
Abstract
Secretogranin III (SgIII) is one of the acidic secretory proteins, designated as granins, which are specifically expressed in neuronal and endocrine cells. To clarify its precise distribution in the anterior lobe of the rat pituitary gland, we raised a polyclonal antiserum against rat SgIII for immunocytochemical analyses. By immunohistochemistry using semithin sections, positive signals for SgIII were detected intensely in mammotropes and thyrotropes, moderately in gonadotropes and corticotropes, but not in somatotropes. The distribution pattern of SgIII in the pituitary gland was similar to that of chromogranin B (CgB), also of the granin protein family, suggesting that the expressions of these two granins are regulated by common mechanisms. The localization of SgIII in endocrine cells was confirmed by immunoelectron microscopy. In particular, secretory granules of mammotropes and thyrotropes were densely and preferentially co-labeled for SgIII and CgB in their periphery. Moreover, positive signals for SgIII were occasionally found in cells containing both prolactin and TSH in secretory granules. These lines of evidence suggest that SgIII and CgB are closely associated with the secretory granule membrane and that this membrane association might contribute to gathering and anchoring of other soluble constituents to the secretory granule membrane.
Keywords
A
In addition to these major members of the family, at least two other acidic secretory proteins, 7B2 and secretogranin III (SgIII), are also regarded as granins. 7B2 was originally identified as a pituitary-specific protein (Marcinkiewicz et al. 1987; Martens 1988; Seidah et al. 1983), and later it was revealed that 7B2 acts as a kind of molecular chaperone for the proprotein convertase 2 (PC2) in POMC-producing cells (Braks and Martens 1994). Accumulated evidence suggests that the 7B2 protein is involved in the transport and activation processes of PC2 by interacting with it inside the regulated secretory pathway (Westphal et al. 1999; for review see Mbikay et al. 2001).
In contrast to 7B2, biochemical properties and the tissue distribution of SgIII have not been fully elucidated. SgIII was first identified as a product of a neuron-specific gene 1B1075 of mouse and, on the basis of its biochemical properties, it was later renamed SgIII (Dopazo et al. 1993; Kingsley et al. 1990; Ottiger et al. 1990). The distribution of SgIII in the central and peripheral nervous systems has been examined in detail by immunocytochemical and in situ hybridization methods (Ottiger et al. 1990), showing that immunoreactivity of SgIII is present in the endocrine cells of the anterior lobe of the pituitary gland, but the precise co-localization of SgIII with pituitary hormones remains unknown.
Five types of endocrine cells are present in the anterior lobes of mammalian pituitary glands: corticotropes, gonadotropes, thyrotropes, mammotropes, and somatotropes. Each endocrine cell possesses characteristic secretory granules and secretes distinct peptide hormones in response to appropriate stimuli. Further understanding of the precise localization of SgIII in these endocrine cells of the anterior pituitary would provide us with important information on its putative function in endocrine cells. Moreover, we recently identified SgIII as a physiological binding protein for CgA by yeast two-hybrid screens, and demonstrated that the deletion of a SgIII-binding domain of CgA results in missorting of CgA into a constitutive secretory pathway in endocrine-derived AtT-20 cells (Hosaka et al. 2002). These findings suggest that SgIII plays an important role in the sorting and retention processes of regulated secretory proteins, such as other granins and peptide hormones, in endocrine cells.
The present report describes the immunocytochemical localization of SgIII and other granins in the anterior lobe of the male rat pituitary gland. For this purpose we raised an antiserum against a recombinant protein containing a partial amino acid sequence of rat SgIII. The results showed that SgIII was distributed in anterior pituitary endocrine cells, except for somatotropes, and particularly in mammotropes and thyrotropes. Interestingly, its localization in these endocrine cells was limited to the peripheral region of secretory granules together with CgB, suggesting a role in the organization of secretory granules. We also discuss the relationship between granin expression and the cell lineage of endocrine cells in the anterior lobe of the pituitary gland, by comparing the localization of SgIII with those of other granins.
Materials and Methods
Antibodies
A rabbit polyclonal anti-SgIII antiserum (SgIII-C#6) was raised against a GST-fusion protein containing the C-terminal sequence of rat SgIII (residues 373–491). The antigen emulsified with an equal volume of TiterMax (CytRx; Norcross, GA) was injected SC to rabbits six times at 2-week intervals. The rabbits were bled 7 days after the sixth injection, and specificities of the antisera were checked with immunoblotting analyses (see Results).
Rabbit polyclonal anti-CgA (CgA-C#101), anti-CgB (CgB-C#4), and anti-SgII (SgII-C#23) antisera were raised against synthetic peptides containing C-terminal sequences of rat CgA (amino acid residues 430–442; AIEAELEKVA-HQL), CgB (residues 602–618; YDSEEQMGPHQEAEDEK), and SgII (residues 568–583; YLNQEQAEQGREHLAK), respectively. These peptides were conjugated to keyhole limpet hemocyanin (Sigma Chemical; St Louis, MO), emulsified with TiterMax, and injected SC to rabbits as described above. The specificities of the antisera obtained were also checked with immunoblotting analyses (see Results).
To identify the various endocrine cell types in the anterior pituitary, the following antibodies and antisera were used: anti-ACTH [rabbit polyclonal (code AB902), Chemicon, Temecula, CA]; anti-human LHβ [mouse monoclonal (code O374), Immunotech, Marseille, France]; anti-TSHβ [rabbit polyclonal; generated by Dr. Wakabayashi (Gunma University, Japan)]; anti-prolactin [mouse monoclonal (code 12205), QED Bioscience, San Diego, CA]; and anti-GH [rabbit polyclonal (code AB940), Chemicon].
Immunoblotting
For immunoblotting analyses, three male Wistar rats were used. The pituitaries and adrenal glands obtained from each rat were individually homogenized with 1 ml of 0.01 M Hepes/NaOH (pH 7.4) containing 0.15 M NaCl, 1% Triton X-100, 1 mM EDTA, 10 μg/ml leupeptin (Peptide Institute; Osaka, Japan), 10 μg/ml pepstatin (Peptide Institute), and 0.4 mM p-amidino-phenyl methanesulfonyl fluoride (WAKO Pure Chemicals; Osaka, Japan). Homogenized samples from each rat were individually stored on ice for 1 hr and centrifuged at 10,000 X g for 20 min at 4C to remove debris.
The extracts from each rat were loaded onto 10% SDS-PAGE under reducing conditions, and transferred to nitrocellulose membranes according to Towbin et al. (1979). The membranes were soaked in PBS containing 5% BSA (Sigma Chemical) to block nonspecific binding and then incubated with antisera against rat granins. Dilutions of antisera were 1:2000 (anti-CgA, SgII, and SgIII), or 1:1000 (anti-CgB). Immunodetection was carried out with a chemiluminescent ECL kit (Amersham Pharmacia Biotech; Poole, UK) according to the manufacturer's protocol. The electrophoresis and immunoblotting detection were independently repeated three times to confirm the results.
Tissue Preparation
Sixteen male Wistar rats were purchased at 8 weeks of age and then housed for 2 weeks in a well-ventilated room (temperature 23 ± 1C; relative humidity 55–65%) with food and water ad libitum. The lighting condition was defined as a 12:12-hr dark-light cycle (lights on 0700–1900 hr).
For light microscopy, eight rats were sacrificed by cervical dislocation under light ether anesthesia. The pituitaries were quickly removed, cut into small pieces, and processed according to Grube and Kusumoto (1986). Briefly, pituitary tissue samples were quenched in liquid nitrogen at −210C, freeze-dried at −35C for 72 hr, fixed by vapor-phase diethylpyrocarbonate (60C, 3 hr), and embedded in epoxy resin (Araldite). Serial sections of 0.5 μm were cut and mounted on microscopic slides.
For immunoelectron microscopy, eight rats were anesthetized with pentobarbital (25 mg/kg, IP) and perfused with physiological saline, followed by a solution of 0.1% glutaraldehyde-4% p-formaldehyde in 0.1 M phosphate buffer (pH 7.2). After fixation by perfusion, pituitaries were quickly excised, cut into small pieces, and immersed in the same fixative at 4C for 2 hr. After washing thoroughly with 0.1 M phosphate buffer, pH 7.2, containing 7.5% sucrose, samples were dehydrated with 70% ethanol (1 hr, 4C) and infiltrated into pure LR White resin (London Resin; Basingstoke, Hampshire, UK) at 4C for 12 hr. The pituitaries were then placed at the bottom of gelatin capsules filled with fresh LR White resin and polymerized at 55C for 24 hr.
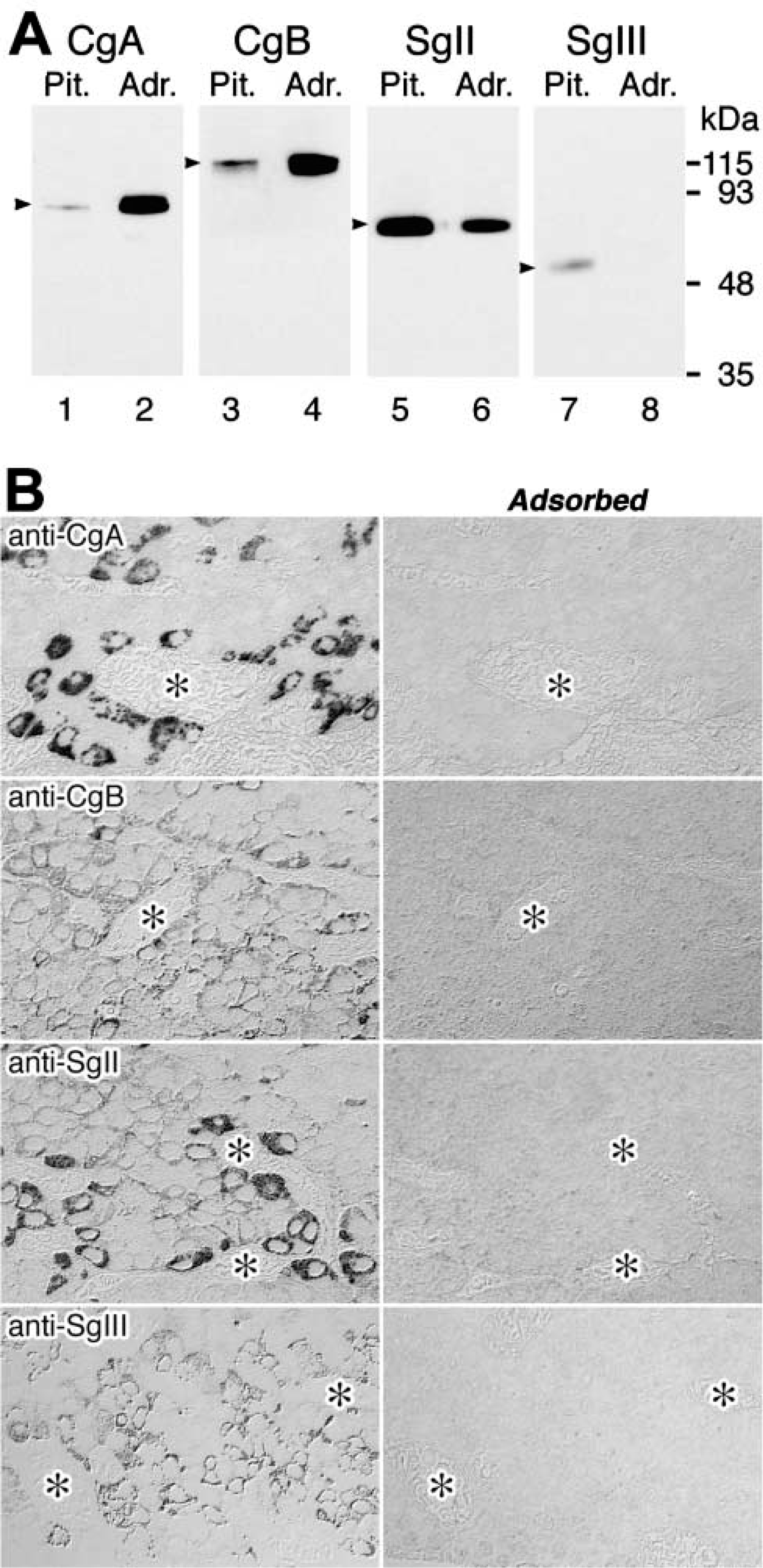
Characterization of antisera against granins by immunoblotting analyses (
Immunocytochemistry
For light microscopic immunocytochemistry, after removal of the resin by sodium methoxide (Grube and Kusumoto 1986) the semithin sections were treated with 0.3% H2O2 (20C, 30 min), incubated with 2% normal goat serum (20C, 30 min), and then incubated with antibodies and antisera against hormones and granins at 4C for 12 hr. They were next incubated with appropriate biotinylated secondary antibodies (goat anti-rabbit or anti-mouse IgG; Vector Laboratories, Burlingame, CA) at 20C for 1 hr and then with avidin-biotin complex (Vector Laboratories) at 20C for 1 hr. Staining for peroxidase was performed using 0.7 mM 3′3′-diaminobenzidine tetrahydrochloride (DAB) and 0.002% H2O2 in 0.05 M Tris-HCl buffer, pH 7.6, for 10 min. Between the various steps of the protocol, the sections were thoroughly rinsed in 0.01 M phosphate-buffered 0.5 M saline, pH 7.6, containing 0.1% Tween-20.
For immunoelectron microscopy, immunogold labeling was performed as described previously (Watanabe et al. 1993). Briefly, ultrathin sections from tissue blocks embedded in LR White resin were incubated with 5% normal goat serum for blocking at 20C for 20 min, and then further incubated with the following rabbit antisera at 4C for 12 hr: anti-rCgA (diluted 1:1000), anti-rCgB (diluted 1:2000), anti-rSgII (diluted 1:1000), or anti-rSgIII (diluted 1:200). For double immunostaining of two different granins and hormones, the two-face technique of Bendayan (1982) was applied. Intracellular localizations of different granins and hormones were distinguished by labeling with different sizes of colloidal gold particles conjugated to goat anti-rabbit IgG and anti-mouse IgG (purchased from British Biocell International, Cardiff, UK; size of particles 5, 10, or 15 nm in diameter). For triple immunostaining of some ultrathin sections in the present study, one face of the sections was simultaneously immunostained with a mixture of a mouse monoclonal antibody MAb against prolactin (PRL; diluted 1:2000) and a rabbit polyclonal antiserum against granins, and then the other face of the sections was immunostained for another granin or hormone. Between each step, the grids were washed three times in 0.02 M Tris-HCl-buffered 0.5 M saline, pH 8.2, containing 0.1% BSA. After the immunoreactions, the sections were contrasted with saturated aqueous solutions of uranyl acetate and lead citrate and examined with an electron microscope (JEM-1010; JEOL, Tokyo, Japan).
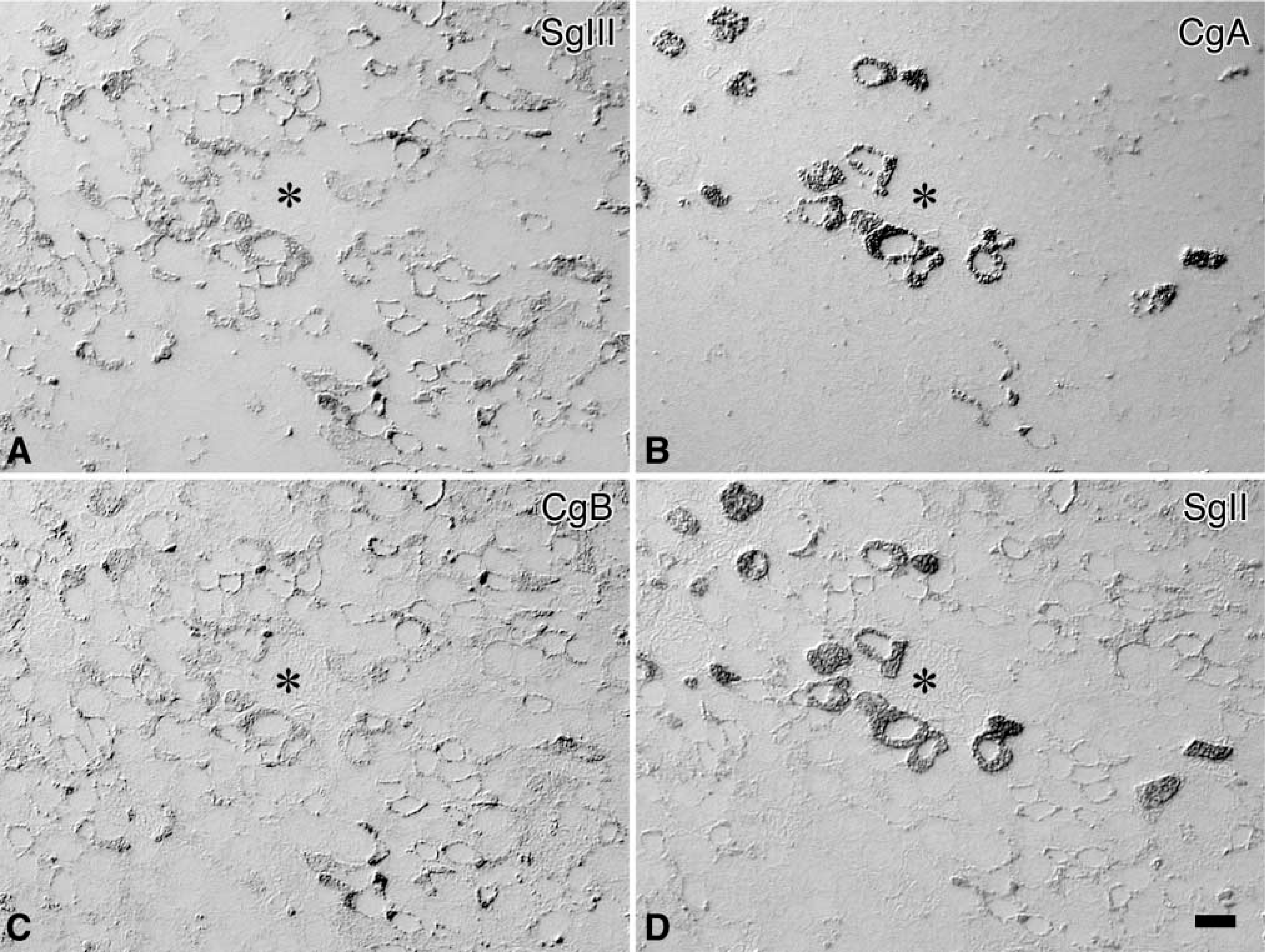
Distribution of granins in anterior lobe of male rat pituitary gland. Four serial semithin sections of the male rat anterior pituitary are immunostained for SgIII (
To semiquantitatively evaluate the subgranular localization of each granin in mammotropes and thyrotropes, cytoplasmic areas of these cells were randomly photographed at a magnification of X12,000 (10 micrographs per each type of endocrine cells) after double immunostaining for granins (SgIII, CgB, or SgII) and hormones (PRL or TSH). The profile area of each secretory granule was divided into two regions: peripheral and central. The peripheral region was defined as the outer 30% of the granule's diameter, and the central region was defined as the rest of the granule matrix in the center. The number of immunogold particles on each region of secretory granules was separately counted, and the percentages of the immunogold particles within peripheral and central regions were calculated. In addition, the distances of the immunogold particles indicative of SgIII, CgB, SgII, and PRL from the limiting membrane of the secretory granule were measured on the profile of mature mammotropes with NIH Image software (written by W. Rasband; National Institutes of Health, Bethesda, MD), and the results were statistically analyzed with Kaleida-Graph software.
Results
Characterization of Antisera Against Rat SgIII and Other Granins
Because the anterior and intermediate lobes of pituitary glands and the medulla of adrenal glands are known major storage sites for granins, the specificities of antisera used in the present study were examined by immunoblotting analyses of extracts of rat pituitary and adrenal glands. As shown in Figure 1A, the antiserum against a GST-SgIII fusion protein, including amino acid residues 373–491 of rat SgIII, properly recognized the native form of SgIII (ca. 60 kD) in the extracts of pituitary glands. The immunoreactivity for SgIII was not detectable in the extract of adrenal glands, as reported previously (Ottiger et al. 1990). Other antisera raised against the synthetic peptides covering the C-terminal regions of rat CgA (430–442), CgB (602–618), and SgII (568–583) also recognized specifically the native granin proteins of appropriate molecular masses in the extracts. The specificity of these antisera was also confirmed immunocytochemically by the fact that the reactions were specifically inhibited by addition of the corresponding antigens (Figure 1B).
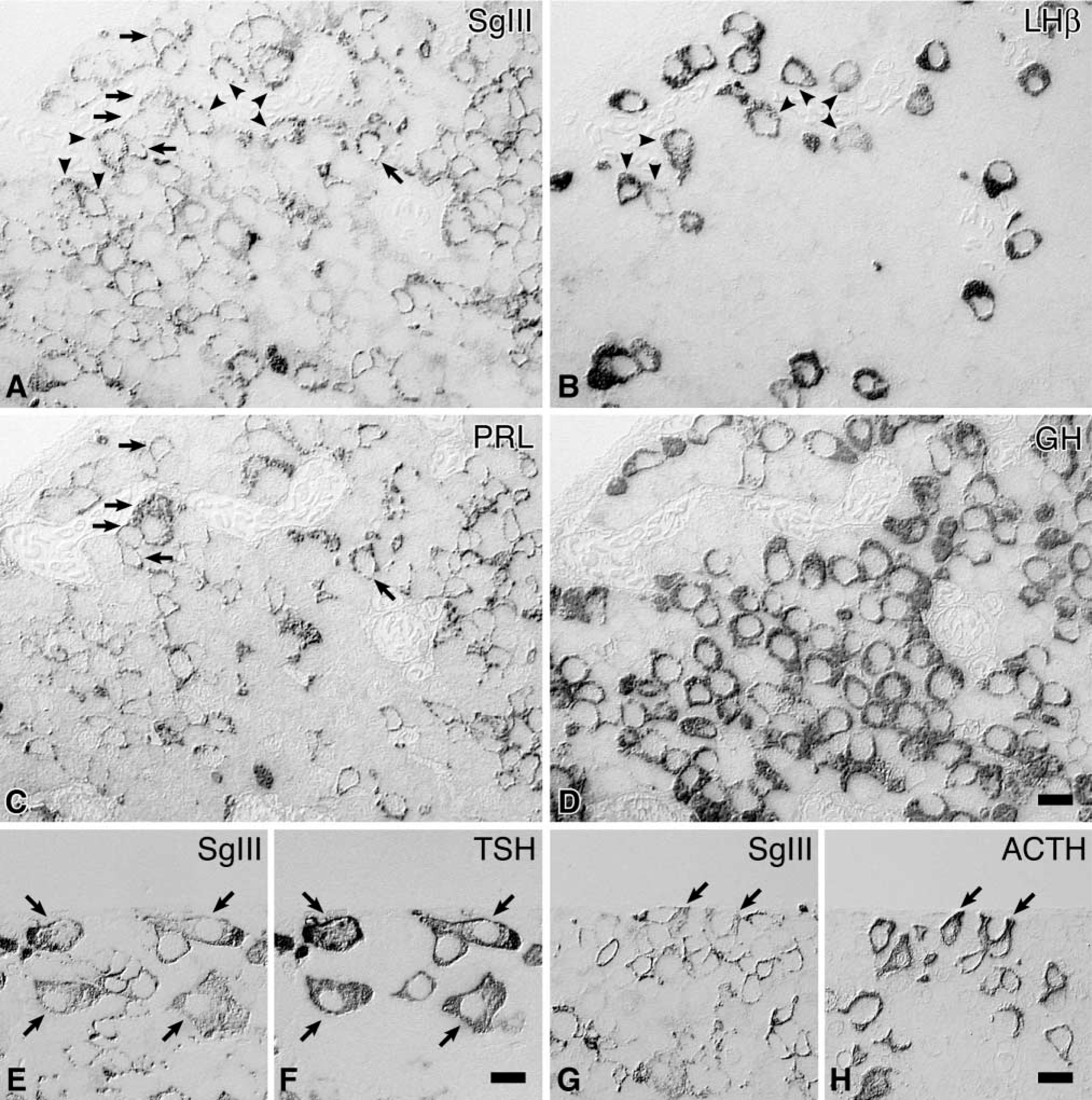
Identification of SgIII-positive cells in the anterior pituitary by immunostaining of the adjacent sections with antisera against various anterior pituitary hormones. The first set of four serial semithin sections immunostained for SgIII (
Distribution of granins in the anterior lobe of the male rat pituitary gland a

aRelative intensities of immunoreactivities are ranked on a scale of - (not detectable) to +++ (strongly positive). The subcellular localization of each granin in gonadotropes is indicated below the symbols, because male rat gonadotropes contain two distinct subtypes of secretory granules, large (large SGs) and small (small SGs), as we previously reported (Watanabe et al. 1991,1993).
Immunocytochemical Localization of SgIII in the Anterior Lobe of Male Rat Pituitary Glands
As shown in Figure 2, the majority of endocrine cells in the anterior pituitary were immunoreactive for SgIII. To compare the localization pattern of SgIII in the anterior pituitary, immunostaining of other granins was performed. Among these staining patterns, the localization pattern of SgIII in these cells resembled that of CgB (Figures 2A and 2C).
By immunostaining of adjacent sections with antisera against various pituitary hormones, four types of pituitary endocrine cells, corticotropes, gonadotropes, mammotropes, and thyrotropes, were identified as containing SgIII (Figure 3). The intensities of immunoreactivity for SgIII were relatively high in mammotropes and thyrotropes, moderate in corticotropes and gonadotropes. No significant immunoreactivity for SgIII was observed in somatotropes. The immunocytochemical localizations of four granins in the anterior pituitary are summarized in Table 1.
Intracellular Localization of SgIII in Pituitary Endocrine Cells
To precisely examine the intracellular localization of SgIII in pituitary endocrine cells, immunoelectron microscopy was applied to the tissues. The data showed that SgIII was localized in the secretory granules of corticotropes, gonadotropes, mammotropes, and thyrotropes, and that its intragranular localizations varied depending on cell type. The immunoreactivity for SgIII was not detected in the rough ER, Golgi complex, and on the plasma membrane of pituitary endocrine cells.
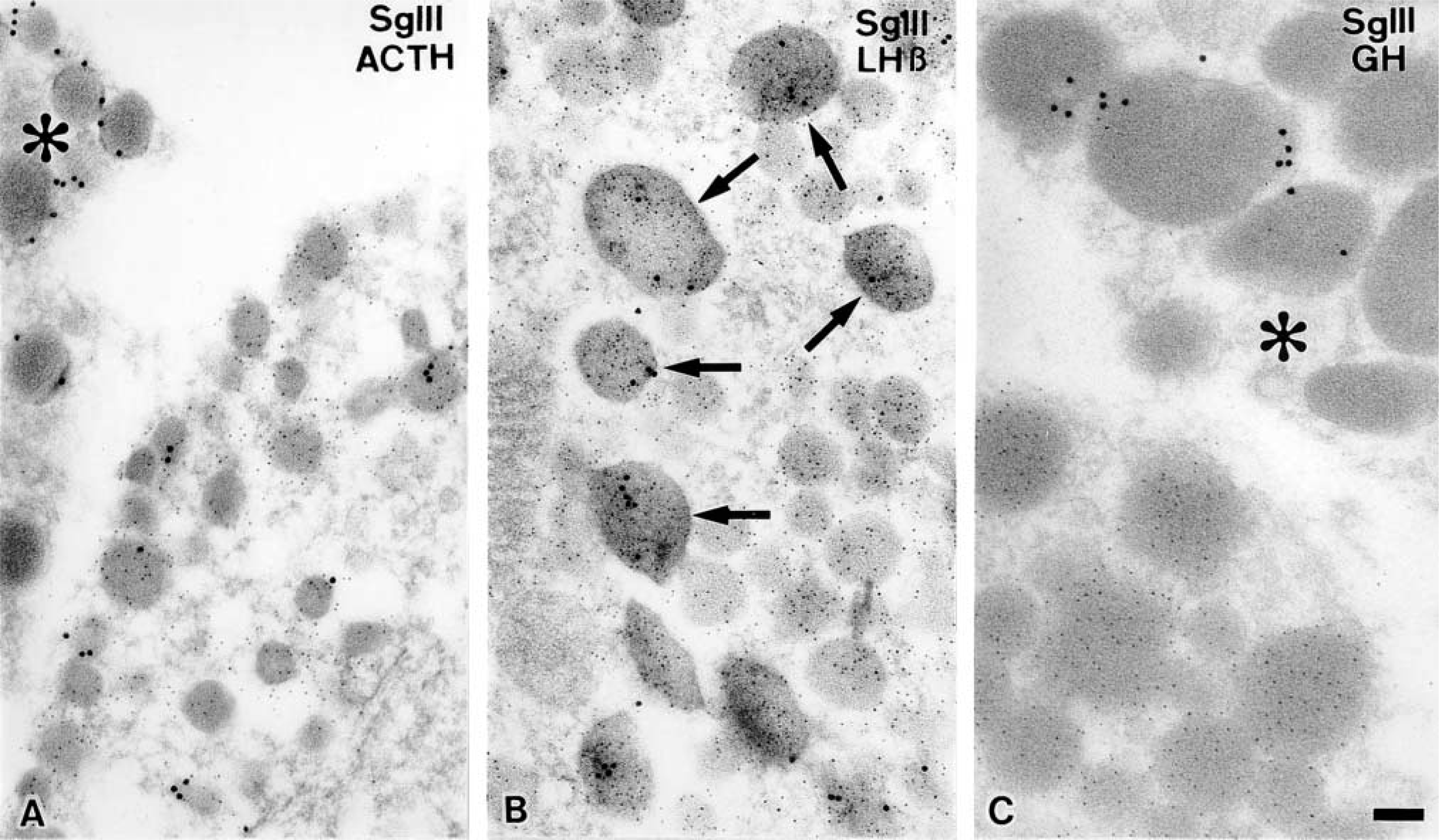
Immunocytochemical localizations of SgIII in a typical male corticotrope (
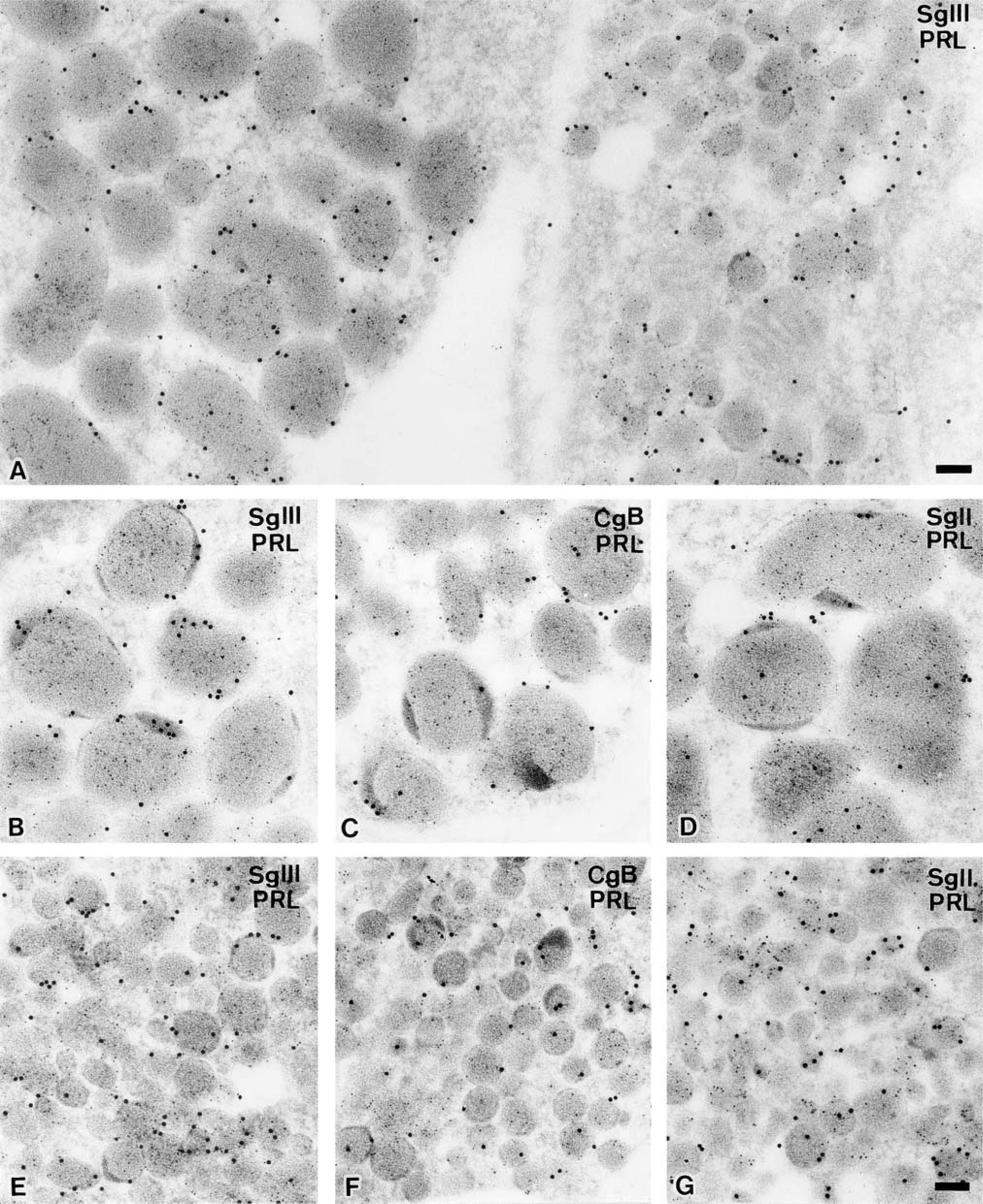
Immunocytochemical localizations of SgIII and other granins in male rat mammotropes. Secretory granules in the immature (
Proportion of immunogold particles localized in peripheral regions within secretory granules in mammotropes and thyrotropes a
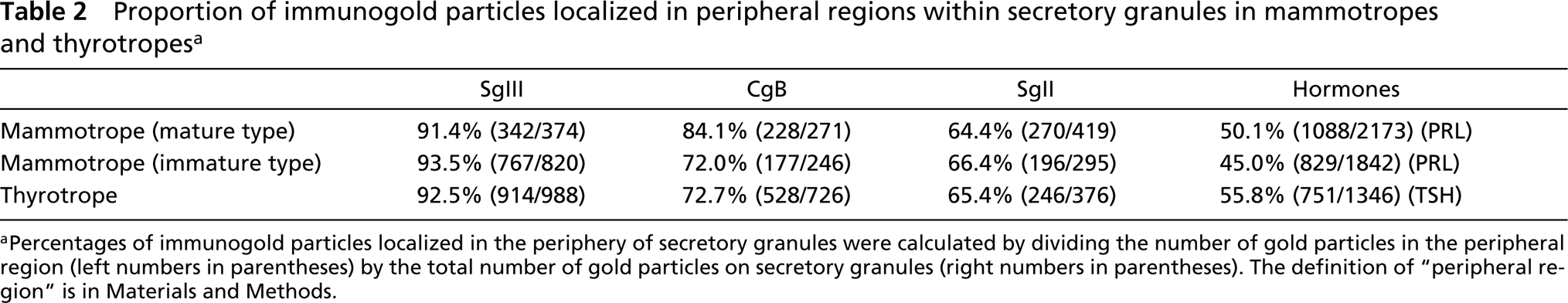
aPercentages of immunogold particles localized in the periphery of secretory granules were calculated by dividing the number of gold particles in the peripheral region (left numbers in parentheses) by the total number of gold particles on secretory granules (right numbers in parentheses). The definition of “peripheral region” is in Materials and Methods.
Corticotropes, Gonadotropes, and Somatotropes
In corticotropes, SgIII was co-localized with ACTH (or its precursor, POMC) in some secretory granules, although labeling density was relatively low (Figure 4A).
Immunogold particles indicative of SgIII were also observed in a limited population of secretory granules within gonadotropes (Figure 4B). The secretory granules immunoreactive for SgIII appeared as large-sized ones in which CgA was specifically accumulated, as we previously reported (Watanabe et al. 1991,1993). The precise co-localization of SgIII and other granins in the pituitary gonadotropes is described in our previous study (Hosaka et al. 2002).
In contrast to these endocrine cells, secretory granules of somatotropes, which contain predominantly GH, were not significantly labeled with the immunogold particles indicative of SgIII, indicating that in a physiological state somatotropes in the rat pituitary gland do not store a detectable level of SgIII (Figure 4C).
Mammotropes
It is widely accepted that there are at least two subtypes of mammotropes, an immature type containing small round secretory granules and a mature type containing large, irregular-shaped granules. Secretory granules of both subtypes of mammotropes, which contain predominantly PRL, were intensely labeled with the immunogold particles indicative of SgIII (Figure 5A). Interestingly, these immunogold particles were specifically concentrated in the peripheral region of the secretory granules, whereas those indicative of PRL were distributed all over the secretory granules. This peripheral localization in the granules was more prominent in mature mammotropes (Figure 5B), but a similar tendency was discerned in the small secretory granules in immature mammotropes (Figure 5E). Immunoreactivities for another two granins, CgB and SgII, were also observed preferentially in the periphery of secretory granules, although the accumulation of these granins in this region was less apparent than that of SgIII (Figures 5C, 5D, 5F, and 5G). These findings were confirmed by counting separately the number of immunogold particles indicative of each granin in the peripheral and central regions of secretory granules (Table 2) and by measuring the distances between each immunogold particle and limiting membrane of secretory granules in mature mammotropes (Figure 6). Secretory granules of mammotropes were not significantly labeled with the immunogold particles indicative of CgA (data not shown).
Thyrotropes
In addition to mammotropes, immunolabeling of SgIII was intensely detected in endocrine cells containing small-sized secretory granules that were labeled densely with anti-TSHβ antiserum (Figure 7A). The size and appearance of the secretory granules in thyrotropes could hardly be distinguished from those of the immature type in mammotropes. Immunogold particles showing SgIII were localized predominantly in the periphery of secretory granules in thyrotropes, as demonstrated in mammotropes (Table 2). The expression patterns of other granins in thyrotropes were also similar to that in mammotropes; CgB and SgII were expressed in these cells but CgA was not (Figures 7B and 7C).
Although secretory granules in the greater parts of thyrotropes and mammotropes contain exclusively just one of the hormones, TSH or PRL (Figures 7D and 7E), an intermediate type of endocrine cells containing both TSH and PRL was found among the endocrine cells immunoreactive for SgIII (Figure 7F). This type of endocrine cell, however, was rarely but distinctly found in the anterior pituitary of the adult male rat. This finding is compatible with the fact that mammotropes and thyrotropes originate from a common progenitor of pituitary cells, which is determined by a transcriptional factor Pit-1 during the developmental stages of the gland.
Discussion
After its identification, SgIII's distribution and function were not investigated as extensively as were other granins, probably because mice missing the 1B1075 gene that encodes SgIII exhibit no obvious defects in viability, fertility, or locomotor behavior (Kingsley et al. 1990). However, a recent study on the expression of SgIII has demonstrated that the mRNA levels of SgIII in the intermediate lobe of the Xenopus pituitary fluctuate drastically during background adaptation (Holthuis and Martens 1996). More recently, we identified SgIII as a physiological binding protein for CgA and demonstrated that a SgIII-binding domain of CgA is indispensable for the proper sorting and retention of CgA in secretory granules within AtT-20 cells (Hosaka et al. 2002). These findings suggest that SgIII plays a particular role in endocrine functions.
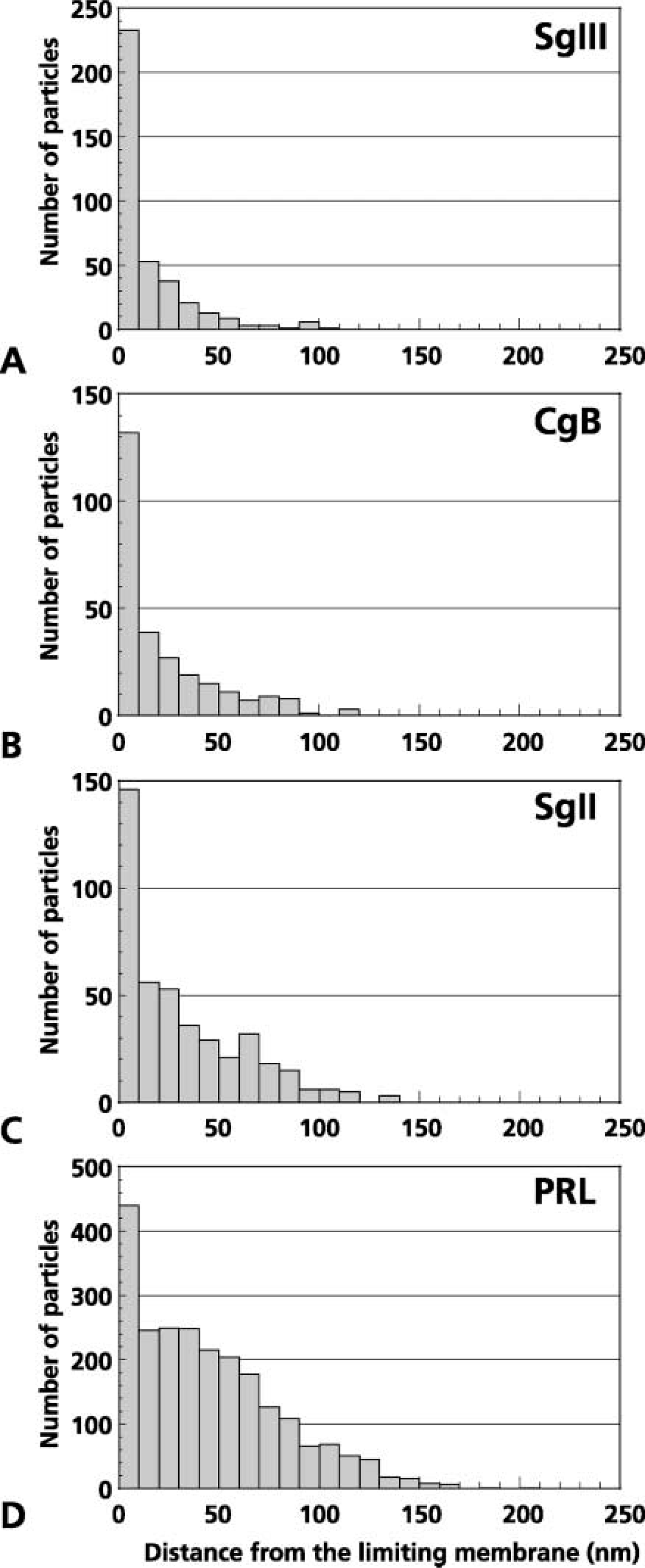
Distribution of distances of immunogold particles indicative of SgIII (
The present study demonstrated that SgIII was intensely expressed in mammotropes and thyrotropes in the anterior lobe of the pituitary gland, and was preferentially co-localized with CgB in the peripheral region within secretory granules. Immunoreactivity for CgA is not detectable in the mammotropes and thyrotropes, suggesting that SgIII in these cells contributes to the sorting and retention of other granins and/or peptide hormones instead of CgA.
The peripheral localization of certain granins within secretory granules has been reported in other endocrine cells. CgA in the pancreatic A-cell is localized more closely to the secretory granule membrane within the secretory granule matrix than is the costored hormone glucagon (Ehrhart et al. 1986). In mammotropes and GH3B6 cells derived from a mammosomatotropic tumor, CgB is localized preferentially at the peripheral region of secretory granules, whereas PRL concentrates in their core region (Ozawa et al. 1994).
Putative specific interaction between granins and the secretory granule membrane have also been demonstrated by biochemical analyses. CgB is tightly associated with the secretory granule membrane of PC12 cells (Pimplikar and Huttner 1992), and a conserved loop structure with a disulfide bond located near the N-terminus of CgA and CgB is associated with membrane fractions of secretory granules prepared from adrenal chromaffin cells (Yoo 1996,1993a,1993b). These findings suggest that certain granin proteins interact with membrane-bound proteins or lipid microdomains on the secretory granule membrane.
It is believed that, to be transported properly to secretory granules, soluble secretory proteins are first aggregated, and the aggregates then may be anchored to certain molecules on the membrane destined for secretory granules (Thiele et al. 1997; Arvan and Castle 1998; Tooze 1998). Semiquantitative analyses on the intragranular distribution of each granin in the present study indicated that SgIII is localized more prominently in the peripheral region within secretory granules of mammotropes and thyrotropes than CgB. These lines of evidence suggest that SgIII is one of the membrane-associated granin proteins, which have been indicated as contributing to gathering and anchoring other soluble constituents of secretory granules to the membranes.
It is also important to understand whether SgIII and CgB function cooperatively or redundantly in endocrine cells of the anterior pituitary. Considering that no clear-cut phenotype appears in mice deficient solely in SgIII (Kingsley et al. 1990), these two granin proteins may function redundantly during storage and transport of hormones in the trans-Golgi network and secretory granules of the endocrine cells. Further studies are required to evaluate the physiological roles of SgIII and/or CgB in pituitary endocrine cells.
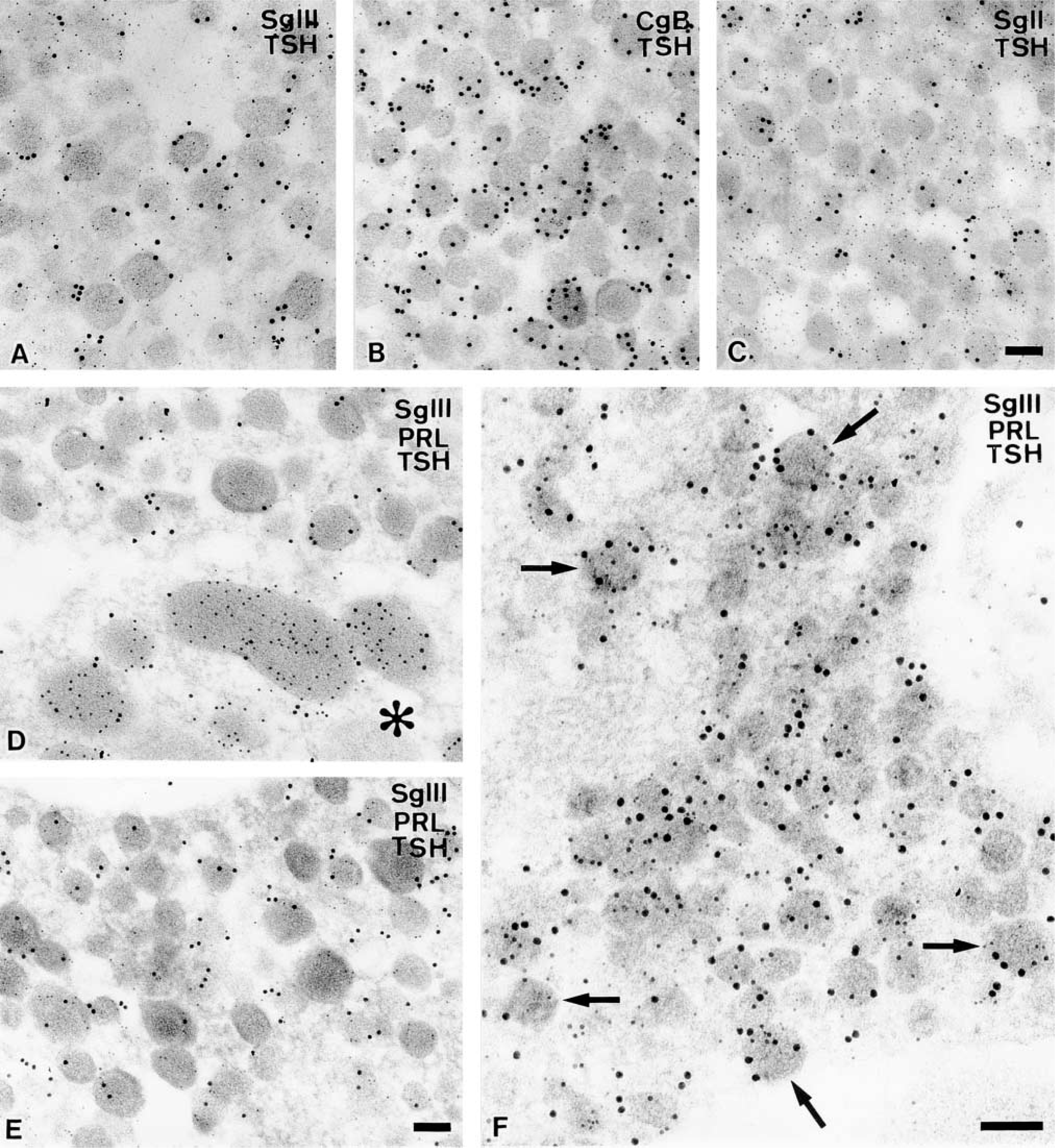
Immunocytochemical localizations of SgIII and other granins in male rat thyrotropes. Small secretory granules labeled with anti-TSHβ (indicated by 5-nm gold particles) are concomitantly labeled with colloidal gold particles indicative of SgIII (15 nm) (
The similarity in the expression and subcellular localization of these two granins in mammotropes and thyrotropes reminds us that these two types of endocrine cells originated from a common precursor cell, determined by a pituitary-specific transcriptional factor, Pit-1 (Li et al. 1990; for reviews see Asa and Ezzat 1999; Treier and Rosenfeld 1996; Scully and Rosenfeld 2002). We observed SgIII-positive intermediate endocrine cells that expressed both PRL and TSH. Similar intermediate types of endocrine cells containing both PRL and TSH have been reported in rat (Losinski et al. 1989) and cow (Bassetti et al. 1990) anterior pituitaries, suggesting that common precursor-like cells of these two endocrine cells reside even in the pituitary glands of adult animals.
To date, no study has shown a direct relationship between pituitary-specific transcriptional factors, such as Pit-1, and the regulation of granin expression. Recent analyses on promoter regions of CgA, CgB, and SgII genes have clearly demonstrated that some characteristic elements such as CRE (cAMP response element) and G/C-rich domains contribute to the neuroendocrine-specific and secretagogue-inducible expression of granins in endocrine cells (Jungling et al. 1994; Mahata et al. 1999; Mahapatra et al. 2000). However, details regarding the regulatory mechanisms of SgIII expression have not been analyzed, and the differences in expression patterns of each granin among the various endocrine cells have yet to be explained. Further investigation is required to understand the issue of whether pituitary-specific transcriptional factors could regulate the expression of granins including SgIII, which might contribute to the establishment of characteristic ultrastructures and the size of secretory granules in each type of endocrine cell.
In summary, the present immunocytochemical study demonstrated that SgIII in the rat anterior pituitary gland is distributed in all types of endocrine cells except for somatotropes. Among the granin proteins examined, the localization pattern of SgIII was similar to that of CgB at both light and electron microscopic levels. In particular, peripheral localizations of SgIII and CgB within secretory granules of mammotropes and thyrotropes suggested that these two granins might be membrane-associated proteins and play a role in anchoring other soluble constituents of secretory granules.
Footnotes
Acknowledgements
We thank Dr K. Wakabayashi (Gumma University, Japan) for the anti-TSHβ antiserum.
Supported in part by a grant-in-aid for Scientific Research (C) from the Japan Society for the Promotion of Science (JSPS) and by a grant from the Akiyama Foundation, Japan.