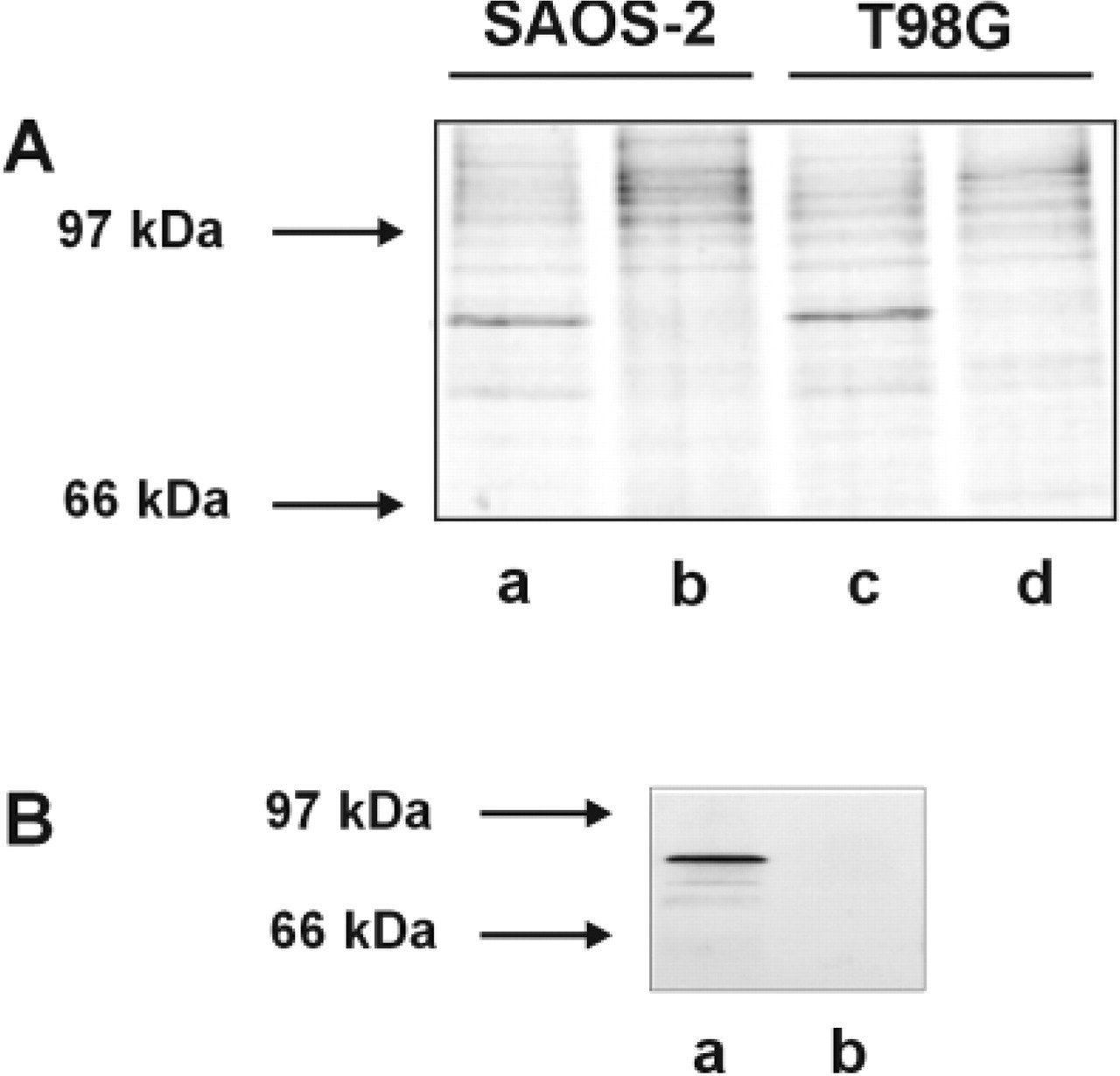
Characterization of a new polyclonal antiserum specific for cyclin T1. (
Expression Pattern of Cyclin T1 in Normal Human Tissues
We used immunohistochemistry to determine the amount and the localization of cyclin T1 in a panel of different human tissues. Specimens from different individuals were analyzed for each of the tissues examined. Cyclin T1 was expressed widely, although a different tissue distribution and/or level of expression was detected in the different organs examined (Table 1). Interestingly, the expression pattern of cyclin T2a, another partner of cdk9, almost completely overlapped the pattern described for cyclin T1 (De Luca et al. 2001).
Expression of Cyclin T1 in Epithelia
In general, epithelial cells, from either simple or stratified epithelium, showed positive staining for cyclin T1. Interestingly, stratified epithelia, such as epidermis and mucosa of the oral cavity, esophagus, cervix, and vagina, expressed a higher level of this protein in the immature basal and suprabasal layers (Figure 3A). A very similar pattern of expression was found for cyclin T2a (Figure 3C). Figures 3B and 3D show the negative controls, performed by substituting the primary antisera with the respective preimmune sera. In the skin, a low expression level for cyclin T1 was found in hair follicles, sebaceous glands, and sweat glands. Strong immunoreactivity was detected in the stratified columnar epithelia of trachea, bronchi, and adjacent glands (Figure 3E, arrow 1). Pneumocytes displayed a temperate nuclear immunoreactivity for the protein. A comparable pattern of expression was found for cyclin T2a (Figure 3F). Glandular breast epithelium showed intermediate levels of expression for cyclin T1.
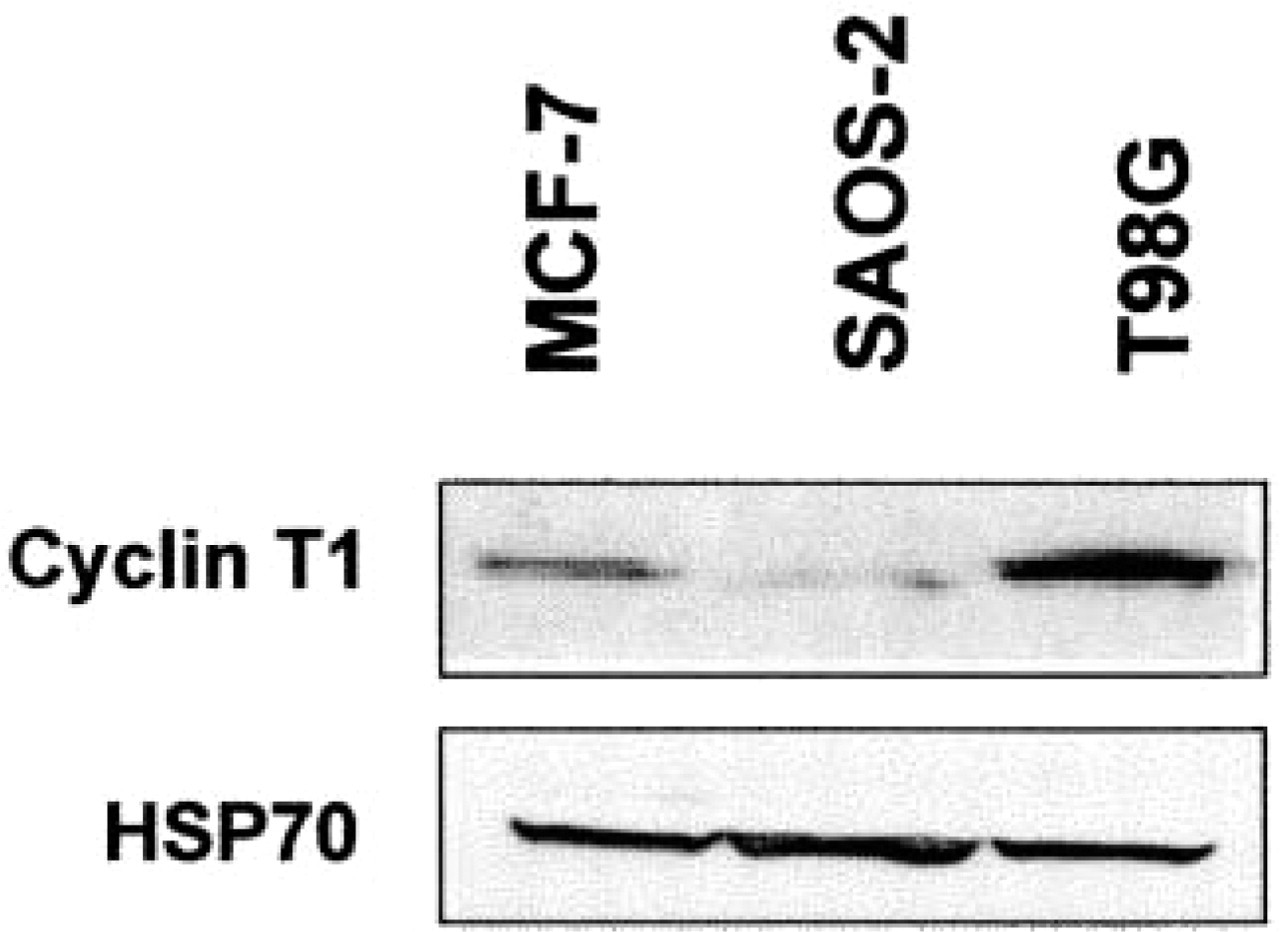
Characterization of a new polyclonal antiserum specific for cyclin T1. Western blotting analysis, showing expression of cyclin T1 in MCF-7, SAOS-2, and T98G cell lysates (40 μg). HSP70 was used for normalization of the protein amounts loaded.
In the gastrointestinal system, medium to low positive nuclear staining for cyclin T1 was found in salivary glands, in the epithelium of the stomach, and in the gallbladder epithelia. A low level of immunoreactivity was found in small and large intestine, esophagus, and liver, with the exception of ductal biliary cells, which displayed a medium level of cyclin T1 expression (Figure 3I). An analogous expression was found for cyclin T2a (Figure 3J). The esocrine portion of the pancreas showed a low level of expression, whereas the exocrine portion displayed a medium level of expression for cyclin T1.
In the urinary system, cyclin T1 was expressed at a low level in all kidney tubules, in the uroepithelium, and in the prostate, whereas the glomeruli showed an intermediate expression level for cyclin T1.
Cyclin T1 was expressed at a low level in thyroid, pituitary, hypophysis, and adrenal glands, where staining was evident both in the chromaffin and in the cortical portion.
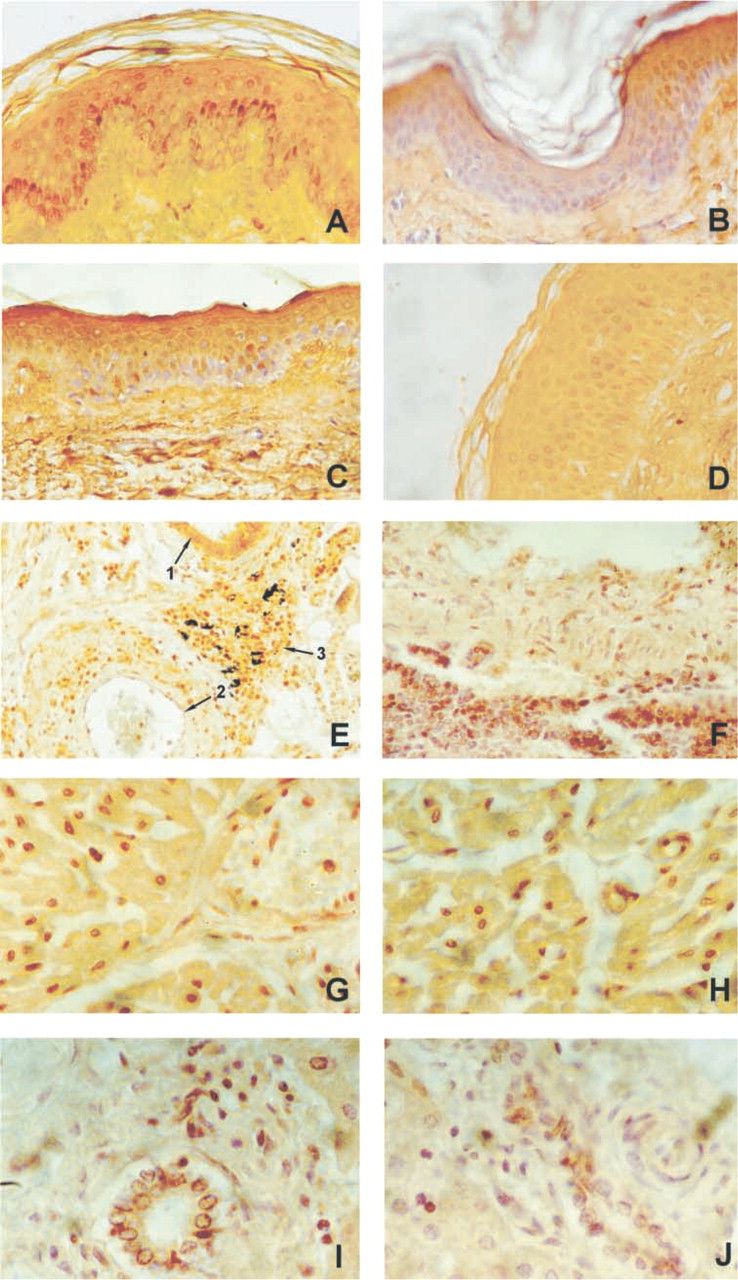
Expression pattern of cyclin T1 in human tissues. (
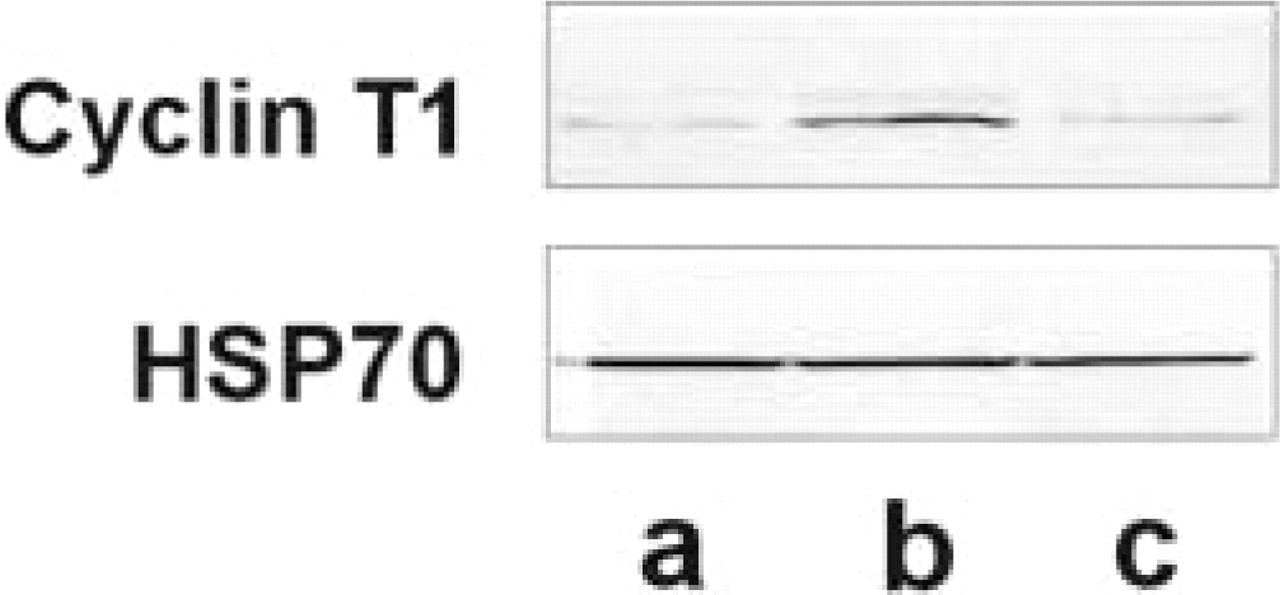
Expression of cyclin T1 in several human tissues revealed by Western blotting analysis. A representative panel including skin (Lane a), skeletal muscle (Lane b), and large intestine (Lane c) is shown. HSP70 was used for normalization of the protein amounts loaded.
In the reproductive system, cyclin T1 was expressed at a very low level in all organs studied.
Expression of Cyclin T1 in Cardiovascular and Connective Tissue
Cyclin T1 showed a peculiar expression pattern in mesenchymal cells. High immunoreactivity for cyclin T1 was observed in skeletal muscle cells and in myocardial cells (Figure 3G) in all specimens examined. An identical phenomenon was observed for cyclin T2a (Figure 3H). A high level also was found in adipocytes, chondrocytes, and endothelial cells (Figure 3E, arrow 2), whereas fibroblasts and smooth muscle cells displayed a very low level of expression for cyclin T1.
Expression of Cyclin T1 in Central and Peripheral Nervous Systems
Neurons from different areas of the brain, such as frontal cortex and midbrain, cells of the granular level of the cerebellum, and Purkinje cells exhibited an intermediate nuclear staining. Perineural and endoneural cells of peripheral nerves and ganglion cells showed low to undetectable levels of the protein. However, astrocytes, oligodendroglial, and microglial cells of brain tissue had a high level of expression of cyclin T1.
Expression of Cyclin T1 in Hematopoieic Cells
A high level of cyclin T1 was observed in T-cells and B-cells, both in blood films and in lymphoid tissues (Figure 3E, arrows 2 and 3), whereas monocytes, eosinophils, neutrophils, and basophils showed medium expression for cyclin T1.
To confirm by an alternative approach the expression pattern of cyclin T1, we performed Western blotting analysis on several human fresh tissues. In Figure 4 a representative panel, including skin (Figure 4, Lane a), skeletal muscle (Figure 4, Lane b) and large intestine (Figure 4, Lane c), is shown. These results were consistent with the immunohistochemical pattern of expression and were in good agreement with the Northern blotting analyses described previously (Peng et al. 1998; Ping et al. 1998).
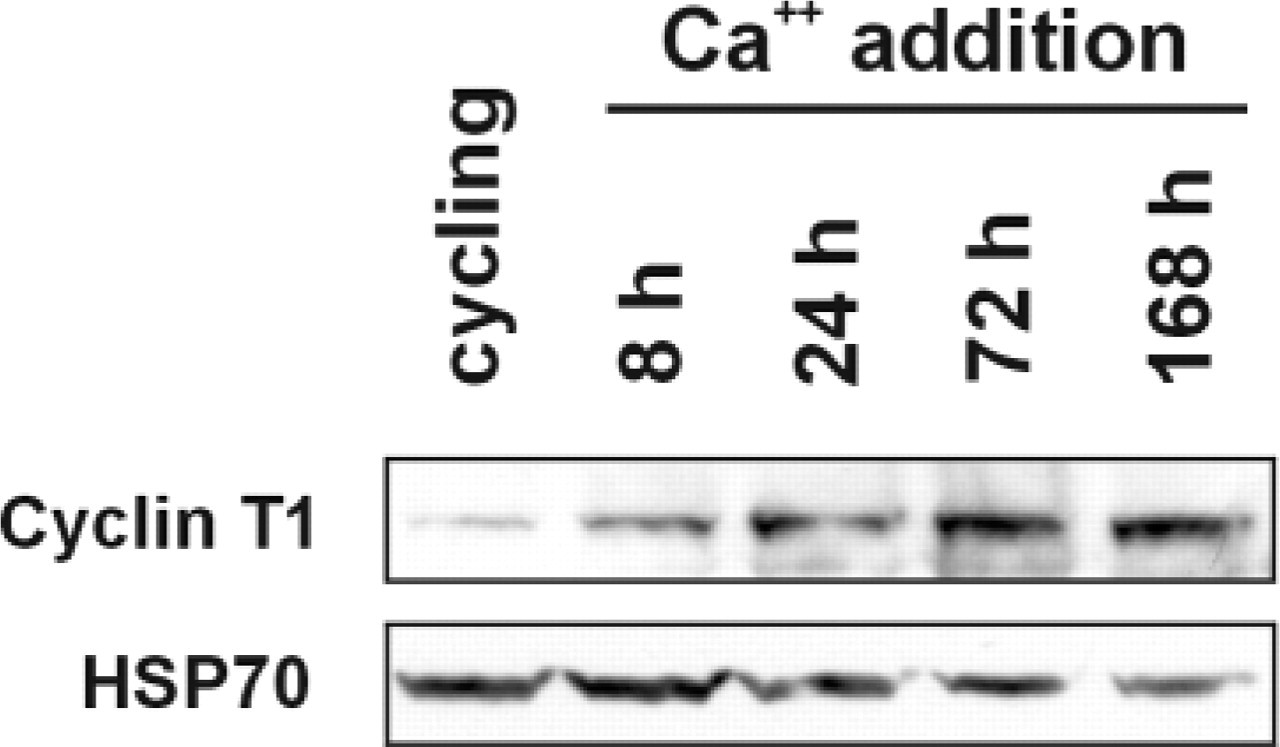
Modulation of cyclin T1 expression in differentiating HaCaT cells. Western blotting analysis, showing expression of cyclin T1 in cycling and in differentiating HaCaT cells. Differentiation was accomplished by the addition of Ca++ to the culture medium and samples were collected at the times indicated. HSP70 was used for normalization of the protein amounts loaded.
Expression of Cyclin T1 in Differentiating Immortalized Keratinocytes (HaCaT Cells)
After Ca++ addition to the culture medium, HaCaT cells undergo modifications that recapitulate the keratinocyte differentiation program (Boukamp et al. 1988). It should be noted that, in spite of normal skin keratinocytes, HaCaT cells do not undergo apoptosis at the end of the differentiation process. The degree of differentiation they reach in vitro is comparable to that of the suprabasal keratinocyte layer in skin sections (Chaturvedi et al. 1999). Cyclin T1 expression in differentiating HaCaT cells increased early after Ca++ addition and remained substantially elevated up until late differentiation stages (Figure 5).
Discussion
Biochemical and immunoistochemical characterization of a new polyclonal antiserum raised against cyclin T1 are described in this study. By taking advantage of a large human tissue bank, the pattern of expression of cyclin T1 in normal human tissues was analyzed. It is important to note that the antiserum assayed against the denatured products of cyclin T1 and cyclin T2a was specific only for cyclin T1. For that reason, it can be assumed that the immunohistochemical results are specific for cyclin T1.
After examination of the immunohistochemical pattern of expression of cyclin T1 in several human tissues, it can be concluded that cyclin T1 has a widespread pattern of expression, which is consistent with previous data on the RNA and protein expression profile for this protein in human tissues (Wei et al. 1998). Moreover, a very similar pattern of expression was found for cyclin T2a (see also De Luca et al. 2001). However, a different tissue distribution and/or level of expression was detected in several organs. For example, all epithelial cells showed unambiguous immunoreactivity for cyclin T1. The expression level in all stratified epithelia was clearly more intense in the immature basal portion of the epithelia, to which actively dividing cells are confined. There is an apparent discrepancy between the immunohistochemical data on cyclin T1 expression in skin sections and those obtained by Western blotting in the immortalized HaCaT keratinocyte cell line, where cyclin T1 expression continues to be elevated at late differentiation stages. This can be explained in part by the extensive physiological apoptosis detectable in terminally differentiated keratinocytes in skin sections, and in part by the degree of differentiation reachable in vitro by the HaCaT cell line (Chaturvedi et al. 1999). Similar discrepancies also have been detected when the analyses of negative regulators of the cell cycle, either in immortalized keratinocytes or in skin sections, were compared (Paramio et al. 1998).
Certain tissues, such as those of the endocrine and reproductive systems, showed low cyclin T1 expression. On the other hand, tissues of mesenchymal origin, such as connective tissue, skeletal muscle, and myocardial cells, adipocytes, chondrocytes, and endothelial cells, as well as blood and lymphoid tissues, exhibited high cyclin T1 expression levels. In the nervous system, the highest level of expression was found in oligodendroglia and microglia.
This peculiar pattern of expression might account for the distinct activities of this protein in selected tissues. It has recently been demonstrated that, in some cell lineages, the expression level of cyclin T1 is not growth- and/or cell cycle-regulated. There is an exception in regard to peripheral blood lymphocytes, in which expression levels and activity of the cyclin T1/cdk9 complex are upregulated during lymphoid cell activation, suggesting a role for the cyclin T1/cdk9 complex in T-cell activation and in monocyte differentiation (Yang et al. 1997; Garriga et al. 1998; Herrmann et al. 1998). The high level of expression for cyclin T1 that we found in peripheral blood lymphocytes is in agreement with these data and correlates well with our previous observation on the peculiar high expression level of cdk9 in primary human lymphoid tissues (De Luca et al. 1997).
When cyclin T1 expression in human tissues was compared with that of cdk9 (De Luca et al. 1997), it appeared that although both proteins in several tissues showed a similar pattern of expression, significant differences were observed in some others. For example, the high expression level for cyclin T1 in tissues of connective origin contrasted with the low level of cdk9. This behavior will require further investigation to be completely understood, as was the case for the expression pattern in lymphoid tissues.
An exhaustive knowledge of the differential cell- and tissue-specific pattern of expression of cyclin T1 in normal human tissues is an essential requirement for critical evaluation of the exact role played by this protein in cell homeostasis. Moreover, the observation that regulation of cyclin T1 expression is dependent on tissue-specific signaling pathways (Garriga et al. 1998), further strengthens the value of these data and suggests possible new roles for cyclin T1 in tissues other than lymphoid.
Footnotes
Acknowledgements
Supported in part by AIRC and Ministero della Sanità grants to MGP, by an FIRC grant to ADL, and by NIH grants RO1 CA 60999-01A1 and PO1 NS 36466 to AG. AB is a recipient of an FIRC fellowship.
We thank Dr P.G. Natali (Regina Elena Institute; Rome, Italy) for useful suggestions and criticism, Dr A. Venuti (Regina Elena Institute; Rome, Italy) for the HaCaT cell line, and Dr J.J. Gartland (Thomas Jefferson University) for editing the manuscript.