
Abstract
Metastatic processes, including cell invasion, extracellular matrix degradation, and tissue remodeling, require cellular reorganization and proliferation. The cell signaling molecules required and the proteins involved in cell restructuring have not been completely elucidated. We have been studying the role of sphingolipids in normal cell activity and in several pathophysiological states. In this study we used immunohistochemistry to observe the presence of the two known subunits of serine palmitoyltransferase (SPT) in proliferating cells, in an in vitro model of wound repair, and in human malignant tissue. We report increased expression of the two subunits, SPT1 and SPT2, in the proliferating cells in these models. We also demonstrate a change in subcellular localization of the SPT subunits from predominantly cytosolic in quiescent cells to nuclear in proliferating cells. In addition, we observed SPT1 and SPT2 immunoreactivity in reactive stromal fibroblasts surrounding the carcinoma cells of some of the tumors. This enhanced SPT expression was absent in the stromal fibroblasts surrounding normal epithelial cells. Our results suggest a potential role for overexpression of SPT in the processes of cell metastasis.
Keywords
C
Serine palmitoyltransferase (SPT) is the initial and rate-limiting enzyme in the de novo sphingolipid biosynthesis cascade. At least two subunits are involved in the enzymatic activity associated with SPT, SPT1 and SPT2 (Nagiec et al. 1996; Hanada et al. 1997). This enzyme catalyzes the condensation of palmitoyl-CoA with serine to form 3-keto-sphinganine, which is further metabolized to generate ceramide (Figure 1). Ceramide can be acted on by a number of enzymes to generate molecules known to be important in both intra- and intercellular signaling. Ceramidase converts ceramide to sphingosine, a static or apoptotic signal in some cells. Sphingosine serves as the substrate for sphingosine kinase to generate sphingosine-1-phosphate (SPP), a known proliferative signal that can be secreted from cells (Spiegel et al. 1994). In addition, ceramide can be converted to glycosphingolipids, which are important in cell recognition, proliferation, and differentiation (Hannun and Bell 1989; Harel and Futerman 1993; Morton et al. 1994; Schwartz et al. 1995). Therefore, the regulation of SPT may have widespread implications for cell responses and pathologies because of its prominent position as the enzyme catalyzing the committed and rate-limiting step of the sphingolipid metabolic cascade.
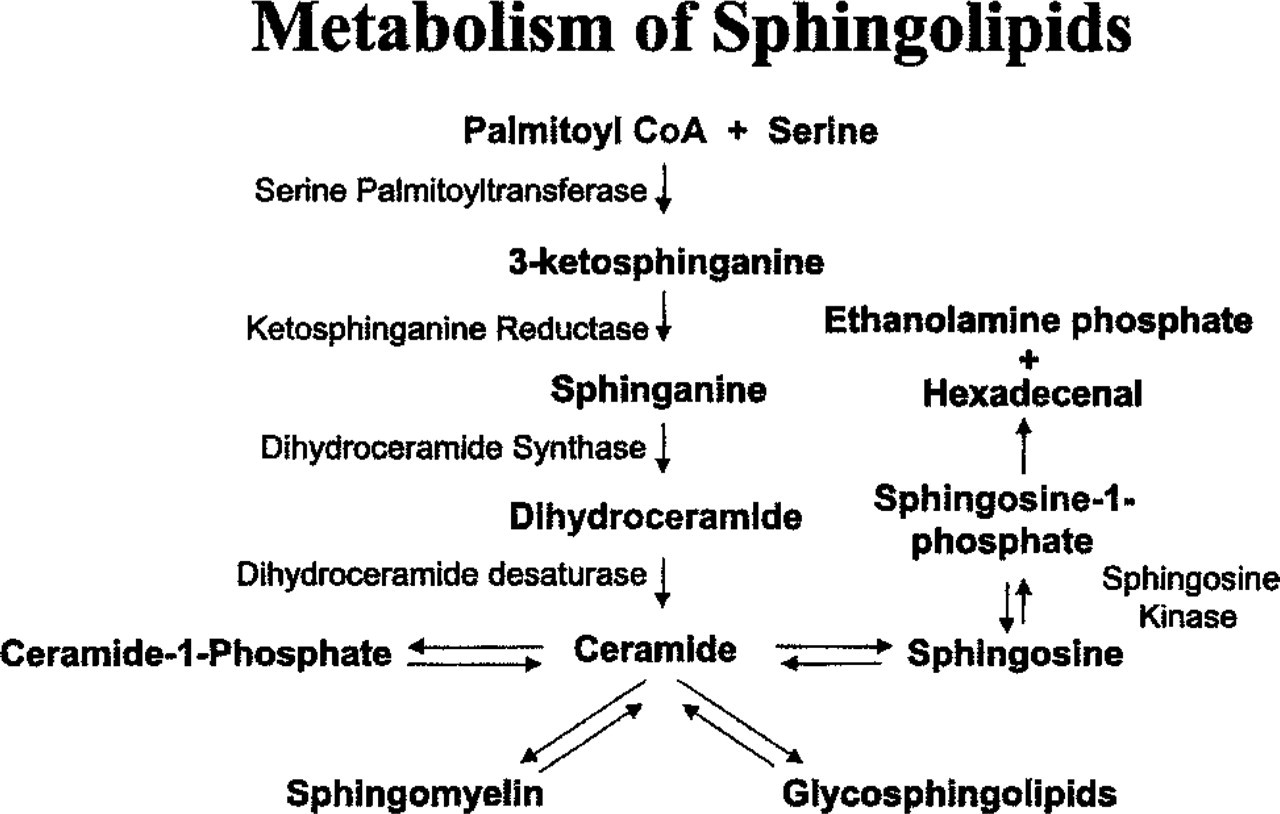
The de novo sphingolipid synthesis pathway.
Changes in lipid metabolic pathways are among the first events in the de-differentiation of normal cells (Sabine 1975). Studies have shown that a higher percentage of sphingolipids is present in the membranes of hepatomas compared to normal liver cells (Williams et al. 1984a,b). In this study we demonstrate that SPT is expressed abundantly in proliferating fibroblasts in culture and in the hosting “reactive” stromal fibroblasts surrounding the malignant cells in some tumors, which was not observed in the surrounding stromal fibroblasts in normal tissue. Prominent stro-mal reaction (desmoplasia) is seen in many invasive carcinomas, suggesting that stromal cells play a role in cancer pathogenesis (Gregoire and Lieubeau 1995). Reactive fibroblasts, also known as myofibroblasts, are frequently associated with different cancers of epithelial origin (Schmitt–Graff et al. 1994) and influence the invasive and metastatic potential of carcinoma cells (Gregoire and Lieubeau 1995). We demonstrate high expression of the SPT subunits in several established human tumor cell lines and in situ in human malignant cells. In addition to an increase in SPT expression, we also observed changes in the subcellular localization of SPT in proliferating fibroblasts and malignant cells.
The role of nuclear lipid metabolism in signal transduction cascades has recently become apparent. Diacylglycerol kinase, an enzyme involved in phospholipid metabolism, has been shown to localize to the nucleus and is involved in nuclear signal transduction (Bregoli et al. 2001). Sphingosine kinase (SPHK) has also been shown to localize to the nucleus of 3T3 cells within 24 hr of mitogenic stimulation, with an increase in nuclear SPHK activity after the treatment (Kleuser et al. 2001). Sphingolipid metabolites are present and active in nuclear preparations, lending support to the idea that sphingolipids play a regulatory role in mediating cell activities from within the nucleus (Albi and Magni 1997; Jaffrezou et al. 2001). Our work demonstrates the nuclear association of SPT in proliferating fibroblasts and malignant cells. This represents the second enzyme involved in sphingolipid metabolism that becomes associated with the nucleus on stimulation. These data suggest a role for sphingolipids as signaling molecules within the nucleus in addition to their previously reported activities in cytoplasmic signaling cascades and as intercellular signaling molecules.
Materials and Methods
Antibody Characterization and Western Blotting
Rabbit polyclonal antibodies to the SPT subunits SPT1 and SPT2 have been characterized and evaluated previously (Uhlinger et al. 2001; Batheja et al. 2003). Nuclear extracts were prepared from human neonatal dermal fibroblasts for Western blotting. Briefly, cells were lysed by hypotonic lysis and nuclei separated by centrifuging the extract at 14,000 rpm for 20 sec. The nuclear pellet was resuspended in PBS and homogenized. Fifty micrograms of nuclear extract were fractionated on a 4–12% SDS-polyacrylamide gel. After transfer to a nitrocellulose membrane, the membrane was probed with 2 μg/ml of SPT1- or SPT2-specific polyclonal serum. The antigen specificity of the polyclonal sera was determined by preincubating the sera with 20 μg/ml antigenic peptides overnight at 4C before probing the immuno-blots. Bound SPT1 antibody was detected with a 1:20,000 dilution of horseradish peroxidase-conjugated goat anti-rabbit IgG (Upstate Cell Signaling Solutions; Waltham, MA) and the CDP-star chemiluminiscence kit (Perkin Elmer; Boston, MA). Bound SPT2 antibody was detected using a 1:10,000 dilution of an alkaline phosphatase conjugated goat anti-rabbit IgG (Santa Cruz Technology; Santa Cruz, CA) and the CDP-star chemiluminiscence kit (Perkin Elmer).
Immunohistochemistry (IHC)
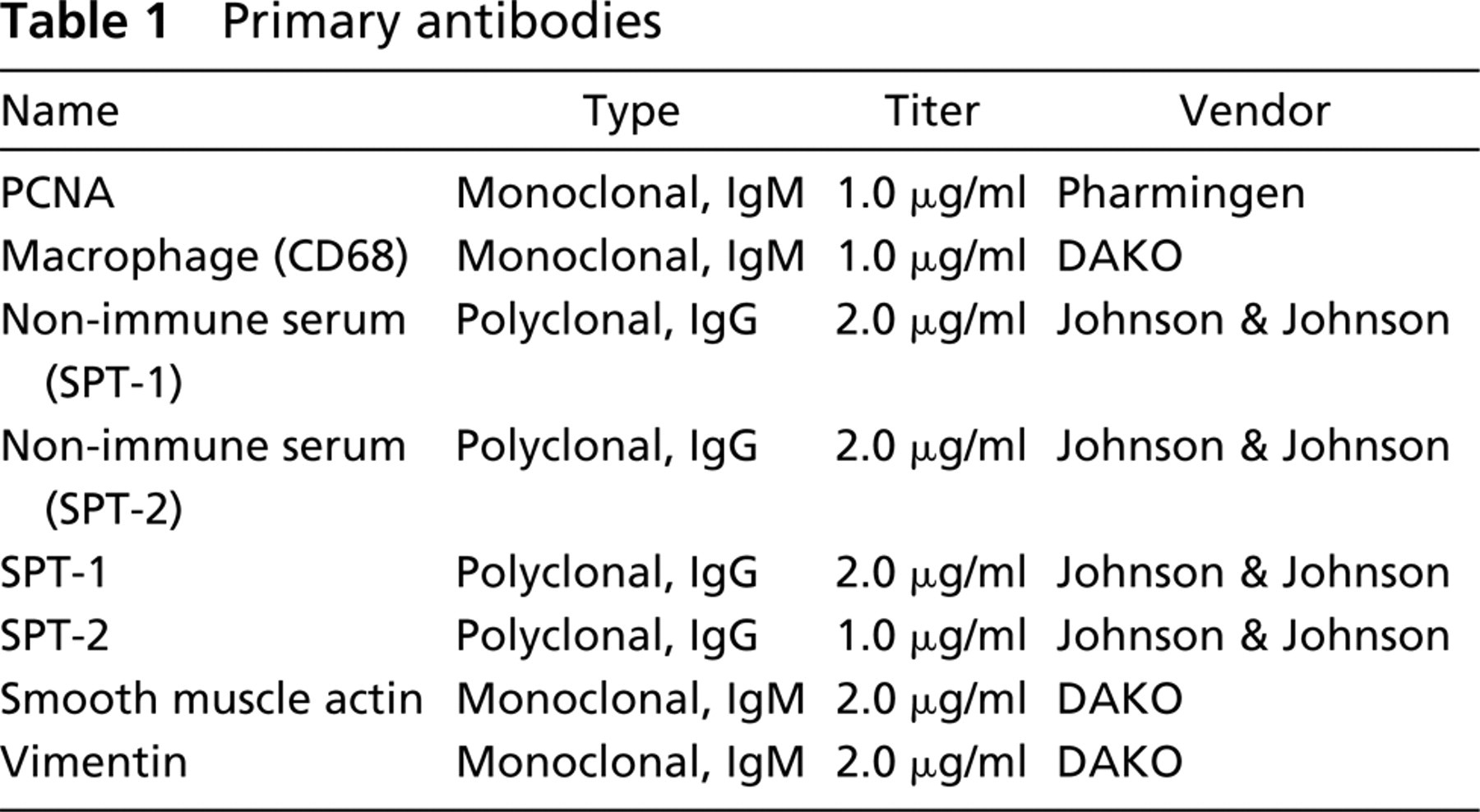
Table 1 lists the titer and source of the positive and negative control antibodies and the experimental antibodies used in these studies. Commercial human normal and tumor checkerboard tissue slides (DAKO, Carpenteria, CA; Biomeda, Foster City, CA) were deparaffinized, hydrated, and processed for routine IHC, which has previously been described (D'Andrea et al. 1998). Briefly, slides were microwaved in Target (DAKO), cooled, placed in PBS pH 7.4, then placed in 3.0% H2O2. Slides were processed through an avidin–biotin blocking system according to the manufacturer's instructions (Vector Labs; Burlingame, CA) and then placed in PBS. All reagent incubations and washes were performed at room temperature. Normal blocking serum (Vector Labs) was placed on all slides for 10 min. After brief rinsing in PBS, primary antibodies (Table 1) were placed on slides for 30 min. The slides were washed and biotinylated secondary antibodies, goat anti-rabbit (polyclonal antibodies) or horse anti-mouse (monoclonal antibodies) placed on the tissue sections for 30 min (Vector Labs). After rinsing in PBS, the avidin–horseradish peroxidase-biotin complex reagent (ABC; Vector Labs) was added for 30 min. Slides were washed and treated with the chromogen 3,3'-diaminobenzidine (DAB; Biomeda), rinsed in dH2O, and counterstained with hematoxylin. Monoclonal antibodies to vimentin (Table 1) were used to demonstrate tissue antigenicity and reagent quality. The negative controls included replacement of the primary antibody with the same-species IgG isotype non-immune serum (Table 1). In addition, we preabsorbed the antibodies with their specific antigen overnight in a 10-fold titer excess of antigen as another negative control. The specificity of the SPT1 and SPT2 antibodies in IHC has also been shown previously (Batheja et al. 2003).
Primary antibodies
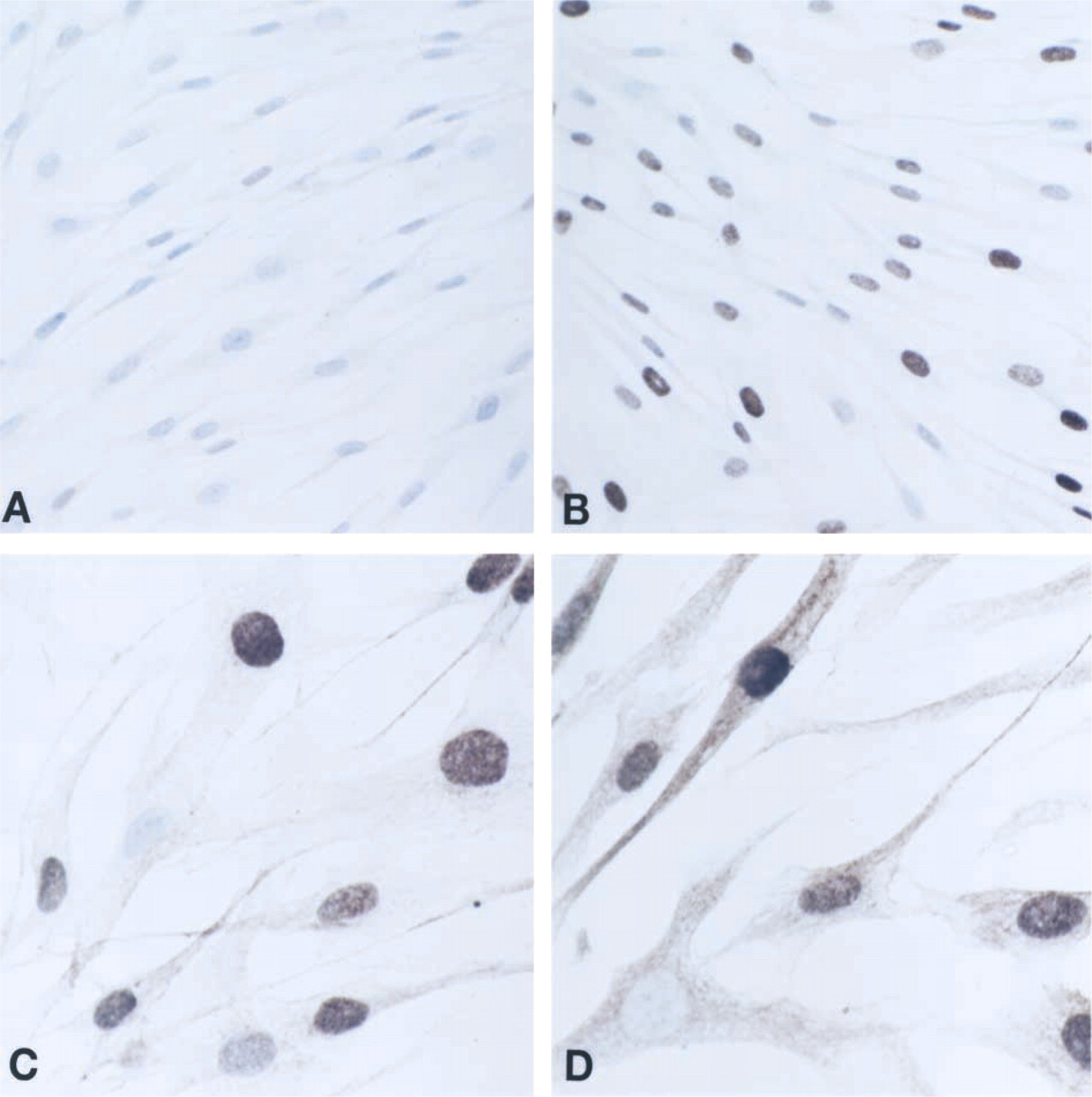
Immunocytochemical analysis of SPT1 and SPT2 expression in subconfluent fibroblasts. Negative control labeling (
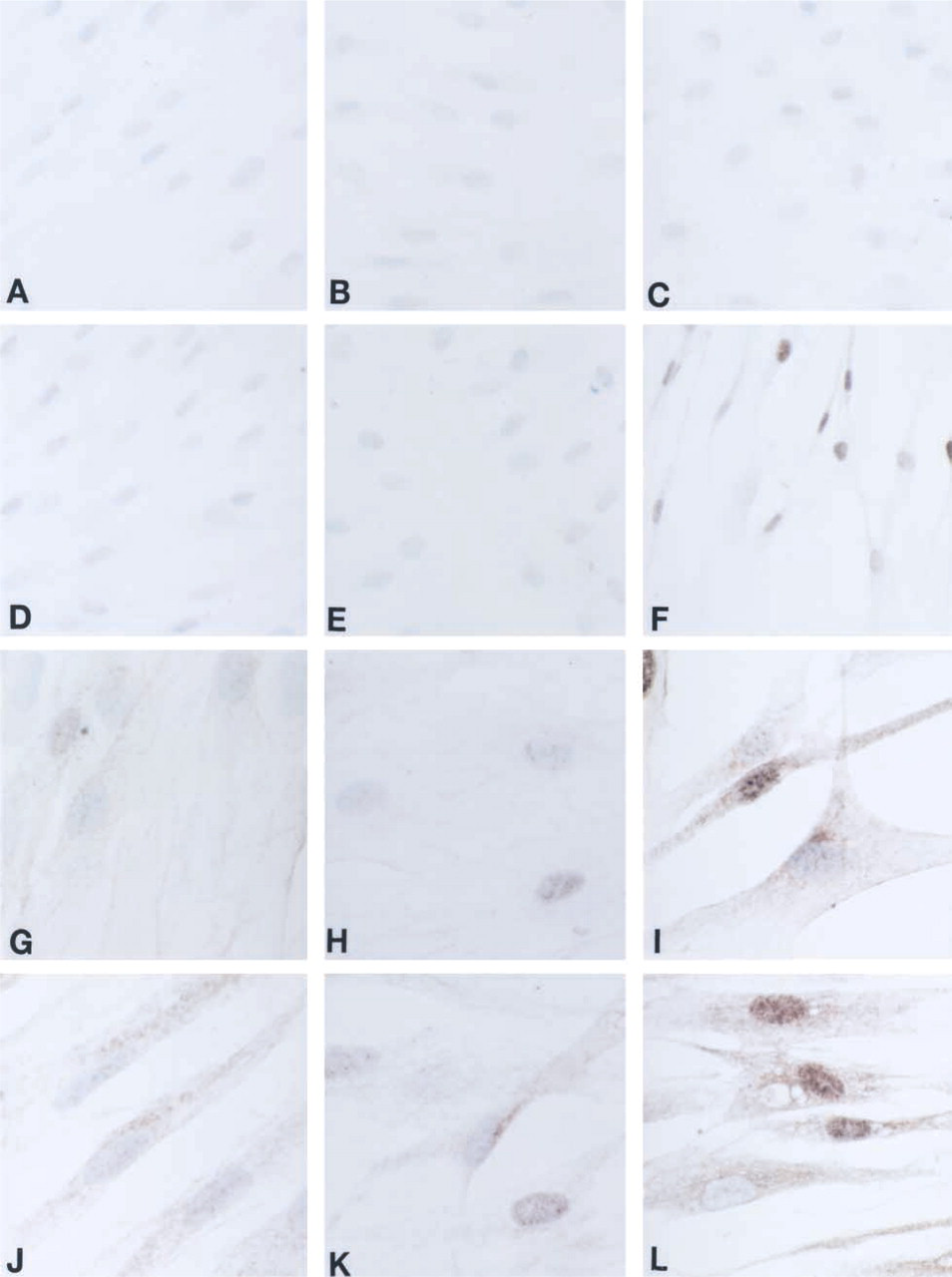
Immunocytochemical analysis of SPT1 and SPT2 expression in quiescent and wounded fibroblasts. Nine-day quiescent fibroblasts (
Cell Culture
Human neonatal dermal fibroblasts and their culture media were obtained from Clonetics/BioWhittaker (Walkersville, MD). Cell suspensions (5 × 104/ml) were seeded in four-well chamber slides (NUNC; Naperville, IL) for immunocy-tochemistry. To mimic the in vivo activation of differentiated quiescent fibroblasts in vitro, a scrape-wounding model was used as described previously (D'Andrea et al. 2001). Briefly, cells were incubated for either 2 days (subconfluent, proliferative conditions) or 9 days (hyperconfluent, quiescent conditions), or a 9-day quiescent culture was subjected to mechanical scraping using the end of a pipette. To assess expression in the wound response, the cells were cultured for an additional 5 days after wounding.
Immunocytochemistry (ICC)
Four-chambered cultured slides were fixed with 10% neutral buffered saline for 10 min at RT, rinsed in PBS, and then assayed for ICC as previously described (Smith–Swintosky et al. 1997; Nierodzik and Chen 1998). All washing steps were performed using automation buffer with Tween-20 (Research Genetics; Huntsville, AL).
Double Immunohistochemistry (IHC:IHC)
To determine if the SPT subunits co-localized in proliferating cells, we used double immunohistochemical methods (IHC:IHC) to simultaneously detect SPT1 or SPT2 expression with detection of a proliferation marker, proliferating cell nuclear antigen (PCNA), using protocols described previously (D'Andrea et al. 1999). Briefly, slides were first processed for single IHC labeling protocols for detection of SPT1 or SPT2 as described above. Without processing the slides for hematoxylin, PCNA antibodies (Pharmingen; San Diego, CA) were placed on the tissues for 30 min. After several PBS washes, the biotinylated horse anti-mouse secondary antibodies (Vector Labs) were similarly incubated. The presence of PCNA-positive cells was visualized using an alkaline phosphatase detection system through incubation with alkaline phosphatase-conjugated ABC (Vector Labs) followed by development using the Fast Red chromogen (Sigma; St. Louis, MO). Slides were then routinely counter-stained and mounted.
Results
SPT catalyzes the initial and rate-limiting step in de novo sphingolipid synthesis, as shown in Figure 1. This biosynthetic pathway generates ceramide, which serves as a second messenger signaling molecule and as the precursor molecule for complex sphingolipids.
To characterize the expression and localization of SPT1 and SPT2 in cells and tissues by ICC and IHC, antibodies specific to SPT1 and SPT2 (Batheja et al. 2003) were used. The positive and negative controls used in these studies in addition to the experimental antibodies are listed in Table 1. The negative controls did not produce observable labeling.
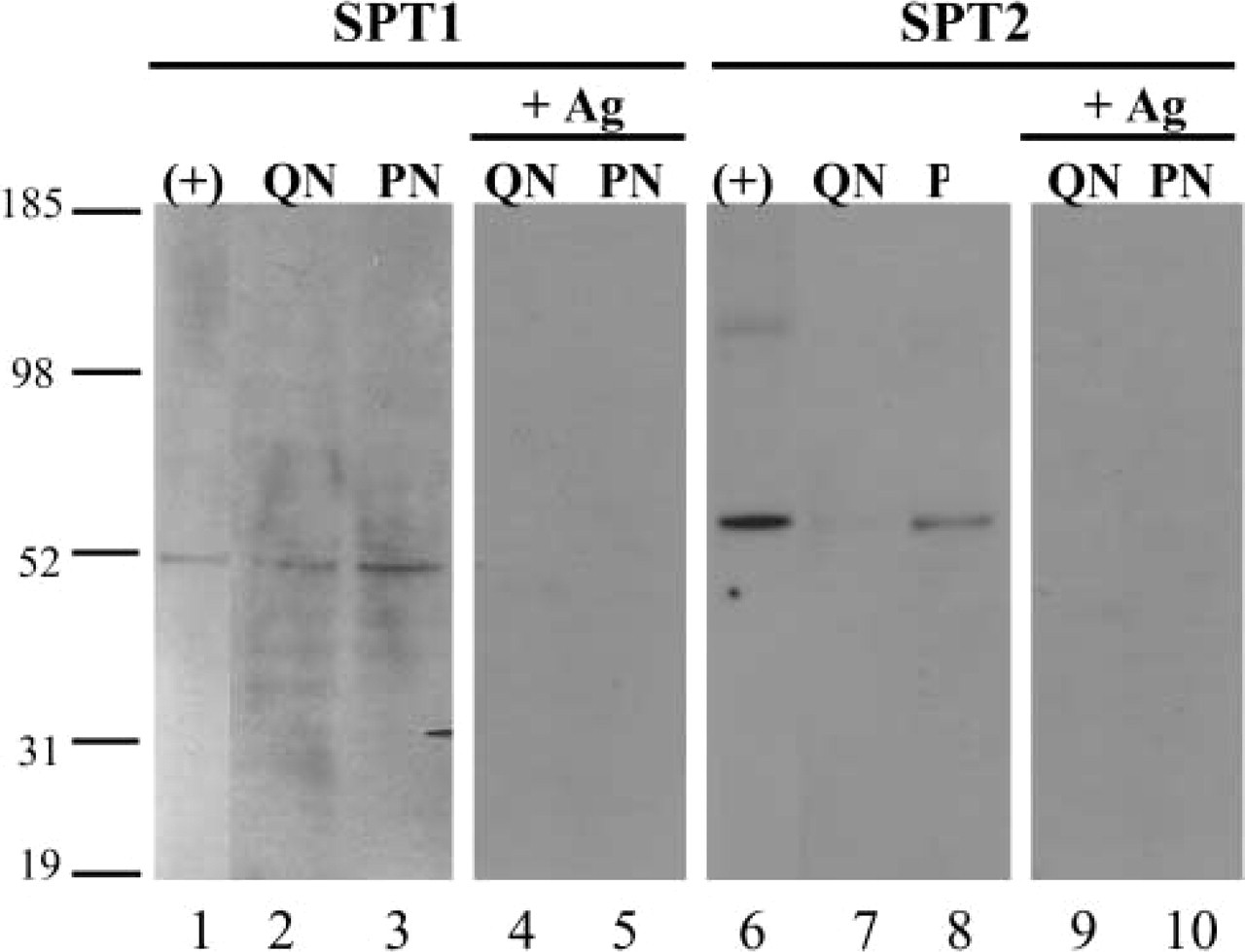
Western blots of quiescent and proliferating neonatal fibroblasts. Nuclear extracts from quiescent (QN) and proliferating (PN) human neonatal dermal fibroblasts were fractionated on a 4–12% SDS-PAGE. Recombinant human SPT1 (Lane 1) and SPT2 (Lane 6) produced with a baculovirus expression system were included as positive controls for the experiment. Strips were probed with either anti-SPT1 (Lanes 1–5) or anti-SPT2 (Lanes 6–10) antibody. Competition with antigenic peptides used to raise the antibodies completely inhibited SPT1 (Lanes 4 and 5) and SPT2 (Lanes 9 and 10) detection. +Ag, competing antigen.
Subconfluent human dermal fibroblasts were immunolabeled by ICC to observe the expression and localization of SPT1 and SPT2 in the cells. Figure 2 shows the results of the negative control labeling (Figure 2A), PCNA labeling (Figure 2B), which appears as nucleus-associated labeling in the proliferating cells, SPT1 (Figure 2C) and SPT2 (Figure 2D) labeling. Interestingly, although some diffuse staining was observed throughout the fibroblasts, the majority of the labeling by the anti-SPT antibodies appeared to be associated with the nucleus in these proliferating fibroblasts. There were, however, nuclei that were not stained by PCNA, SPT1, or SPT2 antibodies.
An in vitro model was used to mimic the in vivo activation by tissue wounding of differentiated quiescent fibroblasts to further characterize the expression of SPT in proliferating cells. Quiescent fibroblast cultures were compared to confluent cultures subjected to mechanical scraping and allowed to recover for 5 days (wound conditions). Figure 3 shows no labeling by ICC in the 9-day quiescent cultures (Figures 3A, 3D, 3G, and 3J) or the 14-day quiescent cultures (Figures 3B, 3E, 3H, and 3K) immunolabeled with the negative control (Figures 3A and 3B) and PCNA (Figures 3D and 3F) antibodies. Light diffuse labeling was observed with the SPT1 (Figures 3G and 3H) and SPT2 (Figures 3J and 3K) antibodies. In contrast, the 14-day wounded fibroblasts (Figures 3C, 3F, 3I, and 3L) showed strong immunolabeling for PCNA (Figure 3F), SPT1 (Figure 3I), and SPT2 (Figure 3L), and no staining in the negative control (Figure 3C). The most dramatic observation was the intense nuclear localization of SPT1 and SPT2 in the 14-day wounded fibroblasts (Figures 3I and 3L) in comparison to the 9 (Figures 3G and 3J)- and 14 (Figures 3H and 3K)-day quiescent cells.
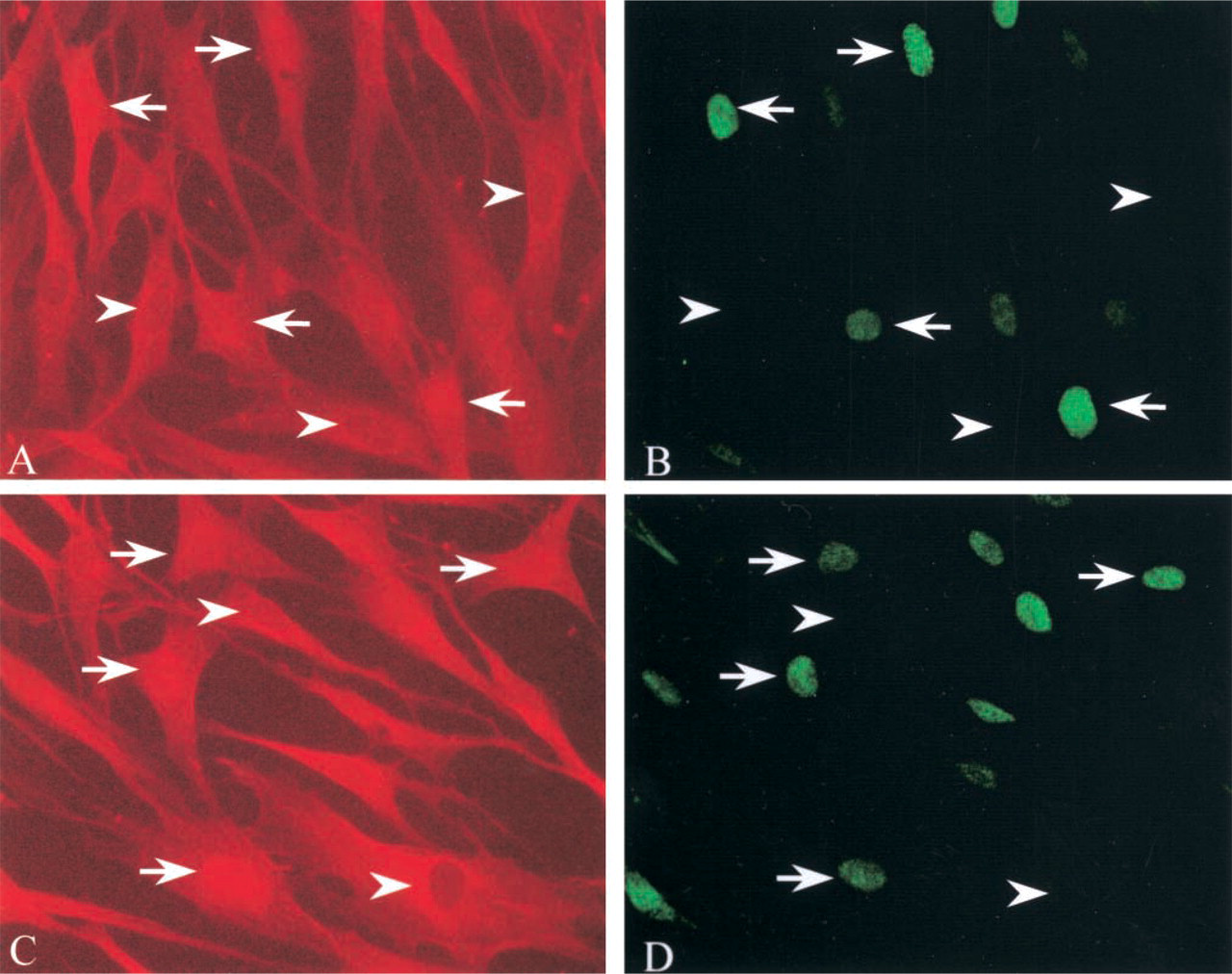
Double staining (IF:IF) of fibroblasts for PCNA and SPT1 or SPT2. Proliferating fibroblasts were stained for PCNA (
To demonstrate an increase in SPT1 and SPT2 proteins in the nuclei of proliferating neonatal fibroblasts, Western blots were performed with equal concentrations of nuclear extracts from quiescent and proliferating dermal fibroblasts, respectively (Figure 4) SPT1 and SPT2 were more abundant in the nuclear protein extract from proliferating cells than that from quiescent cells. The increase in SPT2 in proliferating cells compared with quiescent cells was more dramatic than that observed for SPT1. In competition assays performed simultaneously, the presence of antigenic peptides used for generation of the SPT1 and SPT2 antibodies completely inhibited SPT1 (Figure 4, Lanes 4 and 5) and SPT2 (Figure 4, Lanes 9 and 10) detection.
Immunocytochemical analysis of SPT expression in human tumor cell lines. Jurkat cells (
Immunolocalization of SPT1 and SPT2 in human malignant tissues a
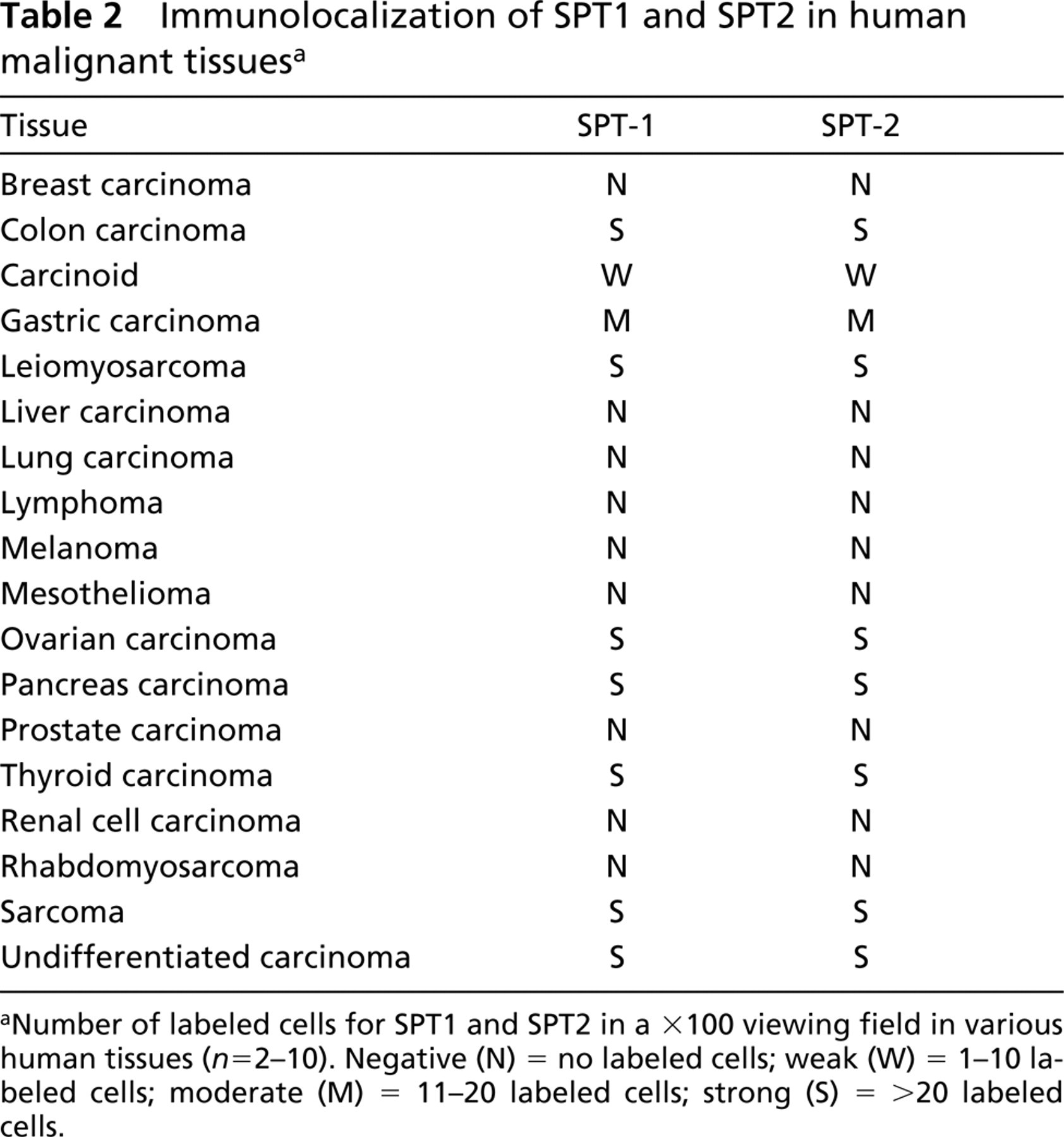
aNumber of labeled cells for SPT1 and SPT2 in a X100 viewing field in various human tissues (n = 2–10). Negative (N) = no labeled cells; weak (W) = 1–10 labeled cells; moderate (M) = 11–20 labeled cells; strong (S) = >20 labeled cells.
To investigate the lack of staining seen in some nuclei (Figures 2 and 3), double staining (IF:IF) (Figure 5) was used to show coincidence of PCNA (Figures 5B and 5D) and increased SPT1 (Figure 5A) and SPT2 (Figure 5C) labeling. Arrows indicate cells in which PCNA was detected, and the SPT labeling appeared to be strong and associated with the nucleus. Arrowheads indicate cells lacking PCNA-label and with diffuse SPT labeling that did not show specific association with the nucleus. From the expression patterns observed in this study and the previously described in vitro wounding model, it appears that SPT expression is increased and nucleus-associated in proliferating cells. In addition to the model of wounding described above, we have recently reported the increased expression of SPT1 and SPT2 in de-differentiated fibroblasts and proliferating vascular smooth muscle cells in balloon-injured rat carotid artery (Uhlinger et al. 2001).
Parallels between wound repair responses and tumor formation have recently become even more apparent (Gospodarowicz 1976; Van Waes 1995; Frazier and Grotendorst 1997). Because we observed increased expression of SPT1 and SPT2 in both in vitro and in vivo wound repair models, as described above, we examined several cancer cell lines for over-expression of SPT1 and SPT2 using ICC. As shown in Figure 6, there was intense SPT immunolabeling in three well-established cancer cell lines, Jurkat (Figures 6A-6C), HT-29 (Figures 6D-6F), and SH-SY 5Y (Figures 6G-6I). We also observed that the distribution of the SPT subunits in the proliferating tumor cell lines was different. Whereas the SPT1 staining was primarily diffuse, the SPT2 staining was mostly localized to the nucleus. Differences between intracellular distribution of SPT1 and SPT2 subunits have also been reported previously (Batheja et al. 2003). Irrespective of the staining pattern, all cells stained strongly positive for SPT. This intriguing observation led us to explore the expression of SPT in human tumors.
Table 2 shows the immunolocalization of SPT1 and SPT2 in a variety of human malignancies. Positive labeling was defined by the presentation of brown staining and was scored according to the number of labeled cells for SPT1 and SPT2 in a X100 viewing field in 18 different human tumors with an average of five fields of each tissue: negative (N) = no labeled cells; weak (W) = 1–10 labeled cells; moderate (M) = 11–20 labeled cells; strong (S) = >20 labeled cells. Five carcinomas (colon, ovarian, pancreas, thyroid, undifferentiated) and two sarcomas demonstrated strong overexpression of both SPT subunits. Other tumors (Table 2) expressed little (W) or no (N) detectable levels of SPT1 or SPT2.
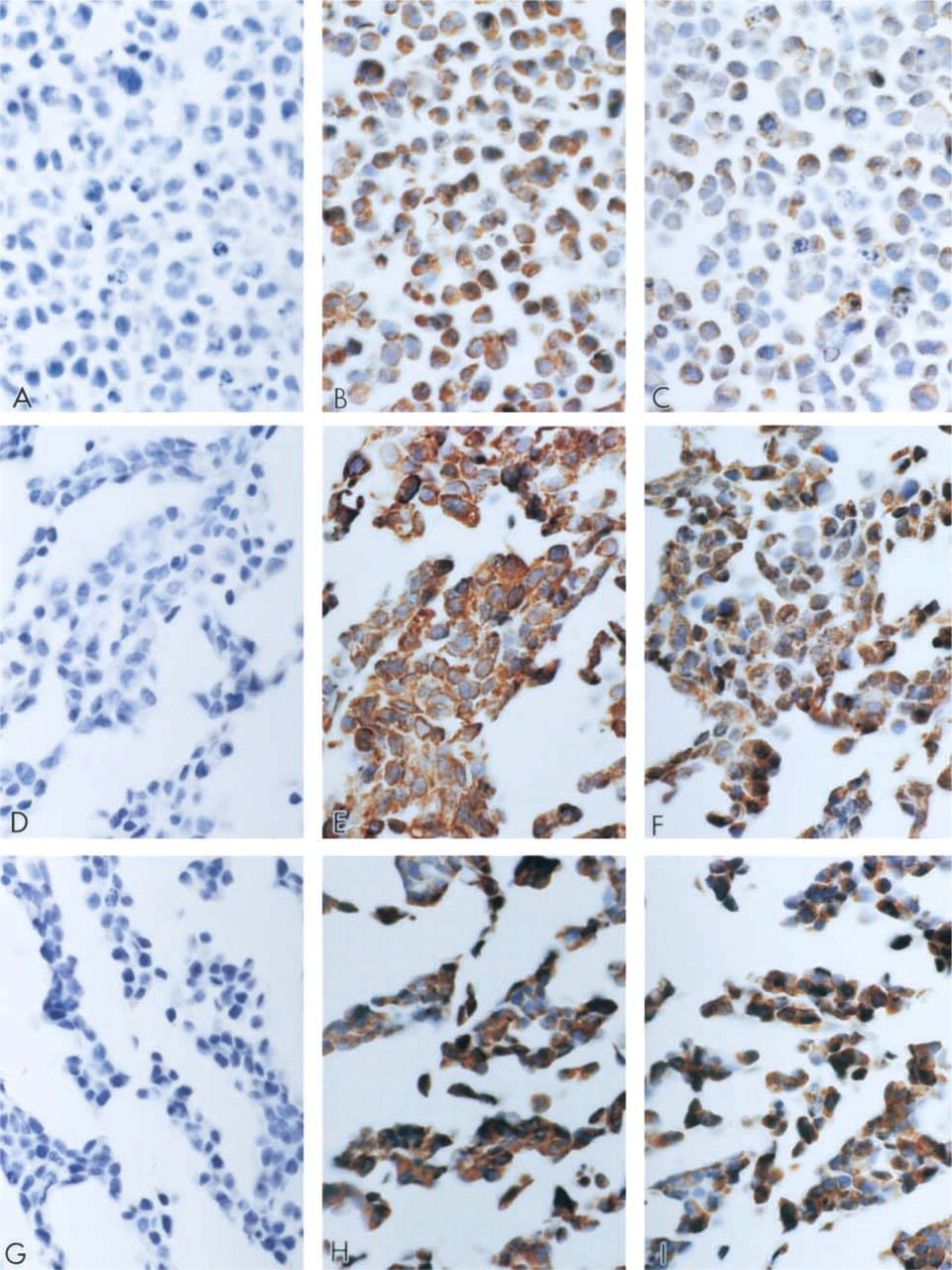
Human malignant colon carcinoma tissues were processed for IHC using antibodies to the preimmune serum (Figure 7A), SPT1 (Figure 7B), and SPT2 (Figure 7C). Figure 7 also shows strong intracellular labeling of SPT1 (small arrowheads) and SPT2 (small arrowheads) in the malignant cells. In addition to the labeling observed in the malignant cells, immunostaining was observed in the stromal fibroblasts adjacent to the tumor (large arrowheads). Human malignant undifferentiated carcinoma tissues (Figures 7D-7F) were processed for IHC using antibodies to the preimmune serum (Figure 7D), SPT1 (Figure 7E), and SPT2 (Figure 7F). There appeared to be a stronger SPT1 signal (arrowheads, Figure 7E) in the undifferentiated human carcinoma tissue relative to the weak SPT2 signal (Figure 7F). These slides were processed simultaneously in a multi-tissue format. Therefore, comparisons of the intensity of the signal among the tissue samples may be relevant. Because the role played by the two SPT subunits in the enzymatic activity, whether catalytic or regulatory, is still unclear, it is not evident whether this enhanced SPT1 expression, or even the ratio of SPT1 to SPT2 expression, is critical to a cell response.
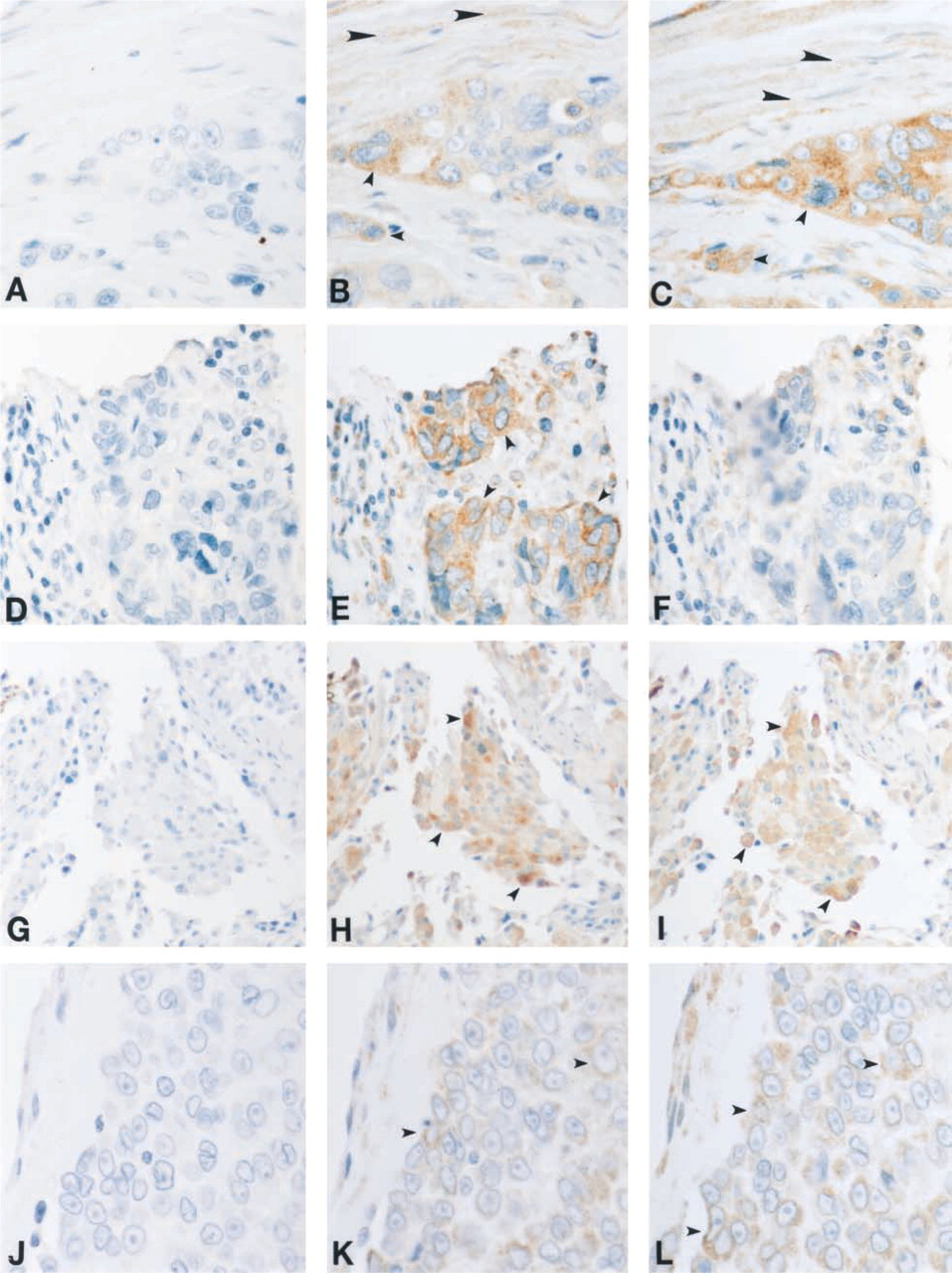
SPT expression in colon carcinoma (
Figures 7G–7I show the observations made in human malignant thyroid carcinoma tissues processed using the preimmune serum (Figure 7G), SPT1 (Figure 7H) and SPT2 (Figure 7I) antibodies. There appeared to be SPT-specific staining in the majority of the cells of this tumor. The intensity of the SPT1 and SPT2 signal (arrowheads) is variable from cell to cell, with some of the malignant cells expressing very high levels of the SPT subunits. Further studies are needed to determine if the various expression levels of these subunits indicate different requirements for the enzyme in tumor cells.
In addition to studying various carcinoma tissue samples, we also examined human malignant sarcoma tissues (Figure 7J–7L). We observed abundant expression of SPT1 and SPT2 (arrowheads) in this malignancy as well as in the carcinomas described above. These results indicate that the overexpression of SPT in cancer cells is not limited to those derived mostly from epithelial cells, such as carcinomas. Cancers such as sarcomas, which are derived from mesenchymal cells, also appear to have induced expression of the SPT subunits.
Discussion
We have developed antibodies to human SPT1 and SPT2, which have enabled us to investigate the expression of these enzyme subunits in normal human tissue (Batheja et al. 2003). Recently, we used these antibodies to demonstrate elevated levels of SPT1 and SPT2 in vascular smooth muscle cells and activated fibroblasts in balloon-injured rat carotid arteries (Uhlinger et al. 2001). In this study we present additional support for the increased expression of SPT subunits in the proliferating cells of wounded tissue. ICC on human dermal fibroblasts demonstrates increased expression of SPT1 and SPT2 in an in vitro wounding model. Quiescent, non-proliferating fibroblasts show light, diffuse SPT1 and SPT2 staining throughout the cells. However, proliferating cells responding to the mechanical scraping insult show a distinct upregulation of both SPT subunits, demonstrated by intense immunolabeling in these cells. In addition to the increase in expression, it is strikingly apparent that a significant amount of the increased SPT1 and SPT2 immunolabeling is associated with the nucleus. The function of SPT1 and SPT2 associating with the cell nucleus in de-differentiated, proliferating fibroblasts is not clear but is now under investigation. It is possible that the nuclear localization of SPT is involved in sphingolipid nuclear signaling (e.g., for mitogenesis). The observation that sphingosine kinase translocates to the nucleus in fibroblasts treated with PDGF supports the hypothesis of sphingolipid involvement in nuclear signaling in proliferating and transformed cells (Kleuser et al. 2001).
Because of the emerging paradigm that some of the key molecules involved in the cell wound repair response are also involved in tumor growth and metastasis, we performed a series of IHC studies on tumor cell lines and human tumor tissues to study SPT1 and SPT2 expression. The SPT subunits are abundantly expressed in several well-established human tumor cell lines, including a lymphoma, an adenocarcinoma, and a neuroblastoma cell line. We provide morphological evidence for increased expression of SPT1 and SPT2 in malignant carcinoma cells and in the cell types forming the tumor microenvironment (TME), such as the reactive stromal fibroblasts and local macrophages. SPT1 and SPT2 were not detected in stromal fibroblasts in similar normal tissues (Batheja et al. 2003). The elevated levels of the SPT subunits in these cell types suggest a role for SPT in cell metastatic activities and proliferation.
Our lab and others have observed enhanced expression of SPT in activated leukocytes involved in an inflammatory response (Memon et al. 1998). The upregulation of SPT in human macrophages has been demonstrated to increase flux through the sphingolipid biosynthetic pathway (Carton, unpublished). The increased expression of SPT in human malignancies may be indicative of regulation of flux through the sphingolipid biosynthetic pathway occurring in cells undergoing neoplastic transformation and in the cells of the surrounding TME. Studies are under way to determine if the upregulation of expression of SPT results in increased expression of sphingolipids that serve as signaling molecules for proliferation of these cells.
Sphingosine-1 phosphate (S-1-P), a sphingolipid that can be generated via the de novo biosynthetic pathway (Figure 1), has clearly been shown to act as a mitogen through intra- and intercellular signaling (Lee et al. 1998; Zhang et al. 1991). Elevated levels of S-1-P within a cell promote cell proliferation, and this activity appears to be through the dynamic balance between the levels of S-1-P and ceramide (Spiegel et al. 1998). Secreted S-1-P is a ligand for several endothelial differentiation gene receptors (EDG-R) specifically Edg-1, −3, −5, and −8 (Ancellin and Hla 1999), and recently Edg-6 (Van Brocklyn et al. 2000). Activation of the EDG receptors by S-1-P promotes a variety of cell responses, including proliferation, migration, adhesion molecule expression, and cytoskeletal changes in many cell types (Spiegel 2000). In particular, the binding of S-1-P by EDG-1 in endothelial cells stimulates angiogenesis (Lee et al. 1998; Wang et al. 1999). The role of S-1-P and its receptor in wound repair and tumors is now under investigation.
Other sphingolipids in addition to S-1-P that can be generated via the de novo synthesis pathway have been implicated in proliferative and metastatic processes. Changes in expression of various glycosphingolipids on the cell surface have been correlated with acquiring and maintaining cancer phenotypes, tumor progression, and metastasis (Hakomori 1981; Hannun and Bell 1989; Harel and Futerman 1993; Morton et al. 1994; Schwarz et al. 1995). For example, ganglioside GD2 levels increase as melanoma tumorigenesis progresses (Fukuda et al. 1986). GD2 has been implicated in the attachment of melanoma cells to solid substrates and may be involved in promoting tumor metastasis (Cheresh et al. 1984).
Elevated levels of glycosphingolipids in cell membranes may bring about changes in cell–cell interactions and could modulate receptor complex formation and signal transduction through membrane receptors. Studies are under way to determine if glycosphingolipid content is being modulated through regulation of de novo sphingolipid synthesis in malignant cells and in the cells that make up the TME. A number of studies have looked at sphingolipid metabolites, particularly ceramide, and their relationship to the effectiveness of various cancer therapeutics (Senchenkov et al. 2001). Many of these studies were performed on transformed cell lines subjected to pharmacological perturbation of the sphingolipid biosynthetic pathway (Wang et al. 2002). In the present study we have demonstrated the enhanced expression of SPT in proliferating fibroblasts, transformed cell lines, and in human tumors. The various sphingolipid metabolites, generated ultimately from SPT activity, elicit a multitude of effects, ranging from the induction of apoptosis to mitogenesis. The relevant metabolite in the systems we have examined remains unknown but is now under investigation. To our knowledge, this is the first in situ histological comparative report describing the expression of SPT1 and SPT2 protein in human malignant tumor cells, local macrophages, and in the “reactive” stromal fibroblasts surrounding the tumor cells. The elevated levels of this enzyme suggest possible mechanisms for some of the aberrant cell activities in the tumor and in the cell types that form the TME. The observed shift in subcellular localization of the SPT subunits in proliferating fibroblasts, from diffuse and cytoplasmic to nucleus-associated, suggests a functional role for this enzyme.
Additional studies will be needed to determine if SPT levels and cell location correlate with specific tumor types or if the relative amounts of SPT1 and SPT2 in the tumor cells and in the stromal fibroblasts are clinically relevant. It is possible that the amounts of SPT1 and SPT2 in the TME cells may be a valid predictor of metastatic activity, thereby imparting diagnostic and prognostic value. More importantly, these data suggest that serine palmitoyltransferase inhibitors may represent a novel class of compounds with therapeutic utility against certain neoplasias.
Footnotes
Acknowledgements
We express our gratitude for the excellent histological and immunohistochemical expertise of Patti A. Reiser, BS, MT, HT (ASCP), Norah A. Gumula, HT (ASCP), Brenda M. Hertzog, BS, MT (ASCP), Debbie A. Polkavitch, BS, MT (ASCP), Barbara Branchide, and Danielle Schmidheiser, BS, of the MorphoMetrics department.