
Abstract
Monoamine oxidase (MAO) oxidatively deaminates vasoactive and biogenic amines and exists in two distinct forms (A and B), coded for by separate genes, which exhibit distinct substrate specificities and inhibitor sensitivities. Using specific primers for MAO-A and MAO-B mRNA in a reverse transcription-polymerase chain reaction (RT-PCR) on RNA from human liver, the predicted products for both enzymes were detected. Furthermore, RT-PCR on RNA from human placenta, believed to contain predominantly (or only) MAO-A protein, also indicated the presence of both A and B gene transcripts. The cellular distribution of MAO mRNA in placental tissue was analyzed by in situ hybridization of MAO-A and MAO-B mRNA-specific cRNA probes on paraffin sections. MAO-A mRNA was mainly evident in the syncytiotrophoblastic layer. None was detected in the vascular endothelium/smooth muscles. Significantly, MAO-B mRNA signal was also evident in the placental villi, notably in the syncytiotrophoblasts, intermediate trophoblasts, cytotrophoblasts, and the vascular endothelium. To our knowledge, this is the first demonstration of the cellular distribution of MAO mRNA in human placenta via in situ hybridization. The expression of MAO-B in placental tissue rather than in blood elements within placenta is also unequivocally demonstrated. These highly specific cRNA probes can now be used to study the distribution of MAO-A and MAO-B expression in other tissues.
Keywords
M
MAO-A and -B are differentially expressed in a variety of tissues. Some, such as human liver and brain, contain both forms of MAO (Johnston 1968; Knoll and Magyar 1972), whereas others, such as human platelets (Bond and Cundall 1977) or placenta (Egashira and Yamanaka 1981; Weyler and Salach 1985), contain primarily one form (MAO-B and MAO-A, respectively). However, the expression of low levels of MAO-B in placenta has been reported, both as immunologically detectable protein and catalytic activity (Riley et al. 1989) and as mRNA (Bach et al. 1988). These observations are believed to be attributable to MAO-B expression in blood lymphocytes and platelets and in the vascular endothelium rather than the placental tissue itself (Riley et al. 1989). The presence of MAO-B mRNA in placental tissue is equivocal (Grimsby et al. 1990). In addition, the exact cellular localization of MAO mRNA is not known.
In this study we demonstrate the use of nonradioactively (digoxigenin, DIG)-labeled complementary RNA (cRNA) probes to specifically locate MAO-A and MAO-B mRNAs in placental sections via in situ hybridization histochemistry. The RNA probes were produced from cDNA clones encompassing the entire protein coding region (Bach et al. 1988), and their specificities were demonstrated by Northern blot hybridization. The MAO-A mRNA-specific probe reacts with MAO-A sense cRNA but not with MAO-B sense cRNA, and the MAO-B mRNA-specific probe reacts with only MAO-B sense cRNA. Thus, a clear picture of the cell types capable of expressing MAO in placenta has been obtained. Sequence-specific primers have also been used for detection of MAO-A and/or MAO-B expression by RT-PCR of RNA purified from human placenta and liver, to confirm the in situ hybridization findings.
Materials and Methods
Synthesis of the cRNA Probes
Plasmids (pSP65) carrying the appropriate cDNA (MAO-A, 2.5
Probes were subjected to limited alkaline hydrolysis in 0.2 M sodium carbonate-bicarbonate buffer (pH 10.2) and incubation at 60C for 20 min (Cox et al. 1984). The reaction was halted by lowering the pH using 3 M sodium acetate (pH 6) and 10% glacial acetic acid. The probe was precipitated with ethanol and resuspended in DEPC-treated water. Fragment length was checked as described above.
RNA Isolation
Placental tissues were obtained fresh from routine elective cesarean section deliveries performed at the Queens Medical Centre, Nottingham, UK, by Mr. G. M. Filshie and his team. The procedures used were in accordance with the ethical standards approved by the Ethics Committee, Queens Medical Centre. Total RNA was isolated using RNAStat RNA extraction solution (Biogenesis; Poole, UK). Briefly, tissues were homogenized in ice-cold RNAStat (1 ml/100 mg tissue). After the addition of chloroform to a concentration of 10% (v/v) and vigorous mixing, each sample was kept on ice for 5 min, followed by centrifugation (10,000 X g) for 20 min at 4C. The upper aqueous phase was then aspirated and mixed with an equal volume of isopropanol and placed on ice for 10 min. Total RNA was collected by centrifugation at 7500 X g for 20 min at 4C and the RNA pellet was vacuumdried after being washed twice with 75% ethanol-DEPC water. RNA was dissolved in DEPC-treated water containing RNasin (1 U/μl) and stored in liquid nitrogen.
Northern Blot Analysis
RNA samples were fractionated on 1% agarose-formaldehyde gels and capillary transferred onto nylon filters (Boehringer Mannheim) using 20 X SSC (saline-sodium citrate 1 X = 0.15 M NaCl, 0.015 M sodium citrate, pH 7.0). After washing with 6 X SSC, filters were allowed to air-dry. Then the transferred RNA was bound to the filter using a UV-translinker (Stratagene; Cambridge, UK). Marker lanes were removed and stained with 0.04% methylene blue. The rest of the filters were prehybridized in hybridization buffer (50% formamide, 4 X SSC, 7% SDS, 50 μg/ml denatured salmon sperm DNA) (Sigma; Poole, UK) at 55C for 4 hr. The positive control probe for Northern hybridization was DIG-labeled human β-actin anti-sense RNA probe (corresponding to bases 69-618 of β-actin) (Boehringer Mannheim), and the test probes were DIG-labeled MAO-A and MAO-B probes in the sense and anti-sense directions. The probe concentrations used were 80 ng/ml for the actin probe and 100 ng/ml for the MAO probes. Filters were then incubated in hybridization solution containing the designated probe at 55C overnight and washed as follows: 2 X SSC, 0.1% SDS for 15 min at 55C; 0.5 X SSC, 0.1% SDS for 15 min at 55C and 0.1 X SSC, 0.1% SDS for 30 min at 65C. Hybridization was detected immunologically. Filters were blocked by immersion in TBS (Tris-buffered saline: 100 mM Tris-HCl, 150 mM NaCl, pH 7.5) containing 2% (v/v) heatinactivated normal ovine serum and 0.3% (w/v) Triton X-100 for 30 min at room temperature (RT) and then incubated with alkaline phosphatase-conjugated sheep anti-digoxigenin (Fab fragment) (Boehringer Mannheim), diluted 1:1000 in the same buffer, for 30 min at RT. After two 15-min washes in TBS, the blots were equilibrated for 5 min in the detection buffer (100 mM Tris-HCl, 100 mM NaCl, pH 9.5). Alkaline phosphatase was visualized by the addition of 250 μM CSPD (disodium3-(4-methoxyspiro{1,2-dioxetane-3,2′-(5′-chloro)tricyclo{3.3.1.13,7]decan}-4yl) phenyl phosphate) chemiluminescent substrate (Perkin-Elmer; Warrington, UK) in detection buffer and the blots were incubated for 15 min at 37C. Signal was detected on Kodak Biomax ML film (Sigma).
In Situ Hybridization
Freshly prepared placental tissue sample blocks were fixed at RT in 4% formaldehyde in PBS for 24 hr, dehydrated, and embedded in paraffin. Sections (8 μm thick) were cut, mounted on silinated slides (Lewis and Wells 1992), dried at 45C for 1 hr, then at 37C overnight, and stored at RT. Sections were deparaffinized for in situ hybridization, rehydrated, and treated with proteinase K (10 μg/ml, 20 min at 37C), then acetylated in 0.25% acetic anhydride, 0.1 M triethanolamine-HCl, pH 8, for 10 min at RT.
Prehybridization involved incubation at 55C for 4 hr in hybridization solution (HS; 100 μl per section) containing 50% formamide, 4 X SSC, 1 X Denhardt solution (0.02% Ficoll, 0.02% polyvinyl pyrrolidone, 10 mg/ml RNase-free bovine serum albumin), 0.5 mg/ml sheared salmon sperm DNA, 0.25 mg/ml yeast tRNA, 5% dextran sulfate. After removal of the prehybridization solution, sections were hybridized by the addition of 30 μl of HS containing 0.05 μg heat-denatured probes (80C for 5 min, 4C for 10 min). Sections were covered with Parafilm, sealed with rubber cement, and incubated at 55C overnight. Hybridization fluid was then aspirated and slides washed in 2 X SSC for 5 min at RT, and 50% formamide, 2 X SSC for 30 min at 55C as the stringency wash. Filters were washed with 0.1 X SSC for 30 min at 55C to remove formamide. Signal detection was as described above, except that levamisole (0.24 mg/ml) was added to the substrate as an inhibitor of endogenous alkaline phosphatase. Color development was terminated by immersing slides in 50 mM Tris-HCl, pH 7.5, 1 mM EDTA buffer. Sections were mounted in gelatin-glycerol (Sigma), which was allowed to set at 4C.
Throughout the in situ hybridization procedure, temperature control was achieved using an Omnislide thermocycler (Hybaid).
Reverse Transcription-Polymerase Chain Reaction
Reverse transcription and PCR were performed to verify the presence or absence of MAO-A and MAO-B mRNA in hepatic and placental RNA. Sequence specific primers were selected from the full cDNA sequences of MAO-A and MAO-B (Bach et al. 1988). Primers for MAO-A were as follows: (5′ MAOA51; 3′ MAOA31), selected from exons 7 and 15, respectively. Primers for MAO-B were as follows: (5′ MAOBF1; 3′ MAOBR1), selected from exons 13 and 15 respectively, as follows:

The fragments flanked by these primers were 745
Results
Northern Hybridization
Total RNA from human liver was used to confirm the specificities of the MAO-A and MAO-B probes. Single bands of 4.9
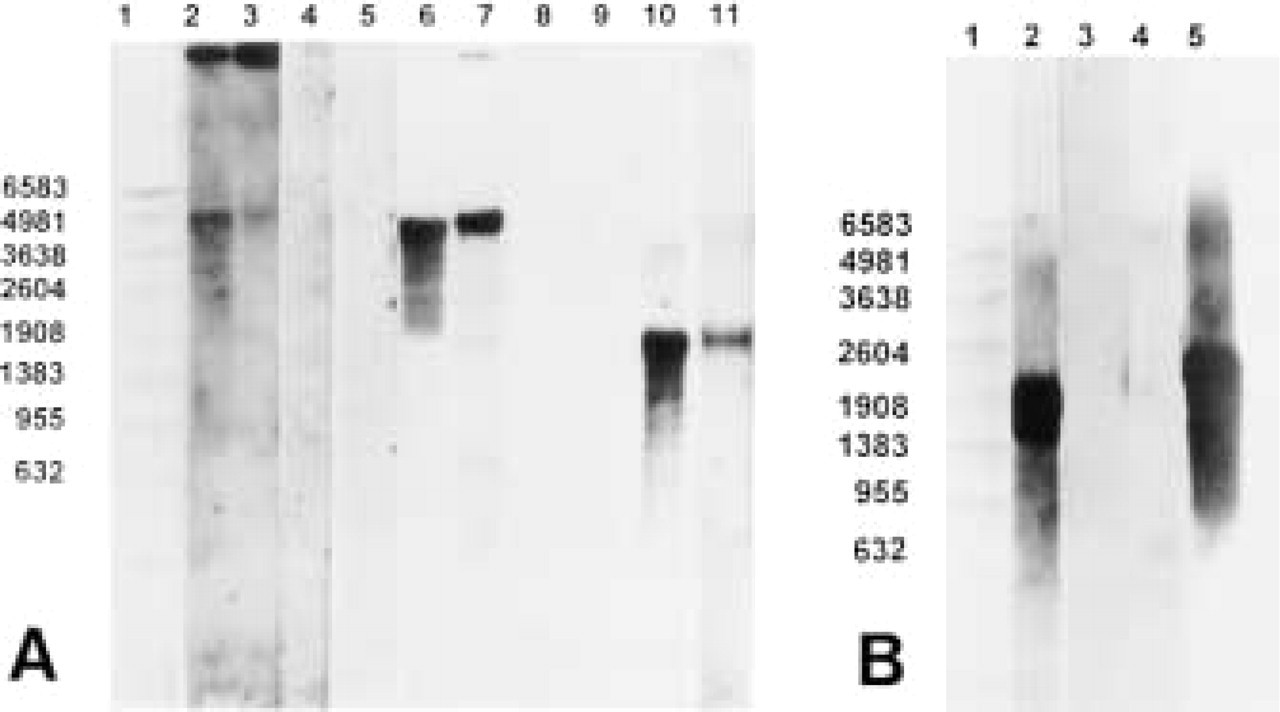
Northern blot analysis of (A) total RNA (10 μg/lane) purified from human liver (Lanes 2, 4, 6, 8, and 10) and placenta (Lanes 3, 5, 7, 9, and 11). MAO-A anti-sense probe was used in Lanes 2 and 3 and MAO-B anti-sense probe was used in Lanes 6 and 7. Lanes 4 and 5 and Lanes 8 and 9 represent MAO-A sense, and MAO-B sense, respectively. Human β-actin antisense RNA probe was used for hybridization on lanes 10 and 11.
Similar signal patterns were obtained for MAO-A and MAO-B transcripts in placental and hepatic RNA when specific digoxigenin-labeled anti-sense oligonucleotide probes were used instead of RNA probes (data not shown).
As expected, the human β-actin anti-sense DIG-labeled RNA probe revealed a single band of approximately 1.8
Reverse Transcription-Polymerase Chain Reaction
The RT-PCR revealed products from MAO-A and MAO-B genes in both liver and placenta. As predicted, a 745-
Finally, these results give further confidence in the use of the RNA probes for in situ hybridization experiments.
In Situ Hybridization
The anti-sense probes for both MAO-A and MAO-B gave signals in paraffin-embedded human liver sections, with the MAO-B mRNA predominating, and the sense probes giving no signals (data not shown).
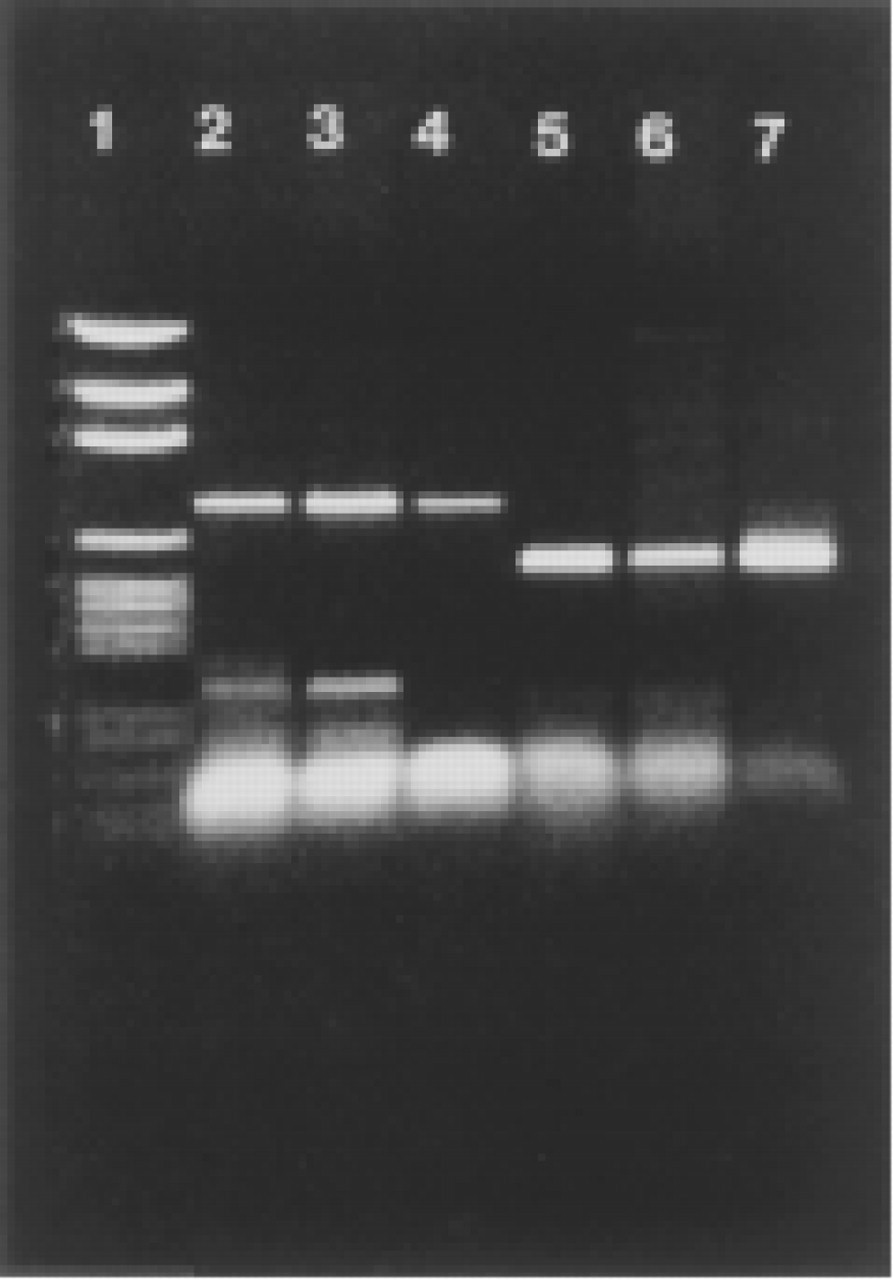
Sequence-specific MAO-A or MAO-B anti-sense primers were used for reverse transcription of human liver and placental (total) RNA. Sense cRNAs, previously transcribed from full-length MAO-A or MAO-B cDNAs (Bach et al. 1988), were used as positive controls. RT products were amplified by PCR and revealed on ethidium bromide-stained agarose gels after electrophoresis. Lanes 2—4 show the predicted 745-
In placental samples, both MAO-A mRNA and MAO-B mRNA were again detected. At low magnification, MAO-A mRNA was evident mainly on the outer (syncytiotrophoblastic) surface of villi (Figure 3A). In the stem villous trunks, a signal was observed only in the cytotrophoblastic cellular groups distributed in the stroma (Figure 3A). Very low levels of MAO-A mRNA were evident in the smooth musculature of placental vessels and the endothelial lining of the arteries and veins (Figure 4A). At higher magnification, the expression of MAO-A mRNA was mainly localized in the syncytiotrophoblastic and the cytotro-phoblastic cells of chorionic villi (Figure 4A, inset).
The use of MAO-B anti-sense probes indicated that the highest levels of MAO-B mRNA are in the syncytiotrophoblastic layer of the small and budding villi; (Figure 3C) and in the cytotrophoblastic cellular groups distributed in the villous trunk (Figures 3C and 4C). In the placental vasculature, expression of MAO-B mRNA was absent from the connective tissue surrounding blood vessels whereas, unlike that of MAO-A, there was strong expression in the smooth muscula-ture and the endothelial lining of the placental arteries and veins (Figure 4C). At higher magnifications, the expression of MAO-B mRNA in the villi was mainly localized in the syncytiotrophoblastic and cytotrophoblastic layers (Figure 4C, inset).
No hybridization was evident with the sense probes for MAO-A or -B (Figures 3B, 3D, 4B, and 4D). Posthybridization treatment with RNase also had no effect on the signal intensity or distribution; this was the case with both the sense and anti-sense probes.
Discussion
In this study we have clearly demonstrated the distribution of monoamine oxidase-A and -B mRNA in placental sections obtained from normal full-term deliveries, using in situ hybridization with DIG-labeled RNA probes. DIG-labeled cRNA probes were synthesized from MAO-A and MAO-B cDNA templates (Bach et al. 1988) in the sense (negative control) and anti-sense orientations and were partially hydrolyzed to around 400 nucleotides, using limited alkaline hydrolysis, to increase their intracellular permeation (Brahic and Haase 1978). Probe specificities were verified by Northern blot analysis. The use of unlabeled sense cRNA of either MAO-A or MAO-B as a template for Northern hybridization revealed the expected products only with their specific probes, and no hybridization signals were evident when the opposite probes were used. These findings prove that, despite the fact that the base sequences of MAO-A and MAO-B mRNAs are 50.5% homologous, (Bach et al. 1988), there was no cross-hybridization between the MAO-A probe and MAO-B mRNA or the MAO-B probe and MAO-A mRNA. Moreover, the studies on human liver RNA, which is known to contain both MAO transcripts (Grimsby et al. 1990), showed that the anti-sense probes hybridized to transcript sizes of approximately 4.9 kb for both MAO-A and MAO-B. Indeed, the products revealed when placental RNA was used for hybridization were also 4.9
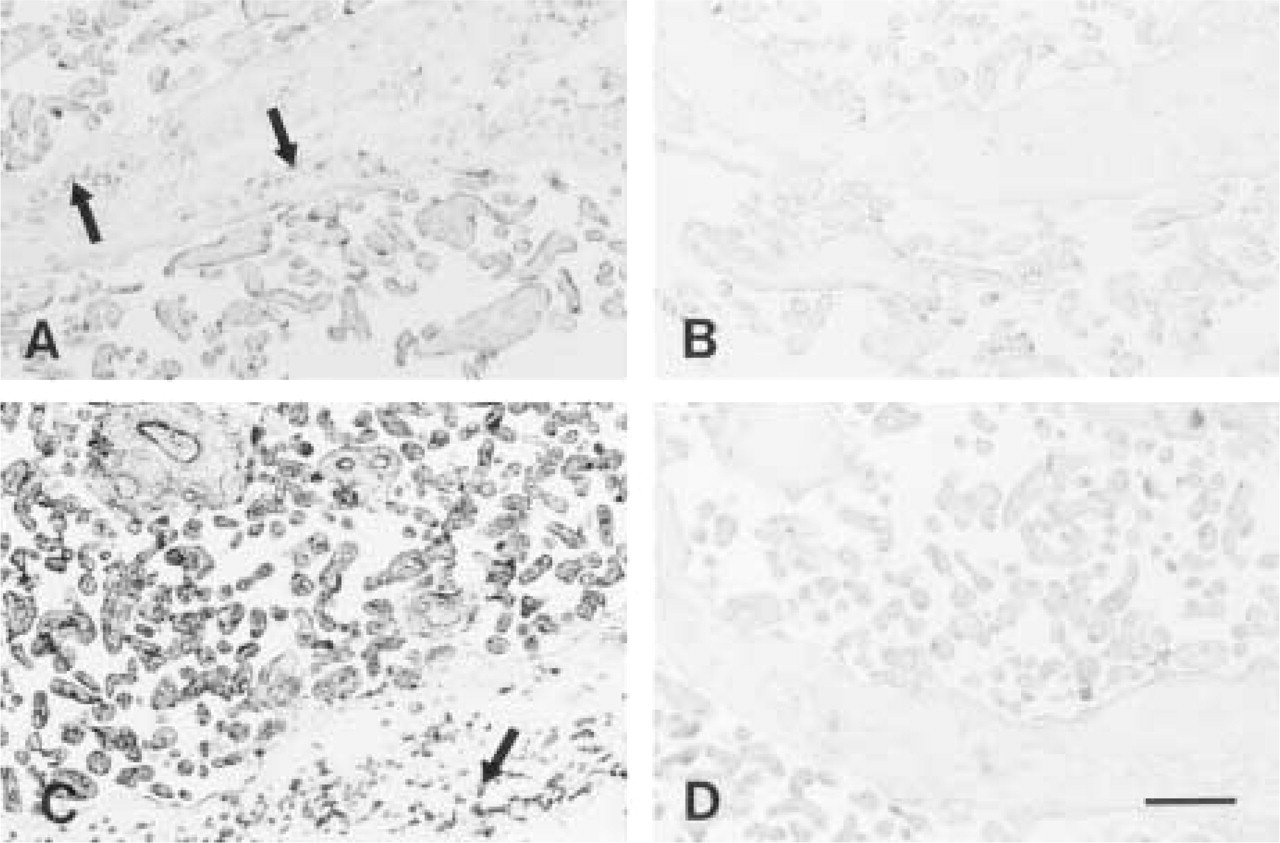
Placental sections (8 mm) hybridized with MAO-A anti-sense
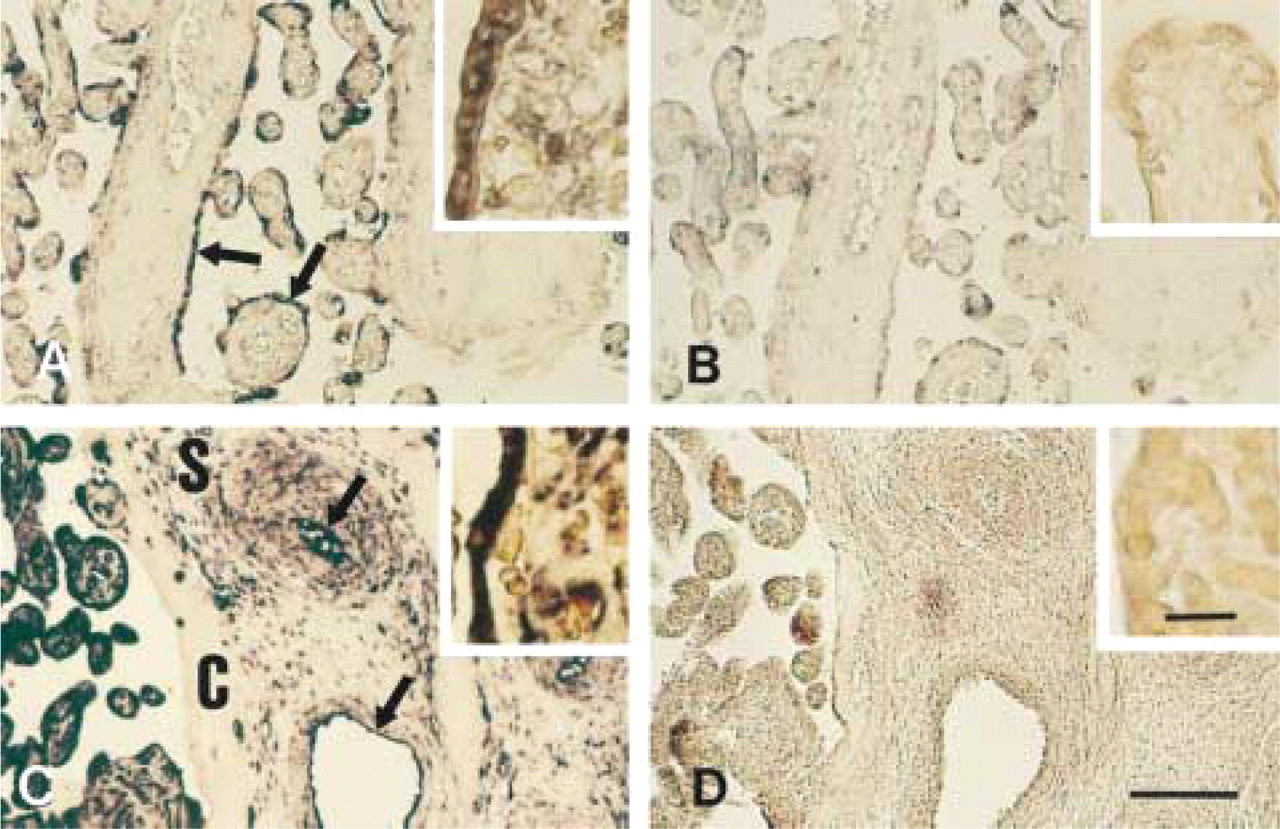
Placental sections hybridized with anti-sense
Previous work using MAO cDNA as a probe has indicated that two MAO-A transcripts occur in placenta, one of between 4.4 and 5.4
When the RNA probes were used for in situ hybridization on sections, MAO-A mRNA was easily detected in the trophoblast cells of the placental villi. This distribution agrees with earlier studies, based on enzyme catalytic activity (Yoshimoto et al. 1986) and immunohistochemical methods (Thorpe et al. 1987). Low levels of MAO-A mRNA appeared to be present also in the smooth musculature and the endothelial lining of placental arteries and veins. This is in agreement with the distribution of MAO-A protein detected using MAO-A-specific monoclonal antibodies in our laboratory (Church et al. 1994). MAO-B mRNA was seen to be highly concentrated in the trophoblast cells of the placental villi and also in the smooth musculature and the endothelial lining of placental arteries and veins. Because it is very difficult to make quantitative comparisons with different probes, the present data do not really enable one to compare the levels of MAO-A mRNA and MAO-B mRNA in placenta.
With histochemical techniques, MAO-B catalytic activity has not been detected in placental sections (Yoshimoto et al. 1986). However, low but significant MAO-B activity has been detected in placental extracts (e.g., Riley et al. 1989). In addition, using MAO-Bspecific monoclonal antibodies, Riley et al. (1989) have also reported the presence of MAO-B protein in these extracts at a level much greater than would be expected if it were derived solely from blood elements. Indeed, Riley et al. show that, under optimal conditions for their antibodies, placental mitochondria contain similar levels of MAO-A and MAO-B protein, a result considered surprising on the basis of the low level of MAO-B activity detected. However, using an MAO-B monoclonal antibody synthesized in our laboratory (Yeomanson and Billett 1992) to immunostain placental sections, we dectected only low levels of MAO-B protein (unpublished results), corroborating the results of Thorpe et al. (1987). On the other hand, our results directly show that MAO-B mRNA is indeed synthesized by cells intrinsic to the placenta (and in our sections is not present at detectable levels in the blood), supporting the suggestion that placental cells synthesize MAO-B in addition to MAO-A. Why MAO-B mRNA should be expressed at a higher level than would be anticipated from protein levels is not clear, but this may point to post-translational regulation of MAO-B expression.
Our findings clearly demonstrated that, using in situ hybridization with DIG-labeled RNA probes, it was possible to study, for the first time, the cellular distribution of MAO-A and MAO-B mRNAs in human placenta. Although in situ hybridization has recently been used to locate MAO mRNA in rat brain samples (Jahng et al. 1997), cDNA and oligonucleotide probes were used rather than the superior RNA probes employed here and, in addition, the probes were radiolabeled. Our work represents the first study using DIG-labeled RNA probes for MAO mRNA in human tissues. Our findings will be useful for further investigative studies on the expression of MAO in placenta and will, for example, allow an assessment of the role of placental MAO in conditions such as preeclamptic hypertension (Gujrati et al. 1985; Weiner 1987) and diabetes mellitus (Barnea et al. 1986). Our probes should also facilitate studies at the cellular level of the time course and regional differences in the pharmacological regulation of MAO gene expression.
Footnotes
Acknowledgments
Supported by a Grant from the Higher Education Funding Council (UK). We thank Prof Jean Chen Shih (Department of Molecular Pharmacology and Toxicology, University of Southern California) for providing the MAO-A and -B cDNA. Prof J. Lowe and Dr G. Robinson (Department of Pathology, University of Nottingham) kindly provided the tissues.