
Abstract
Morphological, contractile, histochemical, and electrophoretical characteristics of slow postural muscles are altered after hindlimb unloading (HU). However, very few data on intrafusal fibers (IFs) are available. Our aim was to determine the effects of 14 days of hindlimb unloading on the morphological and immunohistochemical characteristics of IF in rat soleus muscle. Thirty-three control and 32 unloaded spindles were analyzed. The number and distribution of muscle spindles did not appear to be affected after unloading. There was no significant difference in number, cross-sectional area, and histochemical properties of IF between the two groups. However, after unloading, a significant decrease in slow type 1 MHC isoform and a slight increase in slow-tonic MHC expression were observed in the B and C regions of the bag1 fibers. The α-cardiac MHC expression was significantly decreased along the entire length of the bag2 fibers and in the B and C regions of the bag1 fibers. In 12 muscle spindles, the chain fibers expressed the slow type 1 and α-cardiac MHC isoforms over a short distance of the A region, although these isoforms are not normally expressed. The most striking finding of the study was the relative resistance of muscle spindles to perturbation induced by HU.
T
Materials and Methods
Animals
Eight male Wistar rats (Iffa Credo; l'Arbresle, France) weighing 280–300 g were randomly divided into two groups of four rats: control rats and HU rats. Initially, all of the rats were housed in the same room at a constant temperature (25C) with a 12-hr:12-hr light–dark cycle and were fed and allowed water ad libitum. Animals in the HU group were unloaded for 14 days using Morey's model (Morey et al. 1979). Briefly, an orthopedic tape-adhesive plaster, covering less than half of the cleaned and dried tail, was connected to the top of the cage, where a swivel allowed 360° rotation. The rats were elevated in a head-down position (30°) so that the hindlimbs could not touch the cage floor or walls, while they were able to ambulate freely on their forelimbs. The experiments and the animal housing conditions received authorization from both the Agricultural and Forest in Ministry and the National Education Ministry (Veterinary Service of Health and Animal Protection, authorization 59–00980).
Tissue Processing
Rats were anesthetized with sodium pentobarbital (35 mg·kg−1). In both groups, the right and left soleus muscles were excised, stretched to their resting length, and immediately frozen in an isopentane solution precooled to its freezing point by liquid nitrogen. The muscles were stored at −80C until histochemical and immunohistochemical analyses were performed. The soleus muscles of the control and HU groups were cut into serial frozen transverse sections (10 μm thick) using a cryostat microtome (Leica CM 1800; Heidelberg, Germany) set at −20C. Along the muscle, every 280 μm, 11 sections perpendicular to the muscle longitudinal axis were performed. The first section was used as a negative control in the immunohistochemical analysis. The next two sections were processed for myofibrillar adenosine 5′-triphosphatase (ATPase) with acid (pH 4.3) and alkaline (pH 10.4) preincubations according to the method of Guth and Samaha (1969). The remaining sections were labeled with antibodies against the different MHC isoforms.
Antibodies and Labeling
Eight monoclonal antibodies (MAbs) were used in this study (Table 1). Binding of the primary antibodies was localized by an immunoperoxidase reaction utilizing the Novostain Universal Quick Kit (Tebu-Novocastra; Le Perray–en-Yvelines, France). Serial cross-sections were incubated in prediluted blocking serum (normal horse serum) for about 10 min. The excess serum was blotted and sections were incubated with primary antibodies diluted in PBS for 2 hr. Serial cross-sections were washed for 5 min in PBS. Then the sections were incubated for 30 min in prediluted biotinylated universal secondary antibody. At the end of 30 min, sections were washed with PBS for 5 min and incubated in ready-to-use streptavidin–peroxidase complex reagent for 30 min. To label the serial cross-sections, the peroxidase substrate solution (diaminobenzidine, DAB) was added after the sections had been washed for 5 min with PBS. Positive fibers were characterized by a brown color and negative fibers remained unlabeled. Finally, after dehydration with alcohol and toluene, the slides were mounted in Eukitt resin. The cross-sectional area (CSA) and the densitometry of the antibody labeling along the different IF types were measured using an image analyzer (SAMBA 2005; Villeneuve d'Ascq, France). To avoid a disparity in the labeling, for each antibody, all the sections were identically processed with the same diluted solution. The slides were also incubated simultaneously for 5 min in a common DAB solution. The zero of the densitometer for each preparation was adjusted on the negative fibers in the section; the slight variations of labeling between each slide were therefore discarded. The immunohistochemically labeled slides were analyzed with the image analyzer. This system was made up of a digital camera fixed to a standard optical light microscope. The camera was coupled to a computer with video monitor and image acquisition and storage modules. The measures were quantified to 256 gray levels (level 1 corresponding to black color and to 0% of light transmission, and level 256 to white color and 100% of light transmission), which were then converted automatically into optical density, taking into account the studied area. The densitometric values were expressed in percentage (OD = log10 (1/light transmission). According to Nibbering et al. (1986), the formation of the reaction product after DAB/H2O2 staining for immunoperoxidase-labeled cells indicated a linear relationship between the amount of enzyme-coupled antibodies bound to cells and the amount of enzyme reaction product. With the densitometric approach, the more elevated OD is, the more important the protein amount is. The densitometric analysis has already been used as a semiquantitative approach in rat muscular fibers (Sant'Ana Pereira et al. 1995). In muscle spindle immunohistochemistry, the immunoreactivity is usually scored as absent (-), weak to moderate (+), and strong (++). This way of expressing the results does not allow detection of fine modifications in the expression level of myosin isoforms under a given experimental condition. Consequently, we intentionally chose to express our results as values obtained using densitometric measurements. However, as for the ATPase activity (Kucera 1981), in the most nucleated equatorial region of the IFs, the MAb labeling was present only around the IFs because the central part of the IFs was not labeled. In this case, the densitometric measurement was made only on the colored part of IFs without taking into account the IF center.
Identification of IF Types
The muscle spindles were identified in both control and unloaded soleus muscles groups as encapsulations of small-diameter fibers. As stated previously, each muscle spindle can be divided into two encapsular regions (A and B regions) and one extracapsular portion (C region). Intrafusal fibers were classified as nuclear bag1, nuclear bag2, and nuclear chain fibers according to their morphology and their histochemical reaction after preincubation in acid and alkaline solutions (Soukup 1976; Kucera et al. 1978) and their MHC isoform contents, which were not uniform along their length (Pedrosa-Domellof et al. 1991; Kucera et al. 1992; Soukup et al. 1995). Rat muscle spindles usually contain four IFs: one bag1, one bag2, and two chain fibers (Kucera and Walro, 1988b). The diameter of these IFs decreases in the following order: nuclear bag2 > bag1 > chain fibers (Soukup 1976). Nuclear bag1 fibers exhibit low acid and alkaline ATPase activity along their intracapsular region but have high acid ATPase activity at their poles. Nuclear bag2 fibers have an acid-stable ATPase activity along their length and an intracapsular alkaline-stable ATPase activity that is lost beyond the capsular sleeve. Nuclear chain fibers have low acid and high alkaline ATPase activities along their entire length (Kucera et al. 1978; Khan and Soukup 1988). The nuclear bag1 and bag2 fibers were labeled with both ALD58 MAb, specific for the slow-tonic MHC isoform, and F88 MAb, specific for the α-cardiac MHC isoform. Nuclear chain fibers did not express these two MHC isoforms, except in a very short equatorial length where the slow-tonic isoform was detected (Pedrosa et al. 1989; Pedrosa-Domellof et al. 1991).
Statistical Analysis
All results were expressed as means ± SD. A Student's t-test was used to establish the intergroup comparisons and statistical significance was accepted at p<0.05.
Results
A total of 33 control muscle spindles (33 bag2, 33 bag1, 69 chain fibers) and 32 HU muscle spindles (32 bag2, 32 bag1, 67 chain fibers) were analyzed throughout their entire length. Most of the muscle spindles in the control and unloaded groups contained one bag1, one bag2, and two chain fibers. However, in three control and four HU muscle spindles, there were three nuclear chain fibers. There was no significant different between the number of muscle spindles in control (14.3 ± 1.5) and HU groups (13.5 ± 1.3).
Antibodies used in this study and their specificity in the rat
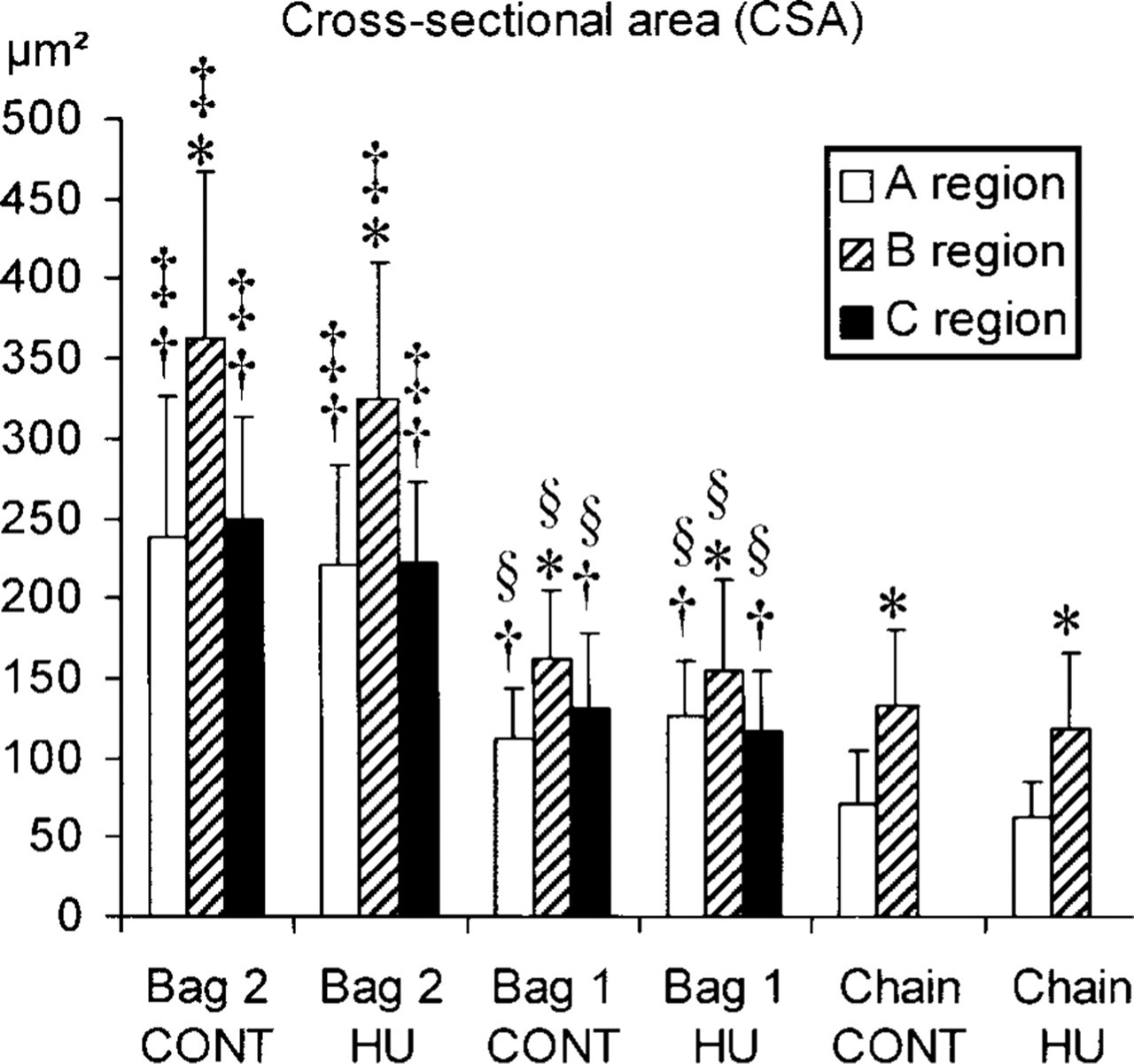
Histogram of cross-sectional areas (CSAs) of the three intrafusal fiber types in A, B, and C regions of the control (CONT) and hindlimb unloaded (HU) groups. Empty bar, A region; hatched bar, B region; full bar, C region. ∗Significant differences of CSA with the A region, †;with the B region, §with bag1 and chain fibers, and §with chain fibers.
Cross-sectional Areas
The CSAs of IFs are shown in Figure 1. For both control and HU groups, the CSAs of bag2 fibers were significantly larger than those of bag1 and chain fibers, and those of bag1 fibers were higher than those of chain fibers, independent of the muscle spindle region. For all IF types in both animal groups, the CSAs in the B region were significantly higher than those in the A and C regions (Figure 1). After 14 days of unloading, no significant difference in the CSAs of all IF types between control and HU groups was observed.
Transverse sections (10 μm thick) through the encapsulated A and B regions and extracapsular C region of a rat soleus spindle in the control group. The spindle contains one bag1 (thin arrow), one bag2 (thick arrow), and two chain fibers (star). Sections were processed for ATPase labeling with acid (pH 4.3,
Histochemical Analysis
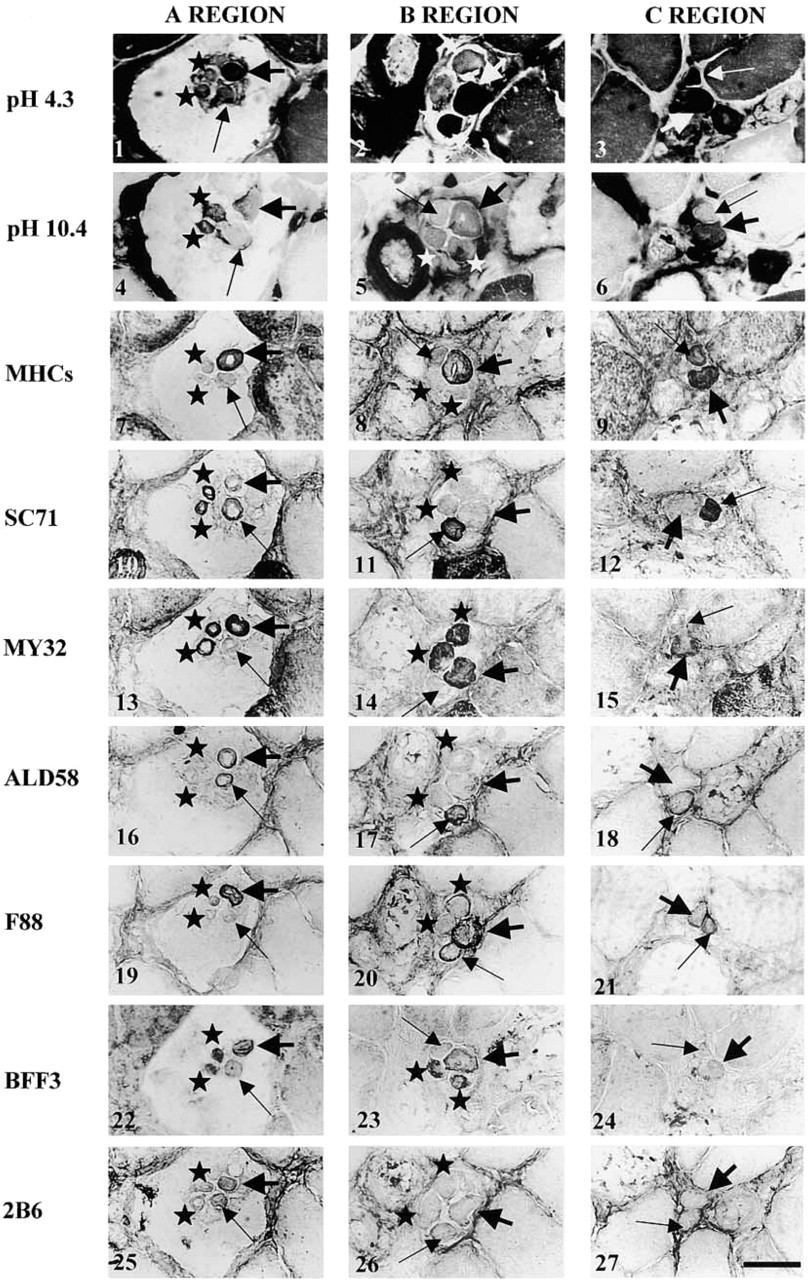
In Figure 2 (control group), the Figures 2.1–2.3 and Figures 2.4–2.6 respectively illustrative the acid (pH 4.3) and alkaline (pH 10.4) ATPase activity of IFs in the A, B, and C regions of a muscle spindle. In the muscle spindles of the control group, the nuclear bag1 fibers had low to moderate acid ATPase activity in A and B regions, but the activity was high at the end of the B region and towards the poles. After alkaline preincubation, they showed a low ATPase activity along their entire length. Nuclear bag2 fibers exhibited high acid ATPase activity and high to moderate alkali ATPase labeling along their length. However, in the more extracapsular region, the nuclear bag2 fibers lost their alkaline ATPase activity. In general, after acid preincubation, nuclear chain fibers exhibited low ATPase activity and high to moderate alkali ATPase labeling along their entire length. However, the nuclear chain fibers in the muscle spindles of Figure 2 exhibited moderate acid ATPase activity in the A region. In the most equatorial region, all IF types had a rim or no ATPase activity. Histochemical labeling did not show any difference between control and HU groups. However, it is obvious that a slight increase or decrease in one of the MHC isoforms would not necessarily be made visible by changes in ATPase activity. Moreover, ATPase labeling does not allow demonstration of the co-expression of several MHC isoforms within a fiber.
Immunohistochemistry
The regional variation of labeling with all antibodies is illustrated in Figure 2 for the control group. Figure 3 illustrates only the antibody labelings that were changed by the HU condition. The results have been refined by densitometric measurements (Table 2, control group and Table 3, HU group).
Control Group
Nuclear Bag1 Fibers. NCL MHCs (Figures 2.7–2.9) labeled only the fibers towards the poles at the end of the B region and in the C region. SC71 (Figures 2.10–2.12) and ALD 58 MAbs (Figures 2.16–2.18) strongly labeled the fibers in A and B regions and the labeling slightly decreased towards the C region (Table 2). The nuclear bag1 fibers were labeled with F88 MAb (Figures 2.19–2.21) over a short distance in the B and C regions. The binding with F88 MAb was strong to moderate in the B region and low to moderate towards the C region. The labeling with 2B6 MAb (Figures. 2.25–2.27), specific for embryonic MHC, was low in the A region. No binding was seen in the B and C regions of bag1 fibers. Whatever the IF MY32 (Figures 2.13–2.15), BFF3 (Figures 2.22–2.24), NCL MHCd MAbs did not label the nuclear bag1 fibers (not shown).
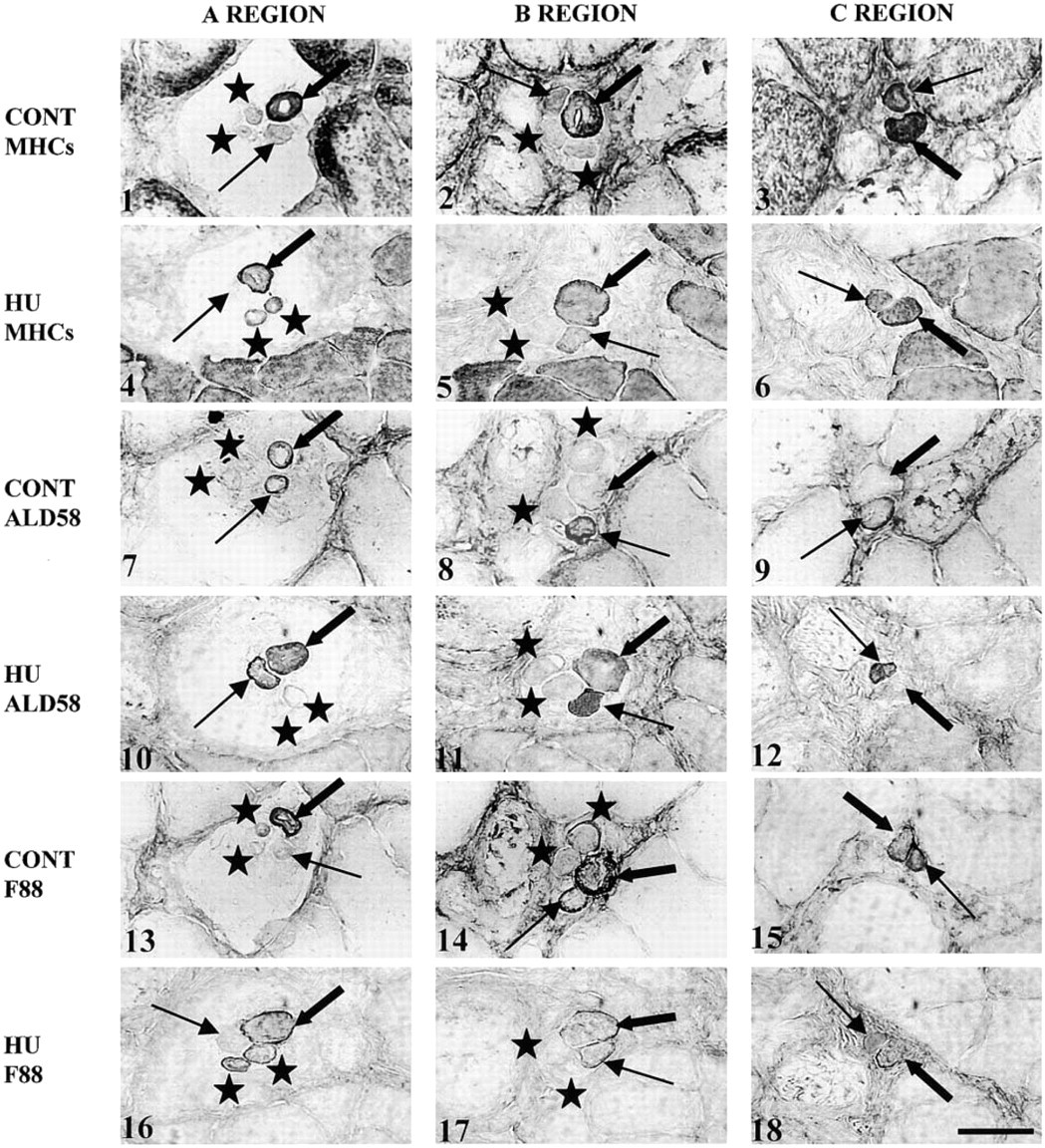
Transverse sections (10 μm thick) through the encapsulated A and B regions and extracapsular C region of a rat soleus spindle in the control (CONT) and in the hindlimb unloaded (HU) groups. The spindle of the HU group contains one bag1 (thin arrow), one bag2 (thick arrow), and two chain fibers (star). Sections were labeled with NCL MHCs named MHCs in the figure (CONT:
Densitometric analysis of the immunohistochemical labeling in intrafusal fibers of the control group (CONT)
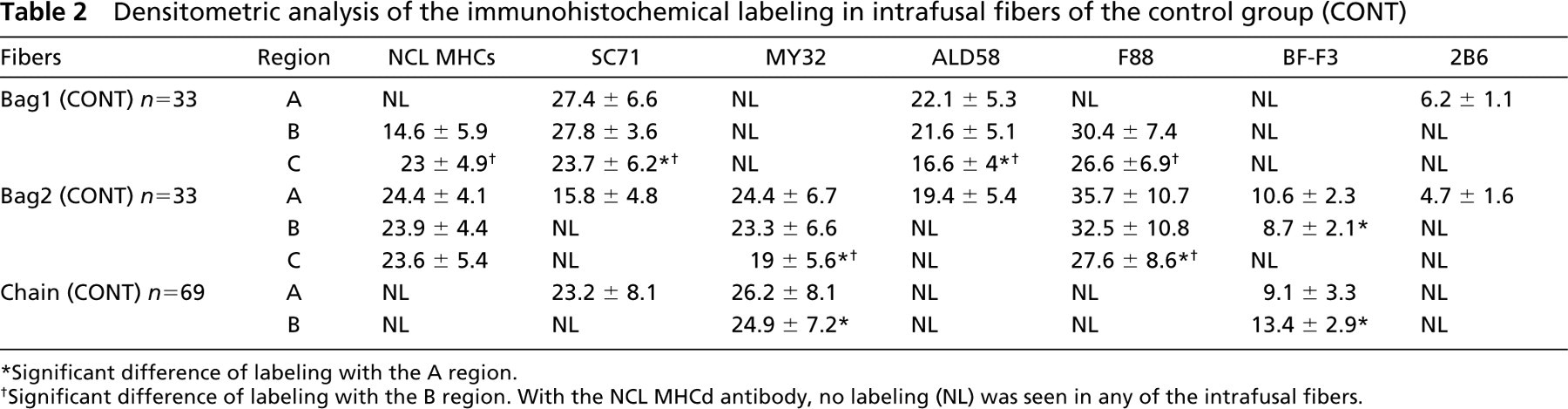
∗Significant difference of labeling with the A region.
†Significant difference of labeling with the B region. With the NCL MHCd antibody, no labeling (NL) was seen in any of the intrafusal fibers.
Nuclear Bag2 Fibers. NCL MHCs MAb reacted strongly with the nuclear bag2 fibers along their entire length. The nuclear bag2 fibers bound MY32 and F88 MAbs along their entire length but the binding intensity decreased from the A to the C region (Table 2). With SC71, ALD58, and 2B6 MAbs, the nuclear bag2 fibers were labeled only in the A region. BF-F3 MAb reacted in both A and B regions of bag2 fibers; the labeling was lower in the B region. NCL MHCd MAb did not label bag2 fibers.
Nuclear Chain Fibers. Nuclear chain fibers reacted with MY32 MAb in the A and B regions. BF-F3 labeled the nuclear chain fibers in the A and B regions, with higher labeling in the B region (Table 2). SC71 MAb bound only the A region of nuclear chain fibers. Our results showed no labeling with NCL MHCs, ALD58, F88, 2B6, and NCL MHCd MAbs in any region of the nuclear chain fibers.
HU Group. There was no difference with the control group in the regional variation and in the labeling intensity with SC71, MY32, BF-F3, 2B6, and NCL MHCd MAbs along the nuclear bag1 and bag2 fibers.
Nuclear Bag1 Fibers. NCL MHCs, ALD58, and F88 MAbs (Figures 3.4–3.6, 3.10–3.12, and 3.16–3.18, respectively) showed modified labeling compared to the control group (Figures 3.1–3.3, 3.7–3.9, and 3.13–3.15, respectively). Densitometric analysis, presented in Table 3, showed that type 1 MHC expression in nuclear bag1 fibers decreased significantly in both B and C regions after unloading. Moreover, the labeling intensity with ALD58 MAb, specific for slow-tonic MHC isoform, was significantly increased in the B and C regions of bag1 fibers. With F88 MAb, our results showed that the expression of α-cardiac MHC isoforms in bag1 fibers was significantly decreased in the B and C regions after unloading.
Nuclear Bag2 Fibers. After unloading, the labeling intensity with F88 MAb significantly decreased in all regions of the nuclear bag2 fibers compared with the control group. There was no difference with the control group in the regional variation and the labeling intensity with ALD58 MAb along bag2 fibers. However, although the difference was not significant, type 1 MHC isoform expression presented a slight decrease in the B and C regions of the nuclear bag2 fibers after a period of unloading.
Nuclear Chain Fibers. There was no difference with the control group in the regional variation and labeling intensity along the nuclear chain fibers with all antibodies, except for NCL MHCs and F88 MAbs. Indeed, among the 32 muscle spindles that were studied, only 12 presented nuclear chain fibers that bound the NCL MHCs and F88 MAbs (Figures 3.4 and 3.16, respectively) over a short distance of the A region (special chain in Table 3), whereas chain fibers in the control group never bound these antibodies.
Densitometric analysis of the immunohistochemical labeling in intrafusal fibers of the HU group a

aDensitometric analysis presented in percentage of bag1, bag2, and chain fibers in the HU group. Values are given for each antibody in the three different muscle spindle regions (A, B, C).
§Significant difference of labeling with control group.
∗Significant difference of labeling with the A region.
†Significant difference of labeling with the B region. With the NCL MHCd antibody, no labeling (NL) was seen in any of the intrafusal fibers. The chain fibers labeled with NCL MHCs and F88 in 12 muscle spindles are called special chain fibers.
Discussion
Morphological Characteristics
The number and distribution of muscle spindles in the rat soleus were not significantly different in the two experimental groups. These data are in agreement with those described previously by Soukup et al. (1990b). Our results showed that, in the control group, the bag2 fiber CSAs were greater than the bag1 fiber CSAs, which were also larger than the chain fiber CSAs. These data are in agreement with the results of other authors (Botterman and Edgerton 1975; Soukup 1976; Soukup et al. 1993). The same results were obtained after unloading. It therefore appears that IFs are more resistant to myogenic atrophy than the extrafusal fibers, as suggested earlier by other authors (Yellin and Eldred 1970; Maier et al. 1972).
Histochemical Analysis
In the control group, our data are similar to those obtained by other authors for the same species (Botterman and Edgerton 1975; Soukup 1976; Kucera et al. 1978; Kucera and Walro 1987). In some control muscle spindles, the nuclear chain fibers have moderate acid-stable ATPase activity. Soukup (1976) has also reported some differences in ATPase labeling. In the rat soleus muscle spindles, the nuclear chain fiber ATP-ase activity was low, medium, or high after acid preincubation.
After a 14-day period of HU, no significant difference was observed in the ATPase labeling in the IFs compared to the control group. The regional variation in ATPase labeling along the IFs is due to the non-uniform expression of MHC isoforms in the different regions of those fibers (Kucera and Walro 1989; Pedrosa et al. 1989; Kucera et al. 1992; Soukup et al. 1995).
MHC Isoform Expression of IFs in Control and HU Groups
Control Group. Our results are in agreement with other studies concerning the regional variation of labeling with NCL MHCs (Pedrosa-Domellof et al. 1991; Kucera et al. 1992; Soukup et al. 1995; Walro et al. 1997; Wang et al. 1997), MY32 (Pedrosa et al. 1989; Kucera et al. 1992), ALD58 (Kucera and Walro 1989; Kucera et al. 1992; Soukup et al. 1995; Soukup and Thornell 1997), and F88 (Pedrosa et al. 1990; McWhorter et al. 1995).
However, some differences were observed with SC71, BF-F3, 2B6, and NCL MHCd antibodies. Our results showed that SC71 bound to the encapsulated polar region of bag1 fibers and labeling was also seen in the juxtaequatorial region in bag1 and chain fibers. Our results were in accordance with those described by Kucera et al. (1992). However, in the A region of our nuclear bag2 fibers, only a low level of labeling was seen with the SC71 MAb. This difference in the labeling with SC71 between our results and those of Kucera et al. (1992) could be due to the antibody dilution. Indeed, Kucera et al. (1992) used the SC71 with a 1:600 dilution, whereas we used a 1:20 dilution.
A difference in labeling was observed in the nuclear bag1 fibers with MY32 and SC71 MAbs. Nuclear bag1 fibers were labeled by SC71 along their entire length, whereas MY32 MAb never labeled these fibers. This was surprising because the MY32 MAb is supposed to react with all fast-twitch MHC isoforms (Schiaffino et al. 1989). SC71 MAb is specific for the MHC 2A isoform and never crossreacts with other MHC isoforms. Therefore, the fast-twitch MHC isoform, recognized in nuclear bag1 fibers by the SC71 but not by MY32, could be a specific muscle spindle MHC isoform whose epitope resembles that recognized by SC71 on the MHC 2A isoform. The possible existence of other specific muscle spindle MHC isoforms not expressed in extrafusal fibers has already been suggested by other authors (Kucera et al. 1992; Pedrosa-Domellof et al. 1993). Kucera et al. (1992) suggested that the labeling intensity and the regional variation with MY32 were higher and broader than those with SC71 (MHC 2A) and BF-F3 (MHC 2B) MAbs, indicating the possible existence of a specific fast-twitch MHC isoform expressed by the IFs but not by the extrafusal fibers.
Our results show that the BF-F3 MAb bound only the nuclear bag2 and chain fibers in the A and B regions, whereas Kucera et al. (1992) showed that these fibers were labeled only in the B region. This difference with our data could therefore be explained by the dilution (1:200) that they used, since we used a lower dilution (1:10).
Only a low level of labeling or no labeling was observed with 2B6 and NCL MHCd antibodies, respectively. These labeling differences reflected either differences in antibody affinities or the existence of more than one embryonic MHC isoform (Silberstein and Blau 1986). 2B6 gave a low level of labeling in the juxtaequatorial region of the nuclear bag fibers, and bag1 fibers were more intensely labeled than the nuclear bag2 fibers. Maier et al. (1988) observed that, with 2B6 MAb diluted at 1:50 (Gambke and Rubinstein 1984), the nuclear bag1 and bag2, but not nuclear chain fibers, were labeled in the juxtaequatorial and polar regions. However, we observed that, at this dilution, 2B6 MAb crossreacted with the fast type 2A myosin in extrafusal fibers (data not shown). We therefore used a 1:2000 dilution to remove this cross-reaction. On the other hand, Harris et al. (1989) showed that 2B6 MAb was effective at a dilution of 1:2000–1:4000 to label the embryonic MHC isoform. Therefore, the labeling observed by Maier et al. (1988) could be due to the expression of both embryonic and MHC 2A isoforms in the nuclear bag fibers. However, our results showed that nuclear chain fibers expressed the MHC 2A isoform in the A region. If the labeling pattern observed with 2B6 MAb in the nuclear bag fibers was also due to the crossreactivity with the MHC 2A isoform, in the study of Maier et al. (1988), the nuclear chain fibers should be labeled by 2B6 MAb. This is not the case. Such a surprising result could support the hypothesis that, in the nuclear bag1 fibers, the MHC isoform recognized by SC71 MAb is not the MHC 2A isoform but a specific fast-twitch isoform not recognized by 2B6 antibody. This is perhaps the reason why, in the study of Maier et al. (1988), 2B6 did not label the nuclear chain fibers at 1:50 dilution.
Kucera et al. (1992), using three different MAbs against embryonic MHC, observed either no, medium, or high-intensity labeling in IFs. Differences in antibody affinities and/or the existence of more than one embryonic MHC isoform (Silberstein and Blau 1986) cannot be ruled out. Our data showed no labeling with NCL MHCd MAb in all the IFs, which was in agreement with the results obtained by Kucera et al. (1992) using the BF-B6, another MAb specific for neonatal MHC. However, using other antibodies, it has been observed that the neonatal MHC is expressed in the nuclear bag2 and nuclear chain fibers (Pedrosa-Domellof et al. 1991; Soukup et al. 1995). Discrepancies among all these results could be related to the different specificities of the antibodies. The fact that no labeling was observed with NCL MHCd could be due to the presence of a small amount of neonatal MHC isoform that was not detected by the immunohis-tochemical method.
HU Group. All the observations on regional variation and labeling intensity in muscle spindles of the control group were also valid for the HU group. However, after unloading, some differences were observed in the expression level of slow type I, slow-tonic, and α-cardiac MHC isoforms.
The regulation of the α-cardiac MHC isoform expression along the length of bag fibers is under the influence of motor innervation. The expression of α-cardiac MHC appears one day after the arrival of bag1 and bag2 γ-motor innervation (Pedrosa et al. 1990). After neonatal de-efferentation, the reactivity of nuclear bag2 fibers to the anti-α-cardiac MHC isoform was decreased and was limited to a shorter portion of the fibers, whereas the nuclear bag1 fibers were unreactive (Pedrosa et al. 1990; Pedrosa-Domellof et al. 1991). Conversely, slow-tonic MHC isoform expression along the length of bag fibers is undoubtedly related to the presence of sensory innervation. Indeed, neonatal denervation (Soukup et al. 1990a; Kucera et al. 1993) and deafferentation (Kucera and Walro 1987,1988a,b) prevented the expression of the slow-tonic MHC isoform. However, after neonatal de-efferentation, the regional variation of this MHC isoform along the intrafusal bag2 fibers is modified and the slow-tonic MHC isoform is expressed more intensely and over most of their length (Soukup et al. 1990a; Pedrosa-Domellof et al. 1991).
In adult rat muscle spindles, deafferentation or deefferentation produced less severe alterations, but the expression pattern of some MHC isoform along the IF was modified (Wang et al. 1997). Wang et al. (1997) observed that, when adult rat muscle spindles were deefferented in the extensor digitorum longus (EDL) muscle, the nuclear bag1 fibers, which normally express the α-cardiac MHC in the outer B region, ceased to express this isoform. Moreover, bag2 fibers continued to express this isoform but less intensely than normally. Pedrosa et al. (1990) and Soukup et al. (1990a) both showed that, after denervation or neonatal deefferentation, the expression of the α-cardiac MHC was decreased and the slow-tonic expression was increased along the length of nuclear bag fibers. According to these studies, we suggest that the decrease in α-cardiac and the increase in slow-tonic MHC isoforms in nuclear bag fibers reflect a decrease in the activity pattern of the motor nerves during unloading. This hypothesis is reinforced by the fact that, when extrafusal fibers are deprived of motor innervation (Harris et al. 1989), the expression of slow type 1 MHC isoform decreases. Similarly, our results showed that the expression of slow MHC isoform significantly decreased in the nuclear bag1 fibers after a period of unloading. However, Walro et al. (1997), who used two kinds of deafferentation, concluded that regulation of MHC expression in adult EDL muscle of rat also depended on neurotrophic factors transported anterogradely from afferent neurons to the IFs.
It is more difficult to understand why slow-twitch and α-cardiac MHC isoforms are expressed over a very short distance of the A region in nuclear chain fibers after unloading. The expression of these two MHC isoforms is usually dependent on motor innervation, whose influence decreases from the C to the A region. Therefore, the expression of slow-twitch and α-cardiac MHC isoforms over only a short portion of the A region was unexpected. No previous studies on fetal, neonatal, or adult muscle spindles have reported a possible afferent or motor influence on the expression of slow-twitch and α-cardiac MHC isoforms in nuclear chain fibers.
To conclude, after unloading, the muscle spindle integrity and the labeling pattern of the majority of MHC isoforms were preserved. However, some differences were observed: a decrease in α-cardiac MHC expression and an increase in slow-tonic expression along the nuclear bag fibers. In the literature, it has been demonstrated that the level of α-cardiac MHC expression (Pedrosa et al. 1990; Soukup et al. 1990a; Pedrosa-Domellof et al. 1991) and the regional variations in slow-tonic MHC expression depended on motor innervation (Soukup et al. 1990a; Pedrosa-Domellof et al. 1991). Consequently, we hypothesized that the observed changes could be due to a decreased influence of motor innervation. In fact, it has been demonstrated that Ia afferents project not only onto α-skeletomotor neurons but also onto β-skeletofusimotor and γ-fusimotor neurons (Bernstein and Goldberg 1995). Therefore, the decrease in motor innervation activity could be due to decreased proprioceptive information. During unloading, the soleus muscle is often in a shortened position (Riley et al. 1990) and electromyographic activity is considerably reduced (Blewett and Elder 1993). As early suggested by Ohira et al. (1992), we supposed that the muscle spindles, which are stretch sensors, are probably little or not at all stimulated. Consequently, the afferent activity of Ia and II fibers originating from these stretch receptors might be reduced, as shown indirectly by Falempin and Fodili In-Albon (1999). We therefore conclude that proprioceptive feedback could regulate the expression of some MHC isoforms in adult muscle spindles, although changes in neurotrophic factors anterogradely transported from afferents to intrafusal fibers during hindlimb unloading cannot be ruled out.
Footnotes
Acknowledgements
Supported by grants from the CNES (3027) and the Conseil Régional du Nord Pas-De-Calais.
The SC-71 and BF-F3 antibodies developed by Schiaffino et al. (1989) were obtained from the Deutsche Sammlung von Mikroorganismen and Zellkulturen Gmbh (DSMZ; Braunschweig, Germany). The ALD58 antibody developed by ![]() was obtained from the Developmental Studies Hybridoma Bank (DSHB; Iowa City, IA) maintained by The University of Iowa (Department of Biological Sciences, Iowa City, IA). We thank Dr A. Kelly and Dr N.A. Rubinstein for their gift of 2B6 to Dr G.S. Butler Browne.
was obtained from the Developmental Studies Hybridoma Bank (DSHB; Iowa City, IA) maintained by The University of Iowa (Department of Biological Sciences, Iowa City, IA). We thank Dr A. Kelly and Dr N.A. Rubinstein for their gift of 2B6 to Dr G.S. Butler Browne.