
Abstract
Kidney androgen-regulated protein (Kap) is the most abundant protein in the mouse kidney, but its function is unknown. We previously observed a significant decrease in Kap mRNA expression in whole kidney tissue from male mice with adenine phosphoribosyltransferase (APRT) deficiency and 2,8-dihydroxyadenine (DHA) nephrolithiasis. The disease phenotype is more severe in male mice and is age-dependent. To identify the cellular basis for differential Kap expression, we used in situ hybridization (ISH) and reverse transcription-polymerase chain reaction ISH (RT-PCR ISH) to identify the cell types expressing this mRNA in paraffin-embedded kidney sections. In 1-month-old wild-type male mice, Kap was detected primarily in S3 proximal tubule segments, but expression was very low in female mice. In 1-month-old APRT-deficient male mice, Kap expression was decreased significantly and was undetectable in female mice. Kap mRNA was not detected in 3- or 6-month-old mice using our standard ISH protocol, but we observed intense cytoplasmic staining in S3 proximal tubules in wild-type male mice of these age groups using an improved RT-PCR ISH procedure. Our studies demonstrate age-, gender-, and APRT genotype-dependent changes in Kap mRNA expression in mouse kidney. Kap expression is under multihormonal control, and hormonal changes in DHA-induced nephrolithiasis may account for the decreased Kap expression in APRT-deficient mice.
Keywords
T
As a first step in understanding the role of Kap in DHA-induced nephrolithiasis, we examined, by in situ hybridization (ISH) and RT-PCR ISH, the expression of Kap mRNA in mouse kidney sections. These findings were then correlated with the renal histological changes in these mice. Here we show that there are age-, gender-, and APRT genotype-related differences in Kap mRNA expression. Decreased Kap expression in APRT-deficient mice may be due, at least in part, to DHA-induced hormonal changes.
Materials and Methods
Preparation of Kidney Sections
All animal studies were conducted in accordance with the NIH Guide for the Care and Use of Laboratory Animals. Strain 129 mice were used in these studies and 6–8-μm thick kidney sections were prepared as described previously (Evan et al. 2001). Two sections mounted side by side on Superfrost/Plus glass slides (Fisher Scientific; Pittsburgh, PA) were used for the studies described below.
In Situ Hybridization
Tissue sections were deparaffinized, rehydrated, equilibrated in 50 mM Tris-HCl buffer (pH 7.5), and then incubated with proteinase K (4 μg/ml) for 5–10 min at 37C in a humid chamber. The digestion conditions were optimized to allow probe access to intracellular mRNA but still maintain tissue integrity (Nuovo 2001). For mRNA localization studies, we used synthetic oligonucleotides rather than cRNA or cDNA as hybridization probes, because oligonucleotides have been shown to give results comparable to those obtained with longer probes (Wisden and Morris 1994). Forty-mer oligonucleotide probes were synthesized by our DNA Core Facility. The sequences of the sense and antisense probes were 5'-CGCCAGCCGCAAACAAACAGAATTCTGAGTTCTCTACTGA3’ and 5'-TCAGTAGAGAACTCAGAATTCTGTTTGTTTGCGGCTGGCG-3', respectively. The probes were labeled at the 3'-end with digoxigenin-dUTP/dATP using an oligonucleotide tailing kit (Roche Molecular Biochemicals; Indianapolis, IN). The labeling efficiency and probe concentration were checked by dot-blot hybridization as described in the manufacturer's instructions.
Tissue sections were pre-hybridized at 42C for 1 hr using ISH buffer (Ambion; Austin, TX), and then hybridized with digoxigenin-labeled probes for 16–18 hr at the same temperature. The samples were sealed with Hybaid EasiSeals to prevent evaporation (Hybaid; Franklin, MA). The slides were washed (37C, 5 min) once in 2 X in situ wash solution (Ambion) and then twice in 1 X concentration of the same solution. For each tissue, adjacent sections were hybridized with the antisense and sense Kap probes. The probe concentration was 5–45 ng/μl (in a total volume of 60 μl) depending on message abundance, but we routinely used a probe concentration of 15 ng/μl. In addition to the sense probe, controls included omission of probe and omission of anti-digoxigenin antibody from the reaction mixture.
The signal was detected as described in the manufacturer's instructions (Roche Molecular Biochemicals). Briefly, after post-hybridization washes, the slides were washed in washing buffer for 1–5 min and then immersed in blocking buffer for 30 min. The sections were incubated in a 1:300 dilution (in blocking buffer) of alkaline phosphatase-conjugated anti-digoxigenin antibody (sheep Fab fragments) for 2 hr at ambient temperature. The slides were then incubated in nitroblue tetrazolium (NBT) and 5-bromo-4-chloro-3-indolylphosphate (BCIP) at 37C for 15–30 min, and the signal visualized as a purple precipitate. The slides were not routinely counterstained. To verify the identity of positively staining structures, selected sections were counterstained with methyl green and adjacent sections were routinely stained with hematoxylin and eosin.
The slides were examined using a Nikon Eclipse E800 microscope (Nikon; Melville, NY). For each section, five frames in a cross-configuration were captured using a X20 objective lens and a MagnaFire camera and software (Optronics; Goleta, CA). Images were processed using Scion Image for Windows (Scion; Fredrick, MD), a freely available version of NIH Image software (http://www.scioncorp.com and http://rsb.info.nih.gov/nih-image/). For each frame, 10 measurements of signal intensity were captured, again in a cross-configuration. Kidney sections from age- and sex-matched wild-type mice were used as controls. The results presented here are based on expression analyses in at least two sections from each of three to five animals of a given APRT genotype and gender. Morphometric measurements were also carried out using Scion Image software, and the data for wild-type and mutant mice were compared using the unpaired t-test.
RT-PCR ISH
We used indirect in situ RT-PCR (here referred to as RT-PCR ISH) for the cellular localization of Kap mRNA when this message could not be detected using our standard ISH protocol. RT-PCR ISH involves the incorporation of unlabeled nucleotides during PCR, followed by ISH using digoxigenin-labeled internal oligonucleotide probes (Chow et al. 1995; Teo and Shaunak 1995; O'Leary et al. 1996; Mee et al. 1997; Peters et al. 1997; Lee et al. 1998; Uhlmann et al. 1998; Walker et al. 1998; Zhou et al. 2000; Steel et al. 2001). The method thus provides increased hybridization specificity in addition to increased sensitivity. Tissue sections were digested with proteinase K as described above. We used the “One Step and One Buffer” protocol for in situ RT-PCR with the ExpeRT-PCR kit (Hybaid) and the GeneAmp PCR System 1000 (Perkin Elmer; Foster City, CA) according to manufacturer's instructions. The Hybaid kit contains AMV reverse transcriptase for the first-strand cDNA synthesis step and Taq/Pwo polymerases for the PCR amplification step.
The reaction mixture (50 μl per section) contained 5 μl of 10 X ExpeRT-PCR buffer (with MgCl2), 1 μl DMSO, 1 μl dNTP mix (final concentration 200 μm each), 300 nM forward and reverse Kap primers, 20 U RNase inhibitor, 15 U AMV reverse transcriptase, and 5 U Taq/Pwo mix. The sequences of the forward and reverse PCR primers were 5'-CTCAGTCTTTGACCAACTGGG-3’ and 5'-CTGTGATGTCTGTGTTCTCAG-3', respectively. cDNA synthesis was carried out at 42C for 1 hr. The sections were then denatured at 90C for 1 min, followed by PCR amplification at 90C for 30 sec, 48C for 30 sec, and 70C for 1 min for six to ten cycles (Mee et al. 1997). A full set of slides (10 in the case of PCR System 1000) was processed in each run, even when the actual number of samples was less than this. The following negative controls were routinely used: (a) omission of reverse transcriptase; (b) omission of the two PCR primers; and (c) omission of the polymerase mix. After PCR, the slides were washed twice in 2 X SSC and then processed for hybridization as described above for ISH.
Results
In Situ Hybridization
We detected Kap mRNA expression primarily in the S3 segments of proximal tubules and, to a lesser extent in the S1/S2 segments, in 1-month-old wild-type male mice (Figure 1A). There was a significant reduction in expression in 1-month-old APRT-deficient male mice (Figure 1B). Kap mRNA was detected mainly in S3 segments in 1-month-old wild-type female mice but expression levels were very low compared with male mice, and were undetectable in APRT-deficient female mice. We did not detect Kap mRNA in kidney sections from 3- or 6-month-old male or female mice of any APRT genotype using our standard ISH protocol (probe concentration 15 ng/μl), or even when the probe concentration was increased to 45 ng/μl.
To determine whether the decrease in Kap expression in proximal tubules was the result of renal histological changes, we measured the number of proximal tubule sections per field, the number of nuclei, the number of glomeruli, and wall thickness of proximal tubules in kidney sections from 1-month-old wild-type and APRT-deficient male mice. Five X20 non-overlapping fields were counted in sections from each of three animals per group. There was no significant difference in the mean number of tubule sections or the mean number of nuclei between wild-type and APRT knockout mice. In APRT knockout mice, the number of glomeruli per field was approximately one half and the wall thickness of proximal tubules was approximately twofold compared with wild-type mice, and both these differences were highly significant (Table 1).
We did not observe any nonspecific hybridization as demonstrated by the use of the corresponding sense probe, and this was confirmed using two additional indicators of hybridization specificity. First, no hybridization signal was observed using a mixture of unlabeled and labeled Kap antisense probes (in the ratio 100:1), indicating that the signal from the labeled probe was competed out by the unlabeled probe. Second, an antisense probe designed to hybridize to a different region of Kap mRNA was used and the hybridization pattern was identical to that obtained with the standard probe (data not shown).
RT-PCR ISH
The lack of Kap mRNA signal in mice older than 1 month may simply be due to the low sensitivity of standard ISH procedures. However, the application of several RT-PCR ISH protocols to kidney sections resulted in extensive tissue damage and loss of amplified material. This may possibly be due to extreme temperature fluctuations during the large number of PCR cycles that are typically used in these procedures and/or to uneven heat distribution over the slides (O'Leary et al. 1996; Mee et al. 1997). We therefore used an improved RT-PCR ISH procedure in which the cycle number is kept low and a full set of slides is processed in each run.
We observed intense cytoplasmic staining primarily in S3 proximal tubule segments in 3-month and older wild-type male mice. Furthermore, a probe concentration of 1–2 ng/μl was sufficient for signal detection, and the signal intensity was comparable to that observed with 15 ng/μl probe concentration in the standard ISH assay, confirming specific amplification of Kap mRNA (Figure 2A). The signal was detectable with both the sense and antisense Kap probes, again demonstrating that cDNA synthesis had occurred. RT-PCR ISH controls were negative, demonstrating the specificity of the hybridization reaction (Figure 2B), and no signal was detected in the absence of RT-PCR, even at a probe concentration of 45 ng/μl (Figure 2C). Because of poor renal histology in APRT-deficient mice, there was variable nonspecific staining in kidney sections from these mice.
Discussion
Kap represents the most abundant mRNA species in the mouse kidney, but its expression pattern is more complex than that of a number of other genes (e.g., β-glucuronidase and ornithine decarboxylase) whose expression in the kidney is also induced by androgens (Meseguer and Catterall 1992; Ding et al. 1997). After androgen stimulation, all three genes are expressed in epithelial cells of proximal convoluted tubules (segments S1 and S2) of cortical nephrons, but Kap is also expressed in the epithelial cells of proximal convoluted tubules in the juxtamedullary region (S3 segments). The differential effects of androgen action on β-glucuronidase and Kap expression have been confirmed in immortalized proximal tubule cells (Ouar et al. 1998).
Kap expression in the S3 segments is under the control of the pituitary hormone thyrotropin and of estrogens in females (Meseguer and Catterall 1992), whereas expression in the S1 and S2 segments is independent of pituitary function but is dependent on thyroid hormone for maximal induction (Sole et al. 1996). The pattern of Kap protein expression and androgen regulation follows that of its mRNA (Cebrián et al. 2001). The specific spatial and temporal expression of Kap and its tight hormonal regulation suggest an important role for this protein in normal physiological processes, but the nature of this role remains to be established.
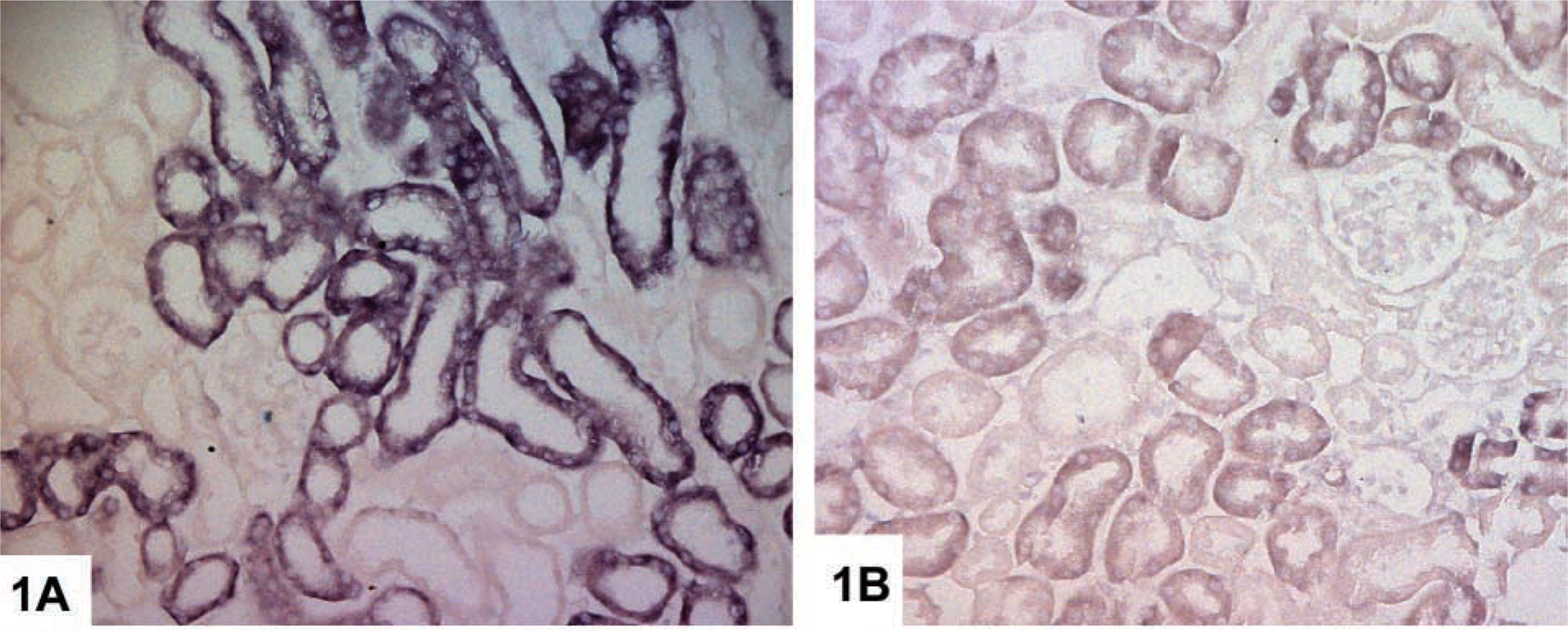
Localization of Kap mRNA in paraffin-embedded kidney sections from 1-month-old male mice by ISH. (
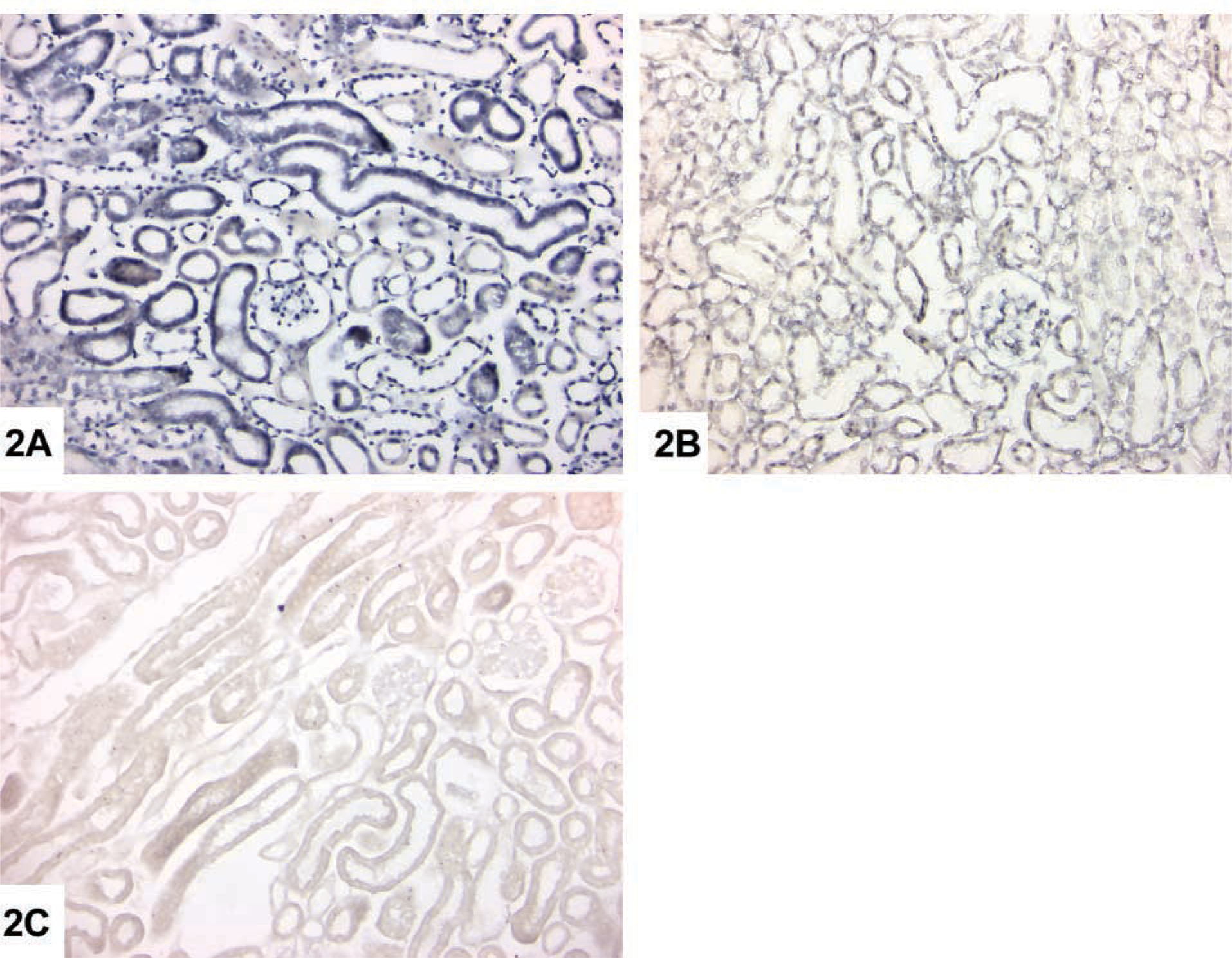
Localization of Kap mRNA in paraffin-embedded kidney sections from 3-month-old wild-type male mice by RT-PCR ISH and ISH. (
In our differential display and RT-PCR studies in APRT-deficient mice, we showed that the renal deposition of DHA leads to changes in the expression of several genes involved in tissue injury and that these changes were more pronounced in male mice (Wang et al. 2000). The events that initiate or promote kidney stone formation are not well understood, but some of the above changes may contribute to the disease phenotype in these mice (Wang et al. 2002). As a first step in understanding the potential role of Kap and other genes in disease pathology, we utilized ISH and RT-PCR ISH procedures for identifying the cell types expressing these genes.
Morphometric measurements in kidney sections from 1-month-old wild-type and APRT-deficient male micea
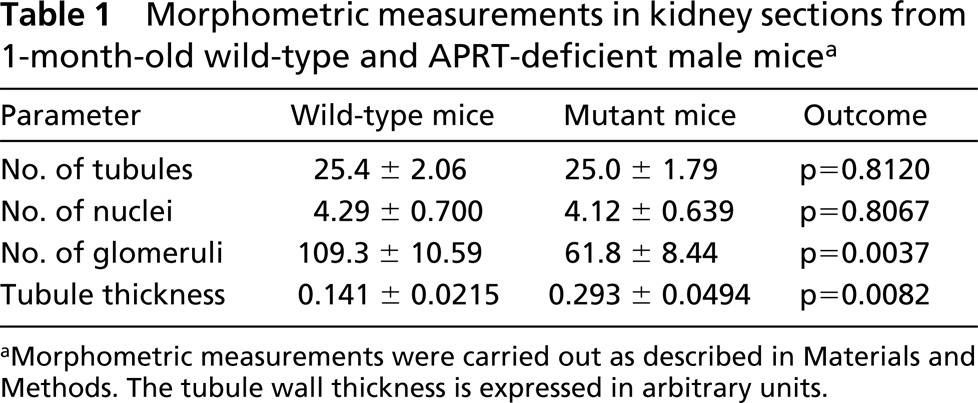
aMorphometric measurements were carried out as described in Materials and Methods. The tubule wall thickness is expressed in arbitrary units.
Our ISH studies showed that Kap mRNA was expressed primarily in the S3 segments of proximal tubules in 1-month-old wild-type male mice and that expression levels were extremely low in wild-type female mice of this age group, confirming previous studies of Kap expression in the kidney (Melantou et al. 1992; Meseguer and Catterall 1992; Meliá et al. 1998; Cebrián et al. 2001). Morphometric measurements indicated that the decreased expression of Kap in 1-month-old APRT-deficient male mice compared with wild-type controls was not the result of proximal tubule loss but may reflect the effects of hormonal changes in APRT deficiency. Androgens have a stimulating effect on cell growth through hypertrophy or hyperplasia (Preisig 1999), but in mouse kidney the primary effect appears to be hypertrophy (Ouar et al. 1998). The growth stimulation seen in proximal tubules from APRT-deficient mice is due to hypertrophy rather than hyperplasia, and this effect may again be due to hormonal changes. Glomerular loss is a known feature of APRT deficiency (Evan et al. 2001), but whether or how the decrease in glomeruli number contributes to reduced Kap expression remains to be elucidated.
There are differences in the progression of non-diabetic chronic renal disease among males and females (Silbiger and Neugarten 1995; Coggins et al. 1998; Neugarten et al. 2000). Differences in sex hormone levels have been implicated in the gender differences, possibly due to effects of these hormones on matrix accumulation, release of inflammatory cytokines, and other processes (Silbiger and Neugarten 1995). As indicated earlier, Kap expression in the kidney is under multihormonal control. We observed an age-dependent decrease in Kap expression in the S3 region and, to a lesser extent, in the S1 and S2 regions in wild-type mice, and this decrease was accelerated in APRT-deficient mice. These findings suggest that there are changes in the circulating levels of sex, pituitary, and thyroid hormones in APRT-deficient mice, and these changes may account for the reduced Kap expression in these mice.
Recent studies suggest that Kap interacts with cyclophilin B (CyPB) (Cebrián et al. 2001), which is a member of the immunophilin family and a receptor for the immunosuppressive agent cyclosporine A (CsA). CsA is also nephrotoxic and can lead to irreversible renal damage, including interstitial fibrosis (Shihab 1996; Healy et al. 1998; Lally et al. 1999). CsA administration to mice resulted in decreased Kap protein expression in the S3 segments of proximal tubules (Cebrián et al. 2001). The nature of the association between DHA nephrolithiasis and reduced Kap expression remains to be determined. Studies in knockout mice, including APRT/Kap double knockouts, are required to determine the biological basis of this association.
We did not detect Kap mRNA using our standard ISH protocol in kidney sections from wild-type or APRT-deficient mice older than 1 month. We therefore optimized an indirect in situ RT-PCR procedure (RT-PCR ISH) for detection of this mRNA. This procedure is inherently more specific than direct in situ RT-PCR, in which labeled nucleotides are included in the PCR step. RT-PCR ISH has been widely used to demonstrate the localization of low copy-number nucleic acid sequences within cells and to investigate the role of viruses in a variety of clinical conditions (Bates et al. 1997; Xiao et al. 2001). The technique also has been greatly improved to detect intracellular mRNAs in various tissue sections (Harrington and O'Leary 1998). Our studies confirm that this technique can be used to localize the site of expression of transcripts that may not be detectable with standard ISH procedures.
Previous studies have emphasized that a key parameter in RT-PCR ISH is the protease digestion step (Nuovo 2001). The success of this step depends on the type and concentration of protease, the extent of digestion, the type of tissue, the fixative used, and the duration of fixation (Martinez et al. 1995; Bates et al. 1997; Wolfahrt et al. 1998). In the present study, we used proteinase K and an empirically determined optimal digestion time. Another critical parameter in RT-PCR ISH is the attainment of correct annealing and denaturation temperatures at the level of tissue sections, where the amplification process occurs. In situ PCR machines employ specifically designed thermal blocks to optimize heat transfer from the block face to the glass slide, but even then it is essential to use a small number of PCR cycles and to process a full set of slides for the machine being used. At lower cycle numbers, problems associated with product diffusion are less likely to occur and tissue morphology is maintained. Higher cycle numbers result in extensive tissue damage, product diffusion, and nonspecific staining (O'Leary et al. 1996; Mee et al. 1997).
A comparison of Figures 1 and 2 shows that tissue morphology after RT-PCR ISH is not of the same quality as in the standard ISH procedure. Therefore, one major limitation of RT-PCR ISH is that it is unlikely to be useful when the tissue to be examined is already extensively damaged, as in the case of, for example, 6-month-old APRT-deficient male mice. In these situations, reliable comparisons of gene expression between normal and diseased tissue are best made using solution RT-PCR or ISH/ immunohistochemical procedures that amplify the detection signal rather than the target mRNA (Speel et al. 1999; Wiedorn et al. 1999; Yang et al. 1999).
Footnotes
Acknowledgements
Supported by NIH grants DK38185, ES05652, and ES06096.