
Abstract
p63 is essential for epithelial cell survival and may function as an oncogene. We examined by immunohistochemistry p63 expression in human normal and tumor salivary gland tissues. In normal salivary glands, p63 was expressed in the nuclei of myoepithelial and basal duct cells. Among 68 representative salivary gland tumors, 63 displayed p63 reactivity. In all tumor types differentiated towards luminal and myoepithelial lineages (pleomorphic adenomas, basal cell adenomas, adenoid cystic carcinomas, and epithelial-myoepithelial carcinomas), p63 was expressed in myoepithelial cells, whereas luminal cells were always negative. Similarly, in mucoepidermoid carcinomas, basal, intermediate, and squamous cells expressed p63, in contrast to luminal mucous cells. p63 reactivity was also restricted to basal cells in Warthin tumors and oncocytomas. Myoepitheliomas and myoepithelial carcinomas all expressed p63. The only five negative tumors were three of four acinar cell carcinomas and two of three adenocarcinomas. In conclusion, p63 is expressed in the nuclei of normal human salivary gland myoepithelial and basal duct cells. p63 expression is retained in the modified myoepithelial and basal cells of human salivary gland tumors, which suggests a role for p63 in oncogenesis of these complex tumors.
The p63 gene, mapping to 3q27–29, is the third member of the p53 family (Schmale and Bamberger 1997; Osada et al. 1998; Senoo et al. 1998; Trink et al. 1998; Yang et al. 1998; Thanos and Bowie 1999; Tan et al. 2001; Zeng et al. 2001). Unlike p53, p63 transcription from two different promoters and alternative splicing in the C-terminal region are responsible for at least six isoforms. Three isoforms (TA-α, TA-β, and TA-γ) contain the N-terminal transactivation (TA) domain, activate transcription from p53-responsive promoters, and can induce apoptosis (Osada et al. 1998; Yang et al. 1998). The ΔNp63 isoforms lack the TA domain and may function as a dominant-negative factor that inhibits the transactivation functions of p53 and TAp63 proteins (Yang et al. 1998).
The p63 −/− mice are characterized by agenesis of stratified squamous epithelia and their derivatives, including mammary, sebaceous, and salivary glands, and defects in limb and craniofacial structures (Mills et al. 1999; Yang et al. 1999). Germline p63 mutations in human patients are responsible for rare limb malformation syndromes (Celli et al. 1999; Ianakiev et al. 2000). These phenotypes have led Yang et al. (1999) to propose that p63 is essential for the survival of epithelial stem cells.
In mouse embryos, p63 is expressed primarily within the ectoderm (Mills et al. 1999). In normal adult human and murine tissues, p63 is expressed in the epithelial basal cells of the skin, the uterine cervix, and the urogenital tract (Yang et al. 1998). Signoretti et al. (2000) reported that p63 is expressed in the prostate basal cells and that p63 is required for prostate development. In addition, p63 expression may be a marker for certain epithelial progenitor cells in the esophagus, including mucosal gland ducts (Glickman et al. 2001). In a recent work, p63 protein has been detected in many human normal tissues: esophagus, tonsil, breast, exocervix, prostate, larynx, lung, skin, bladder, and kidney. In this work, normal salivary glands were not tested (Di Como et al. 2002). Furthermore, p63 is a selective nuclear marker of the myoepithelial cells in the human breast, as shown by Barbareschi et al. (2001).
Somatic p63 mutations are exceedingly rare in human tumors (Osada et al. 1998). p63 is not likely to be a classical tumor suppressor gene (Yang et al. 2002). Several authors have demonstrated high levels of p63 protein in squamous-cell carcinomas of the aerodigestive tract, which frequently show amplification of the 3q27–29 region (Bockmuhl et al. 1996; Hibi et al. 2000; Faridoni-Laurens et al. 2001; Choi et al. 2002). p63 expression has also been observed by immunohistochemistry in lung, skin, and cervical squamous-cell carcinomas (Parsa et al. 1999; Hibi et al. 2000; Quade et al. 2001). In addition, by screening a large number and variety of neoplasms, Di Como et al. (2002) have detected p63 protein in urothelial carcinomas, thymomas, rhabomyosarcomas, and non-Hodgkin lymphomas. In contrast, carcinomas derived from glandular epithelia, including adenocarcinomas of the breast or prostate, fail to consistently express p63 (Signoretti et al. 2000; Barbareschi et al. 2001; Di Como et al. 2002). Salivary gland tumors appear to be an exception to this rule. Di Como et al. (2002) reported p63 expression in four pleomorphic adenomas and two of four carcinomas of the salivary glands, without further mentioning tumor type or marked-cell type. Pelosi et al. (2001) have detected p63 protein in one case of bronchial epithelial-myoepithelial carcinoma. Moreover, Barbareschi et al. (2001) have detected p63 immunoreactivity in two breast adenoid cystic carcinomas.
Salivary glands comprise two lineages: one epithelial luminal lineage and one myoepithelial lineage. Myoepithelial cells express α-smooth muscle actin (Gugliotta et al. 1988), smooth muscle myosin heavy chain (Longtine et al. 1985), and calponin (Carmichael et al. 1994), as well as cytokeratin 14 (Dardick et al. 1987). They represent a basal compartment, which may host stem cells. In the p63 −/− mice, agenesis occurs in tissues in which progenitor cells express p63. Because salivary glands are also absent in p63 −/− mice (Yang et al. 1999), we reasoned that the basal or myoepithelial compartment of normal human salivary glands expresses p63. We also determined whether p63 is expressed in the modified myoepithelial or basal cells, which are an important component of most salivary gland tumors (Ellis and Auclair 1996).
Materials and Methods
Tissue Samples
We investigated by immunohistochemistry six normal and 68 tumor paraffin-embedded samples from human salivary gland: 15 pleomorphic adenomas, four basal cell adenomas, four oncocytomas, four myoepitheliomas, one canalicular adenoma, four Warthin tumors, 11 adenoid cystic carcinomas, nine mucoepidermoid carcinomas, four acinar cell carcinomas, one basal adenocarcinoma, two epithelial-myoepithelial carcinomas, three myoepithelial carcinomas, three adenocarcinomas, NOS, two polymorphous low-grade adenocarcinomas, and one carcinoma ex pleomorphic adenoma. All tumor samples contained salivary gland parenchyma or squamous epithelium adjacent to the tumor. Paraffin-embedded normal human breast tissue from reduction mammoplasty was used as a positive control for p63 immunostaining. Frozen tissue samples from normal human skin and submandibular gland were also immunostained.
Study cases were retrieved from the files of the Service d'Anatomie et de Cytologie Pathologiques, Groupe Hospitalier Pitié-Salpětrière, Paris, France. All cases were reviewed by two pathologists (PF and AHL) and were classified using the diagnostic criteria of the World Health Organization International Histological Classification of Tumors (Seifert 1991).
Primary Antibodies
Two different anti-p63 antibodies were used: the mouse anti-human monoclonal antibody (MAb) 4A4 (NeoMarkers; Freemont, CA) was raised against amino acids 1–205 mapping at the NH2 terminus of ΔNp63; the mouse anti-human MAb 7JUL (Novocastra; Newcastle upon Tyne, UK) was raised against amino acids 319–410. Both MAbs recognize all the different p63 isoforms. MAb 4A4 gave the strongest staining on paraffin sections but failed to localize p63 on frozen sections. Immunoreactivity with MAb 7JUL was similar to 4A4 staining on paraffin sections and could be detected on frozen sections. The mouse anti-human monoclonal antibody CKB1 (Sigma; St. Louis, MO), which recognizes cytokeratin 14, was used in frozen sections to stain myoepithelial cells.
Immunohistochemical Analysis
Immunohistochemical staining was performed according to standard procedures. Antigen retrieval was performed by bainmarie heating for 30 min at 98C in 10 mmol/liter citrate buffer, pH 6.0 (BioGenex; San Ramon, CA). Endogenous peroxidase activity was blocked by incubation in 3% H2O2 for 5 min. MAb 4A4 was applied at room temperature (RT) for 1 hr at a 1:200 dilution. The detection step was performed using an LSAB+ kit and DAB (DAKO; Carpinteria, CA) as chromogen. Slides were counterstained with hematoxylin. A positive control (normal human breast tissue) was included in each analysis. As negative staining control, the antibody anti-p63 was substituted for a mouse monoclonal anti-IgG of the same subclass (anti-IgG1; Sigma).
Double Immunofluorescence Staining
The 7JUL (anti-p63) and CKB1 (anti-cytokeratin 14) antibodies were used to realize double immunofluorescence staining in acetone-fixed frozen tissue sections of human normal salivary gland. A sequential method was performed. We used the Vectastain Universal ABC-AP kit (Vector Laboratories; Burlingame, CA) with a red substrate (Fast Red; DAKO) to detect p63 first. Nonspecific binding was blocked by horse serum. Slides were incubated overnight at 4C with MAb 7JUL at a 1:50 dilution. Fast Red was applied for 30 min. As a negative control for this staining, the 7JUL antibody was substituted for a mouse MAb of the same subclass (anti-IgG1). After rinsing, slides were incubated with goat serum and directly incubated with the CKB1 antibody at a 1:50 dilution for 60 min at RT. After washing, cytokeratin 14 signals were detected by a second FITC-conjugated antibody (FITC-conjugated AffiniPure F(ab′)2 fragment goat anti-mouse IgM μ-chain-specific; Jackson ImmunoResearch Laboratories, West Grove, PA) at a 1:100 dilution for 30 min. Slides were then rinsed and coverslipped with Immu-Mount (Chandon; CML, Nemours, France). As a negative staining control MAb CKB1 was substituted for a mouse monoclonal anti-IgM (Sigma). The preparations were analyzed with a Zeiss Axioplan 2 Imaging microscope. p63 detected with Fast Red substrate was visualized using a conventional rhodamine filter set, resulting in a red emission. To observe double staining, an FITC filter set was used, resulting in an orange emission for p63 and a green emission for cytokeratin 14. Digital images were acquired using Axiovision 3.0 software.
Results
p63 Expression in Normal Salivary Gland
p63-reactive nuclei were seen in basal cells around luminal cells in intercalated, striated, and some interlobular ducts (Figure 1). Small, often elongated nuclei at the periphery of the acini were also p63-reactive. The location of these p63-reactive nuclei suggested that the p63-reactive cells in the acini were myoepithelial cells. Luminal duct cells and acinar cells were unreactive.
To confirm that p63-reactive nuclei belonged to myoepithelial cells, we realized a double immunofluorescence staining for p63 and cytokeratin 14. The p63-reactive nuclei were seen closely associated with cytokeratin 14-expressing cell processes. Although this association was seen in most cells, a few isolated p63-reactive cells were negative for cytokeratin 14.
p63 Expression in Benign Epithelial Tumors
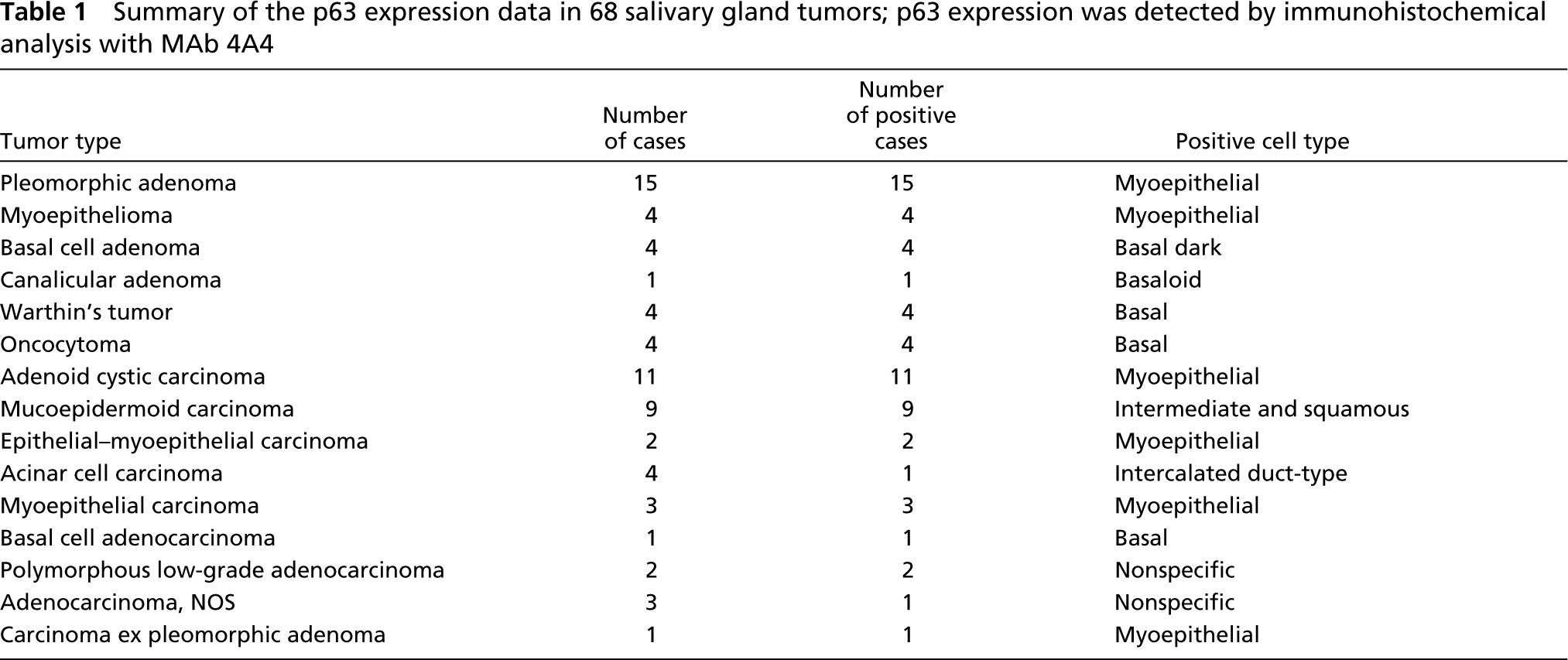
Fifteen of 15 pleomorphic adenomas demonstrated p63 nuclear staining in the modified myoepithelial cells (Figures 2a–2i). p63 reactivity was present in chondromyxoid areas as well as cellular areas, and did not correlate with tumor type (whether or not the tumor was predominantly myxoid or cellular). Tumor data are summarized in Table 1. Immunoreactivity was diffuse in 13 cases, although there was great variability in the approximate proportions of reactive tumor cells (between 10% and 90% of tumor cells). Focal positivity was observed in two cases. One had a large negative myxoid area; the other comprised a uniformly reactive myxoid zone and a large cellular zone in which only very few reactive cells were seen. All cell types were reactive, including stellate, plasmacytoid, spindle, clear cells, and cells appearing to be in transition from one type to another. The modified myoepithelial cells surrounding duct-like structures were often the most obviously reactive. p63 reactivity was also strong and consistent in basaloid cells at the periphery of squamous islands. In contrast, p63 reactivity was always absent in cuboidal or columnar epithelial cells lining duct lumens.
The neoplastic cells of the four myoepitheliomas demonstrated p63 immunoreactivity. p63 protein was more consistently seen in spindle cells, whereas plasmacytoid and clear cells, which were predominant in one case, were occasionally reactive.
p63 reactivity was prominent in palisading nuclei of the dark cells along the peripheral stromal interface of the epithelial nests in four of four basal cell adenomas. Staining of the larger light cells was more variable. All cuboidal duct cells surrounding small lumens were negative.
Immunoreactivity was found in several foci of basaloid-appearing cells in one case of canalicular adenoma of the upper lip. A few columnar cells were also p63-reactive.
Four of four Warthin tumors demonstrated strong and constant p63 reactivity in basal cells. The palisading nuclei of luminal columnar cells were unreactive, although a strong nonspecific cytoplasmic staining could be seen. Scattered stromal lymphoid cells exhibited nuclear reactivity. Oncocytes in four oncocytomas also displayed moderate to strong nonspecific cytoplasmic staining, but specific immunoreactivity could not be observed in their centrally placed nuclei. However, variable numbers of smaller reactive nuclei were seen at the stromal interface of the oncocytic cell clusters.
p63 Expression in Primary Malignant Epithelial Tumors
The 11 adenoid cystic carcinomas showed a mixture of cribriform, tubular, and solid growth patterns (Figures 2j–2t). Whatever the predominant growth pattern, the cell immunoreactivity was very consistent from one tumor to another. Abluminal cells of myoepithelial differentiation were diffusely and strongly reactive for p63. In contrast, the luminal cells were unreactive.
The nine mucoepidermoid carcinomas were predominantly cystic in four cases and solid in five cases. Using the AFIP criteria (Ellis and Auclair 1996), tumors could be classified into low (four cases), intermediate (three cases), and high (two cases) grade. Strong and diffuse nuclear staining was noted in each case. There was no correlation between p63 immunoreactivity and microscopic grade. In cystic tumors, reactive nuclei were found in foci of intermediate or squamous cells and in basal cells at the periphery of mucous cells that lined cystic spaces. In solid tumors, sheets or islands of intermediate, squamous, and clear cells demonstrated p63 nuclear reactivity. p63 protein was absent from mucous and columnar cells.
The two epithelial-myoepithelial carcinomas had a similar immunohistochemical staining profile, with clear myoepithelial cells being strongly reactive, whereas the darker luminal cells were unreactive.
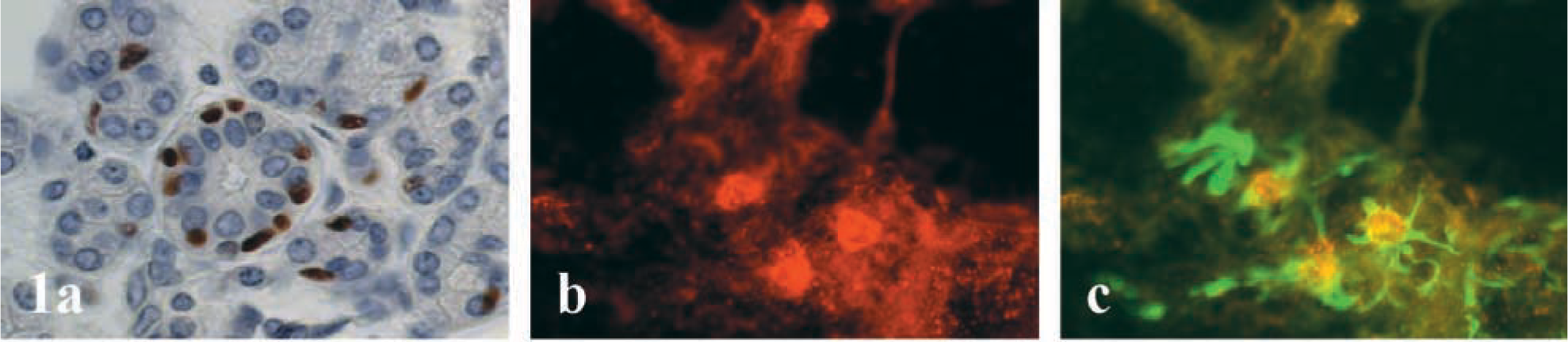
Immunohistochemical analysis of p63 expression in human normal salivary gland tissues. Representative examples of p63 nuclear immunostaining are presented. Original magnification X400. (
Summary of the p63 expression data in 68 salivary gland tumors; p63 expression was detected by immunohistochemical analysis with MAb 4A4
Three of four acinar cell carcinomas, in which various numbers of acinar, intercalated duct-like, vacuolated, and nonspecific glandular cells were observed, were entirely negative for p63. However, in one papillary-cystic case, intercalated duct-type epithelial cells were immunoreactive.
The three myoepithelial carcinomas exhibited focal reactivity in 10–30% of tumor cells.
Most basaloid tumor cells were strongly reactive in one case of basal cell adenocarcinoma.
Two polymorphous low-grade adenocarcinoma cases exhibited diffuse nuclear reactivity. Some of these p63-reactive cells lined central lumens in small tubular structures.
Of three adenocarcinoma cases, NOS, one poorly differentiated case demonstrated p63 reactivity in few scattered tumor nuclei. p63 protein was not detected in two other cases, although in one case residual p63-immunoreactive myoepithelial cells were seen around an intraductal component.
One case of carcinoma ex pleomorphic adenoma showed scattered p63-reactive tumor nuclei.
Discussion
Normal human salivary gland myoepithelium is reactive with antibodies to cytokeratins and muscle-specific actin. Among the several subtypes of cytokeratin, cytokeratin 14 is expressed most consistently and is specific for myoepithelial and ductal basal cells (Dardick et al. 1987). Here we show that normal human myoepithelial and basal ductal cells do react with antibodies to p63 protein. As opposed to S-100 protein (Dardick et al. 1991), p63 protein is detected whether or not the salivary gland parenchyma is adjacent to expanding tumors.
In contrast to p53 mutant mice, mice lacking the p63 gene have severe developmental defects involving all squamous epithelia and their derivatives, including skin, hair, and teeth, as well as mammary, lachrymal, and salivary glands (Mills et al. 1999; Yang et al. 1999). This phenotype has been attributed to a failure of the corresponding squamous or glandular tissue stem cells to be formed or maintained (Yang et al. 1999). p63 is expressed in the basal cell layer of the skin and myoepithelial cells of the breast (Yang et al. 1998; Barbareschi et al. 2001). p63 is also a prostate basal cell marker and is required for prostate development (Signoretti et al. 2000). The authors of these previous reports have generally concluded that p63-reactive cells in these tissues may include stem cells. It is also likely that the p63-immunoreactive cells in normal human salivary glands include stem cells. However, the entire population of salivary gland myoepithelial cells reacts with p63 antibodies, and this far exceeds the expected number of stem cells. Moreover, the lineage relationships between salivary gland luminal and myoepithelial cells are still matters of controversy. The epithelial compartment of the human breast, as in salivary glands, also comprises two distinct lineages: a luminal and a myoepithelial lineage. Some authors have reported that the human breast stem cell is a basal cell (Bocker et al. 2002). Others have provided evidence indicating that the luminal epithelial lineage may function as precursor cells for both luminal and myoepithelial human breast lineages (Gudjonsson et al. 2002). Our study of normal salivary gland tissue shows a strong association of p63 expression with cytokeratin 14 expression that characterizes myoepithelial and basal duct cells. However, we also found a few cells staining for p63 but not for cytokeratin 14. It remains to be determined whether this small subpopulation of cells truly represents the putative precursor cells of the human salivary glands.
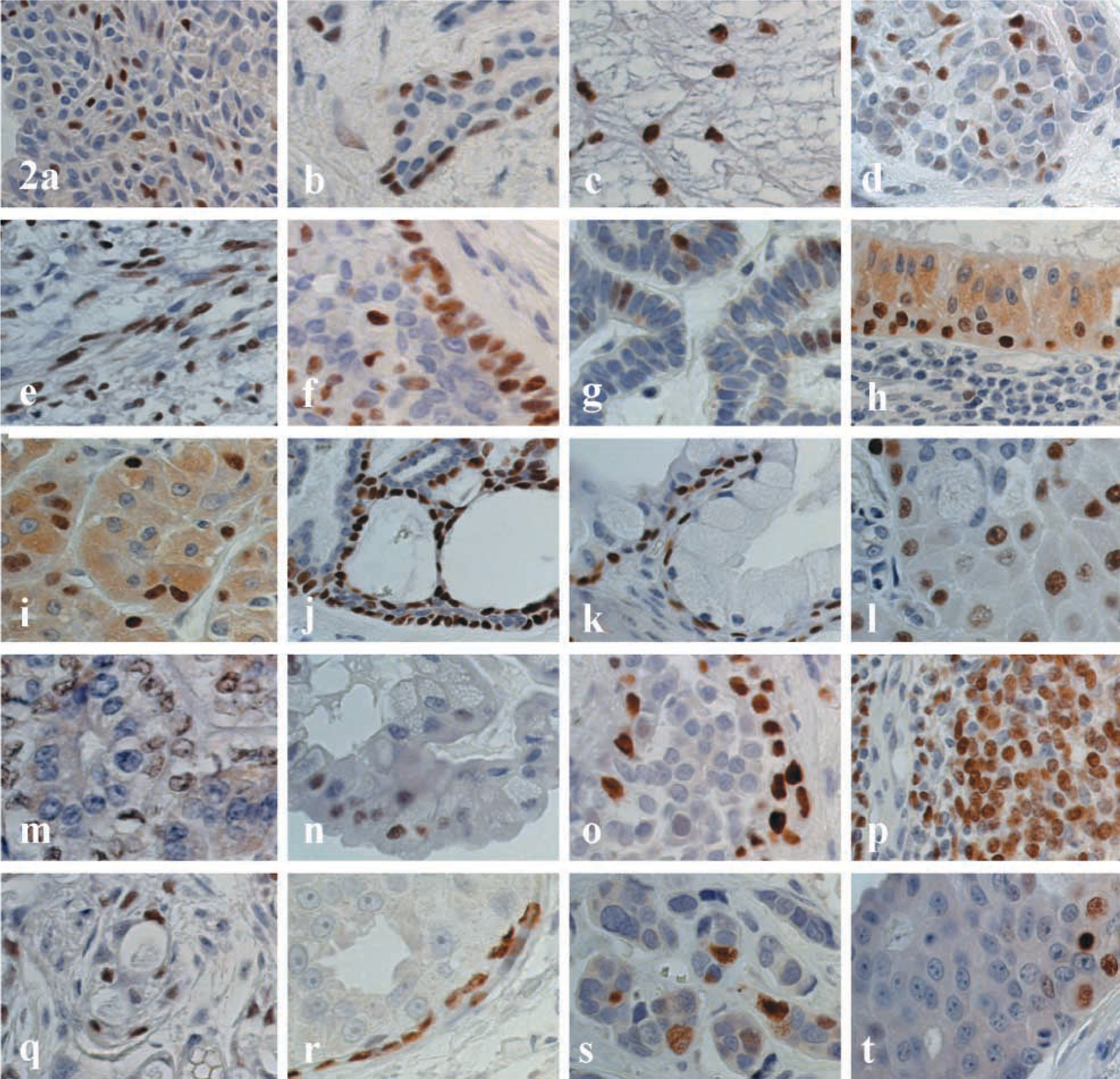
Salivary gland tumors. Immunoperoxidase staining of paraffin sections. p63 was detected with DAB (brown) and nuclei were counterstained with hematoxylin (blue). Original magnification X200. (
This report is the first to determine the frequent expression of p63 in a large series of human salivary gland tumors encompassing the most commonly encountered tumor types as well as some less common neoplasms. Among other human tumors, p63 expression is frequently found in squamous-cell carcinomas, urothelial carcinomas, thymomas, and in a subset of non-Hodgkin lymphomas (Parsa et al. 1999; Hibi et al. 2000; Faridoni-Laurens et al. 2001; Quade et al. 2001; Choi et al. 2002; Di Como et al. 2002). p63 is also expressed in the corresponding normal squamous, urothelial, thymic, and lymphoid tissues (Yang et al. 1998; Di Como et al. 2002). It is therefore not surprising that salivary gland tumors, which often include proliferating myoepithelial cells, express p63 so widely. Myoepithelial cell differentiation occurs to variable degrees in pleomorphic adenoma, myoepithelioma, basal cell adenoma, adenoid cystic carcinoma, polymorphous low-grade adenocarcinoma, epithelial-myoepithelial carcinoma, myoepithelial carcinoma, and basal cell adenocarcinoma (Ellis and Auclair 1996). Intermediate cells in mucoepidermoid carcinoma also demonstrate ultrastructural characteristics of modified myoepithelial cells (Dardick et al. 1990). In our study, every case of the above-mentioned tumors expressed p63, with consistency in all tumor types for which this could be evaluated. The salivary tumors that did not consistently express p63 were acinar cell carcinomas and adenocarcinomas, NOS. Acinar cell carcinomas are differentiated towards specialized luminal cells, whereas adenocarcinomas, NOS, lack the distinctive features of the other, more specific tumor types. In this regard, salivary gland adenocarcinoma, NOS, is similar to breast and prostate adenocarcinoma, in which a rim of non-neoplastic p63-reactive myoepithelial or basal cells surrounding unreactive carcinoma in situ cells may occur (Signoretti et al. 2000; Barbareschi et al. 2001).
One of the interesting findings in our study is the correlation between cell type and p63 expression in tumors with differentiation along myoepithelial (p63-reactive) and luminal (p63-unreactive) lineages such as pleomorphic adenoma, basal cell adenoma, adenoid cystic carcinoma, mucoepidermoid carcinoma, and epithelial-myoepithelial carcinoma. Nevertheless, it should be noted that “aberrant” p63 expression can be seen in some tumor cells having little or no myoepithelial differentiation, such as columnar cells in canalicular adenoma, intercalated ductal cells in acinar cell carcinoma, and a few nonspecific glandular cells in adenocarcinoma, NOS. Notwithstanding these exceptions, p63 expression seems related to the fact that salivary gland tumor tissues retain the compartmentalization and differentiation seen in their normal counterpart.
Despite striking sequence similarities and conservation of functional domains among p53 family members, p63 is not a classical tumor suppressor gene (Yang et al. 2002). The p63 gene is located in a region 3q27–29 ter that is actually amplified in squamous cell carcinoma (Bockmuhl et al. 1996; Hibi et al. 2000). Whereas the TA isoforms are capable of transactivating p53 target genes, the ΔN variants may function as a dominant-negative factor that blocks the transcription-activating ability of TAp63, p53, or the related gene p73 (Yang et al. 1998). Therefore, ΔNp63 may function as an oncogene (Hibi et al. 2000). We are now investigating which forms of p63 are expressed in normal and tumor salivary gland tissues.
Mesenchymal elements, including fibroblasts, myofibroblasts, endothelial cells, and smooth muscle cells, have undetectable p63 (Yang et al. 1998; Barbareschi et al. 2001; Di Como et al. 2002). The corresponding soft tissue tumors do not display p63 immunoreactivity. p63 staining may be helpful to distinguish spindle-cell myoepithelioma and myoepithelial carcinoma from mesenchymal tumors. However, some malignant peripheral nerve sheath tumors and a subset of rhabdomyosarcoma also express p63 (Di Como et al. 2002). p63 immunoreactivity may also help in the identification of myoepithelial cells in salivary gland aspirates, although these cells are clearly not specific for a tumor type.
In conclusion, p63 is expressed in the nuclei of normal human salivary gland myoepithelial and basal duct cells as well as the modified myoepithelial and basal cells of human salivary gland tumors. A better characterization of salivary gland myoepithelial cells may provide valuable information regarding maintenance of this tissue, histogenesis and oncogenesis of salivary gland tumors, and may have clinical utility for the diagnosis.
Footnotes
Acknowledgements
We thank Edith Joly, Annette Lesot, and Nicole Vignot for excellent technical assistance.