
Abstract
To determine which part of the smooth muscle cells (SMCs) of the ductus arteriosus (DA) contribute to duct closure after birth, we looked for areas in which SM2 myosin heavy chain (MHC) mRNA expression, which is associated with contraction of smooth muscle, and apoptosis could be detected in the DA during development. In situ hybridization revealed that the SM2 MHC mRNA was strongly positive in the longitudinally oriented SMCs and inner layer of the circularly oriented SMCsjust before birth. Apoptotic cells were detected in the SMCs of the DA from 1 day after birth. Histochemical analysis using terminal deoxynucleotidyl transferase-mediated dUTP-digoxigenin nick end-labeling (TUNEL) revealed significant numbers of TUNEL-positive nuclei in the longitudinally oriented SMCs and the inner layer of the circularly oriented SMCs. Masson-stained sections showed that the TUNEL-positive area in the DA was replaced by connective tissue from 1 day after birth. These results suggest that the increase in the SM2 MHC mRNA expression and the induction of apoptosis are present at the same site in the media of the DA. Therefore, the SMCs in this area may play an important role in duct constriction and remodeling of the vessel wall after birth.
Keywords
T
In the blood vessel smooth muscle, at least two smooth muscle-type myosin heavy chain (MHC) isoforms, SM1 and SM2 (Babij and Periasamy 1989; Nagai et al. 1989), and two non-muscle-type MHC isoforms, NMA and NMB (Katsuragawa et al. 1989; Saez et al. 1990), have been identified. Unfortunately, functional differences among these four MHCs are not yet known. However, although there is no direct evidence, several studies have suggested that expression of the SM2 MHC isoform is closely related to the contractile phenotype of the SMCs (Imamura et al. 1992; Kim et al. 1993; Sakurai et al. 1996).
In situ hybridization of SM2 MHC mRNA (
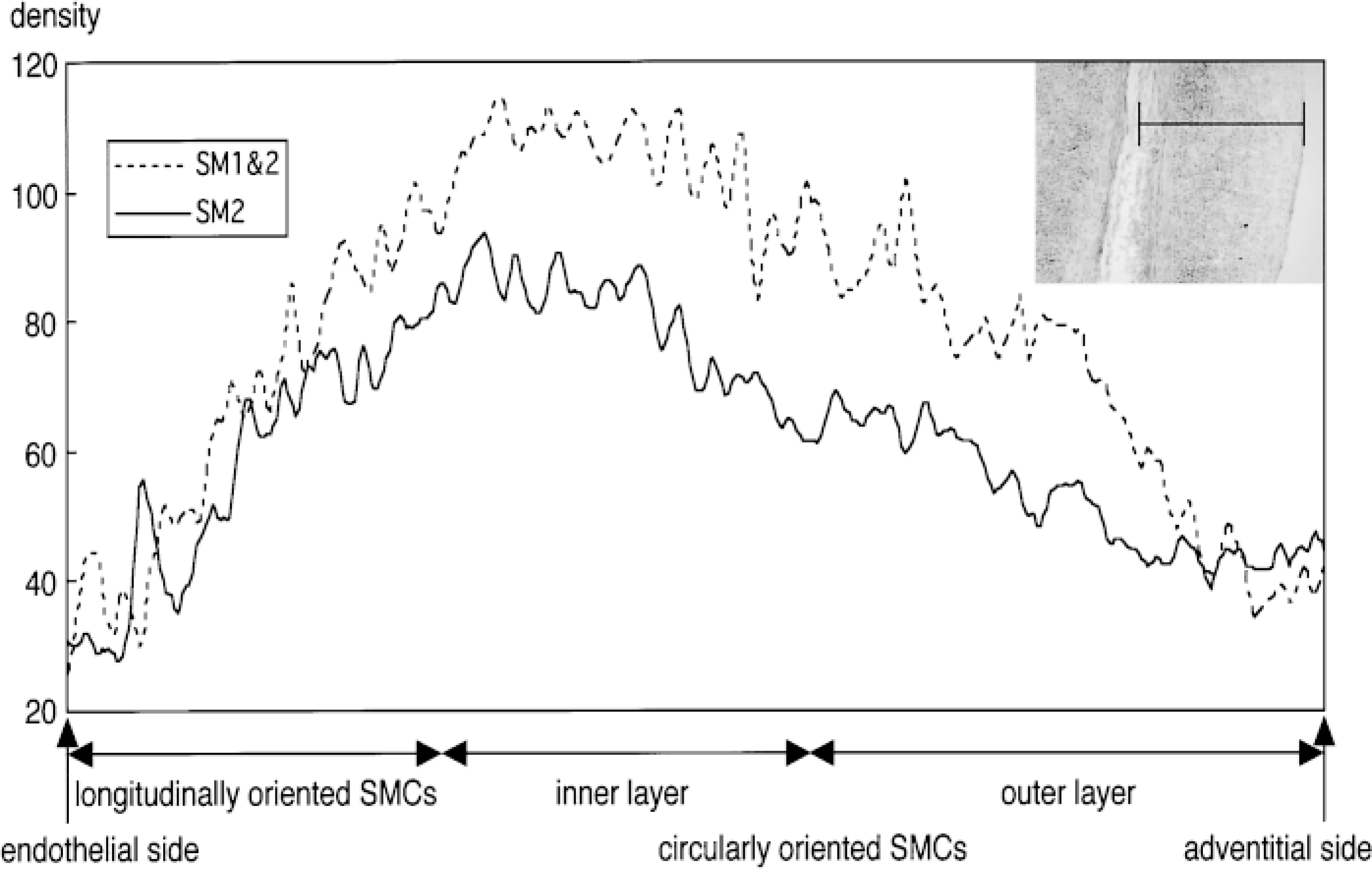
Densitometric data calculated by NIH Image (ver. 1.61) from the area shown in the tissue photograph. In the longitudinally oriented SMCs, only the SM2 MHC mRNA was detected. Strong SM2 MHC mRNA expression was detected in the longitudinally oriented and inner layer of the circularly oriented SMCs. SM1 MHC mRNA expression can be seen in the inner and outer layers of the circularly oriented SMCs when the density of the SM2 is subtracted from that of the SM1 and SM2.
Recent studies have shown that apoptosis, originally described by Kerr et al. (1972), contributes to arterial remodeling in the lamb during development (Cho et al. 1995) and to the regulation of cellularity in experimental intimal thickening in the rat (Bochaton-Piallat et al. 1995). Furthermore, Slomp et al. (1997) reported that apoptosis was found in the inner intima of the SMCs during development of the human DA. It is possible that this apoptosis is associated with ligament formation of the DA after birth.
To determine which part of the SMCs of the DA contribute to its closure after birth, we looked for areas in which SM2 MHC mRNA expression and apoptosis could be detected in the DA during development, using fetal and neonatal rabbits. Our results indicate that the longitudinally oriented SMCs and the inner layer of the circularly oriented SMCs play an important role in duct constriction and remodeling of the vessel wall after birth.
Materials and Methods
Sample Preparation
Japanese White rabbits on gestational Day 29 and at 1, 3, 5, 7, and 9 days after birth were used. Briefly, the DA was excised from the animal with or without the pulmonary artery (PA) and aortic artery (Ao). A few samples were embedded in OCT compound (Miles; Elkhart, IN), quickly frozen in normal hexane-liquid nitrogen, and stored at −80C for in situ hybridization to detect smooth muscle MHC mRNAs. For the terminal deoxynucleotidyl transferase-mediated dUTP-digoxigenin nick end-labeling (TUNEL) analysis, samples were fixed in 10% neutral buffered formalin at room temperature (RT) overnight. For electron microscopic analysis, samples were fixed in 3% glutaraldehyde buffered in 0.1 M sodium cacodylate (pH 7.3) at 4C for 1.5 hr and in 1% osmium oxide buffered in 0.1 M sodium cacodylate (pH 7.3) at 4C for 1.5 hr. For electrophoretic analysis of DNA ladder formation, samples were frozen in liquid nitrogen and stored at −80C until assayed.
In situ hybridization of 28S ribosomal RNA (positive control,
DNA Probes
The probe (antisense oligo-DNA with auxiliary ATT repeats at the 3' and 5' ends; antisense SM2 probe) was designed from the 39 nucleotides from rabbit SM2 smooth muscle cDNA which were inserted in SM1, and is therefore not homologous with SM1 (Sakurai et al. 1996). The probe was irradiated with ultraviolet light at a dose of about 10 kJ/m2 to form thymine-thymine (T-T) dimers (Koji and Nakane 1990). A 39-nucleotide sequence of the homologous region of the SM1 and SM2 was used as a positive control probe (antisense SM1 and SM2 probe). A 34-nucleotide sequence complementary to the part of the 28S ribosomal RNA (rRNA) was also used as a positive control probe (Koji and Nakane 1996). The SM2 sense oligo-DNA, SM1 and SM2 sense oligo-DNA, and nonirradiated complementary 28S rRNA probes were used as negative control probes.
In Situ Hybridization
The procedures, sensitivity, and reliability of in situ hybridization using the T-T dimerized oligo-DNA probe have been described previously in detail (Koji and Nakane 1990; Koji et al. 1994). Briefly, frozen longitudinal sections (5-6 μm) through the DA were placed on silane-coated glass slides and dried at RT. Tissue sections were fixed in 4% paraformaldehyde (PFA; Nacalai Tesque, Tokyo, Japan) in PBS, pH 7.4, at RT for 20 min. After rinsing in PBS (RT, 5 min, three times), the slides were treated successively with 0.2N HCl (RT, 20 min), 0.2% Triton X-100 (Sigma; St Louis, MO) in PBS (RT, 10 min), and 1 μg/ml proteinase K (Boehringer; Mannheim, Germany) in PBS (37C, 15 min). The sections were postfixed with 4% PFA/PBS and then immersed in 2 mg/ml glycine (Kanto Chemical; Tokyo, Japan) in PBS (RT, 15 min, two times) and in 40% deionized formamide (Boehringer) in 4 × SSC (SSC; 0.15 M NaCl and 0.015 M sodium citrate, pH 7.0) for 30 min at RT until hybridized. Hybridization was performed using 5 μg/ml T-T dimerized SM2 MHC cDNA dissolved in 10 mM Tris-HCl, pH 7.4, containing 0.6 M NaCl, 1.0 mM EDTA, 40% deionized formamide, 1 × Denhart's solution (Sigma), 250 μg/ml yeast tRNA (Sigma), 125 μg/ml salmon testis DNA (Sigma), and 10% dextran sulfate (Sigma) at 39C for 15-17 hr. After lengthy washing with 50% formamide/2 × SSC (37C, 1 hr, five times), 2 × SSC alone (RT, 15 min, two times), and PBS (RT, 5 min, two times), the sections were incubated for 1 hr at RT with PBS containing 5% BSA, 100 μg/ml salmon testis DNA, 100 μg/ml yeast tRNA, and 500 μg/ml of normal mouse IgG (Sigma). They were then reacted overnight at RT with HRP-mouse anti-(T-T dimer) antibody (1:60; Kyowa Medex, Tokyo, Japan) dissolved in PBS containing 5% BSA, 100 μg/ml salmon testis DNA, and 100 μg/ml yeast tRNA. The sections were then washed with 0.075% Brij 35 (Sigma) in PBS (RT, 15 min, four times), and a signal was detected (RT, 5 min) with 0.1 M sodium phosphate buffer (pH 7.5) containing 0.5 mg/ml DAB (Wako Pure Chemicals; Osaka, Japan), 0.025% CoCl2 (Kanto Chemical; Tokyo, Japan), 0.02% NiSO4(NH4)2SO4 (Wako), and 0.01% hydrogen peroxide (Wako).
Densitometry
Densitometric analysis was performed on a Macintosh computer using the public domain NIH Image (ver. 1.61) program (developed at the U.S. National Institutes of Health and available from the Internet by anonymous FTP from zippy.nimh.nih.gov, or on floppy disk from the National Technical Information Service, Springfield, Virginia, part number PB95-500195GEI).
In Situ Detection of DNA Strand Breaks
To identify nuclei with DNA strand breaks at the cellular level, TUNEL was performed using an ApopTag Plus kit (Oncor; Gaithersburg, MD). Briefly, paraffin-embedded 3-μm-thick longitudinal sections through the DA were cut onto silane-coated glass slides and dewaxed in a routine manner. After washing with PBS, the sections were treated with 50 μg/ml proteinase K in PBS at RT for 20 min. Endogenous peroxidase activity was quenched with 2% hydrogen peroxide in PBS at RT for 5 min. Then the ApopTag Plus kit was used. Counterstaining was performed with methyl green. Hematoxylin-eosin (H-E) and Masson trichrome staining was also performed for control and detection of connective tissue, respectively.
Transmission Electron Microscopy
Transmission electron microscopic analysis was performed to confirm the existence of nuclear shrinkage and chromatin condensation. Fixed samples were embedded in epoxy resin (Poly/Bed 812 embedding kit; PolySciences, Warrington, PA) and ultrathin sections were double stained with saturated uranyl acetate (Merck; Darmstadt, Germany) and 0.4% lead citrate (Nacalai Tesque) and examined in a JEM 1200EX (JEOL, Japan) electron microscope.
Electrophoretic Analysis of Extracted DNA
Frozen samples were minced and 5-10 μl of the samples were used. Forty microliters of lysis buffer [50 mM Tris-HCl (pH 7.8), 10 mM EDTA, 0.5% w/v sodium-N-lauroylsarcosinate (Nacalai Tesque)] was added and mixed well. Then 4 μl of proteinase K (10 mg/ml) was added and mixed well, and the samples were incubated at 50C for 90 min. Next, 2 μl of RNase A (10 mg/ml; Sigma) was added and the samples were incubated at 50C for 30 min. These samples were then run on 2% agarose gel (1 g of agarose/50 ml, 40 mM Trizma base, 5 mM glacial acetic acid, 2 mM EDTA) and stained with ethidium bromide.
Results
In Situ Hybridization of SM2 MHC mRNA
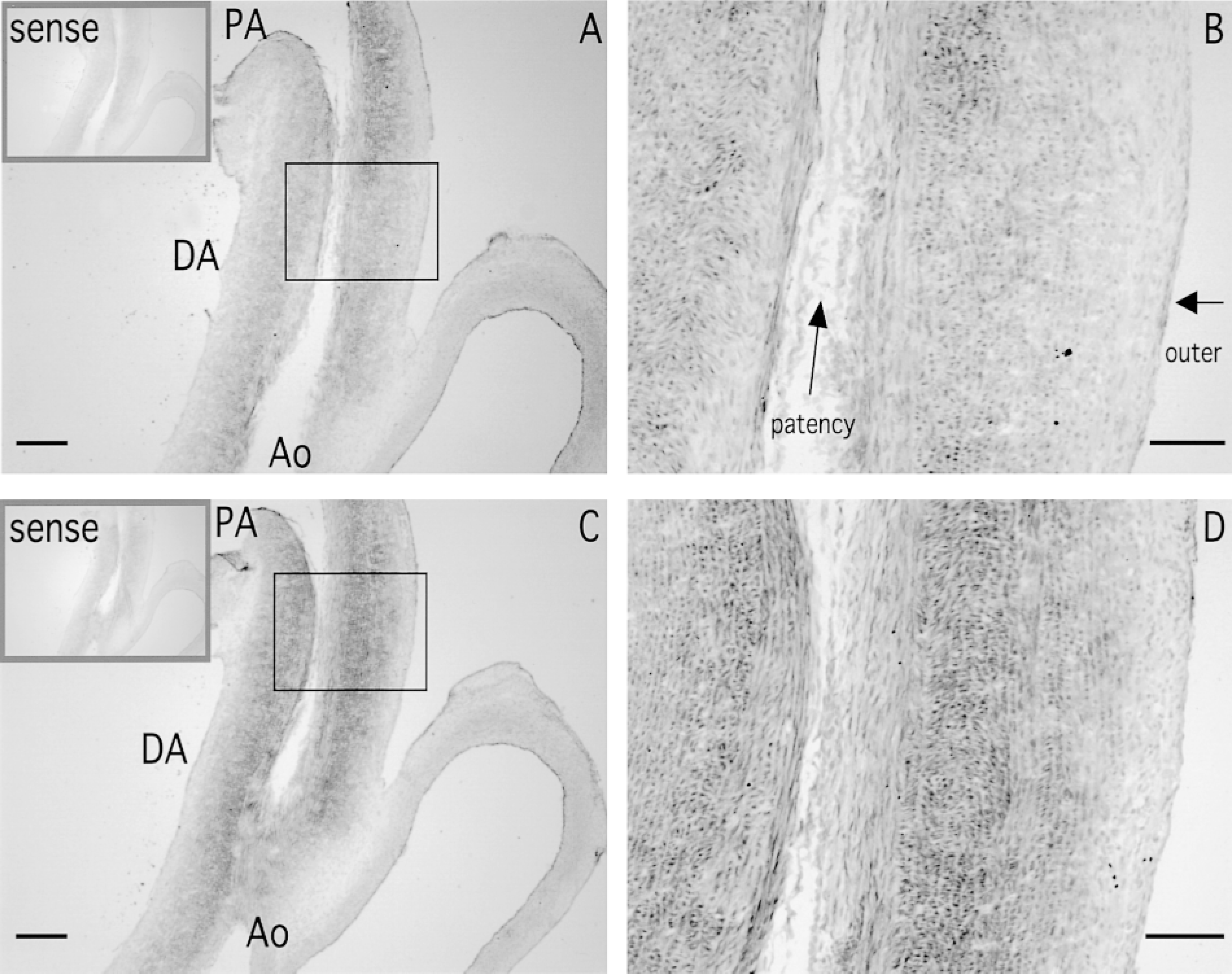
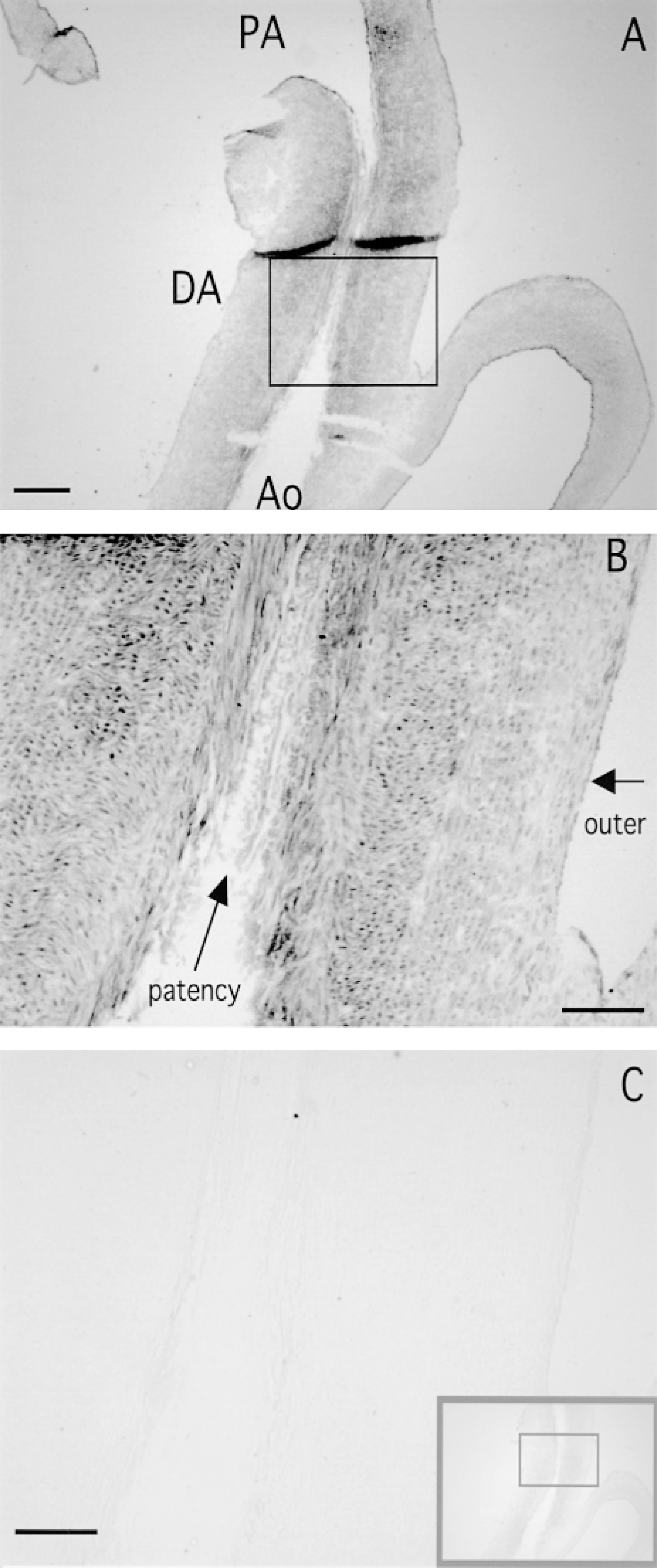
On gestational Day 29 (just before birth, when the DA is still patent), strong SM2 MHC mRNA expression was detected in the longitudinally oriented SMCs and in the inner layer of the circularly oriented SMCs of the DA (Figures 1A, 1B, and 2). However, in the outer layer of the circularly oriented SMCs of the DA, and in the PA and Ao, SM2 MHC mRNA expression was weak (Figures 1A and 1B). It has been shown that the SM2 MHC protein also appears strongly in the same area of the DA (Kim et al. 1993), which supports our data. The SM1 + SM2 MHC mRNA expression appeared more strongly in almost the same area as the SM2 only (Figures 1C and 1D). However, densitometric data showed that the longitudinally oriented SMCs expressed only the SM2 MHC mRNA, whereas SM1 MHC mRNA was expressed at the inner and outer layers of the circularly oriented SMCs (Figure 2).
A longitudinal section of a ductus arteriosus (DA) with a pulmonary artery (PA) and an aortic artery (Ao) taken on gestational Day 29 (
To verify the specificity of signals, we used an irradiated complementary 28S rRNA probe as a positive control, and sense probes and a nonirradiated complementary 28S rRNA probe as negative controls. Strong positive signals were detected with the irradiated complementary 28S rRNA probe (Figures 3A and 3B), but no signals were detected with the sense probes (gray squares in Figures 1A and 1C) and the nonirradiated complementary 28S rRNA probe (Figure 3C). Therefore, the signals of the SM2 MHC mRNA with the SM2 antisense probe confirmed its specificity.
Apoptotic Features by TUNEL
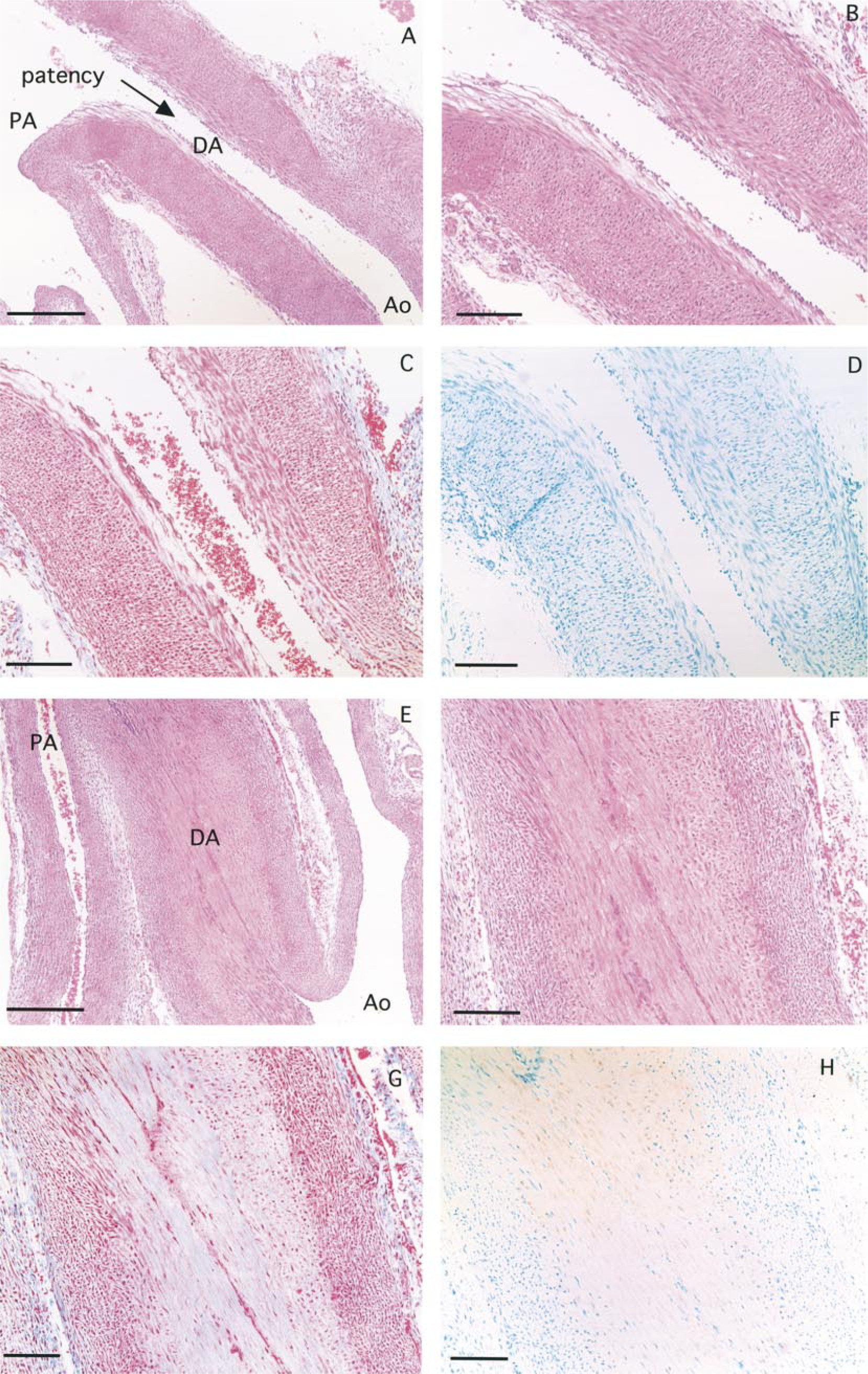
On gestational Day 29, the TUNEL-positive nuclei could not be seen in the cells of the DA, PA, and Ao (Figure 4D). However, TUNEL analysis revealed stained nuclei in the SMCs of the DA (Figure 4H) which was completely closed (Figure 4E) at 1 day after birth. In the PA and Ao, TUNEL-positive nuclei were not detected. Interestingly, a significant number (>90%) of TUNEL-positive nuclei were detected in the longitudinally oriented SMCs and the inner layer of the circularly oriented SMCs of the DA (Figure 4H). Very few nuclei of the circularly oriented SMCs in the media were stained by TUNEL. Furthermore, the Masson-stained section showed that the TUNEL-positive area in the DA was replaced by connective tissue from 1 day after birth (Figure 4G). A significant number (almost 100%) of TUNEL-positive nuclei were also observed in the longitudinally oriented SMCs and the inner layer of the circularly oriented SMCs of the DA, and this area was also replaced by connective tissue at 3 days after birth (Figures 5C and 5D). At 9 days after birth, the DA had become thin (Figure 5E). The number of TUNEL-positive nuclei had decreased and the amount of connective tissue had increased. However, a few SMCs in the media (outer layer) were still alive (Figures 5F–5H).
Apoptotic Features by Transmission Electron Microscopy
The transmission electron micrograph of the SMCs of the DA obtained on gestational Day 29 is shown as a control (Figure 6A). At 1 and 3 days after birth, the SMCs of the DA had become smaller and nuclear chromatin condensation and nuclear shrinkage could be seen, but the integrity of the plasma membrane was retained (Figures 6B and 6C). These observations are characteristic features of the apoptotic process (Arends et al. 1990). Figures 6D–6F show transmission electron micrographs of the SMCs of the DA at 5, 7, and 9 days after birth, respectively. Apoptotic features are clearly demonstrated. However, apoptotic cells were not observed in the SMCs of the PA and Ao (not shown).
Apoptotic Features by Electrophoretic Analysis
As shown in Figure 7, a faint but visible ladder, which is an early characteristic feature of apoptosis (Arends et al. 1990), was observable in the DA from 1 day after birth. In contrast, no ladder formation could be detected in DNA obtained from the PA and Ao.
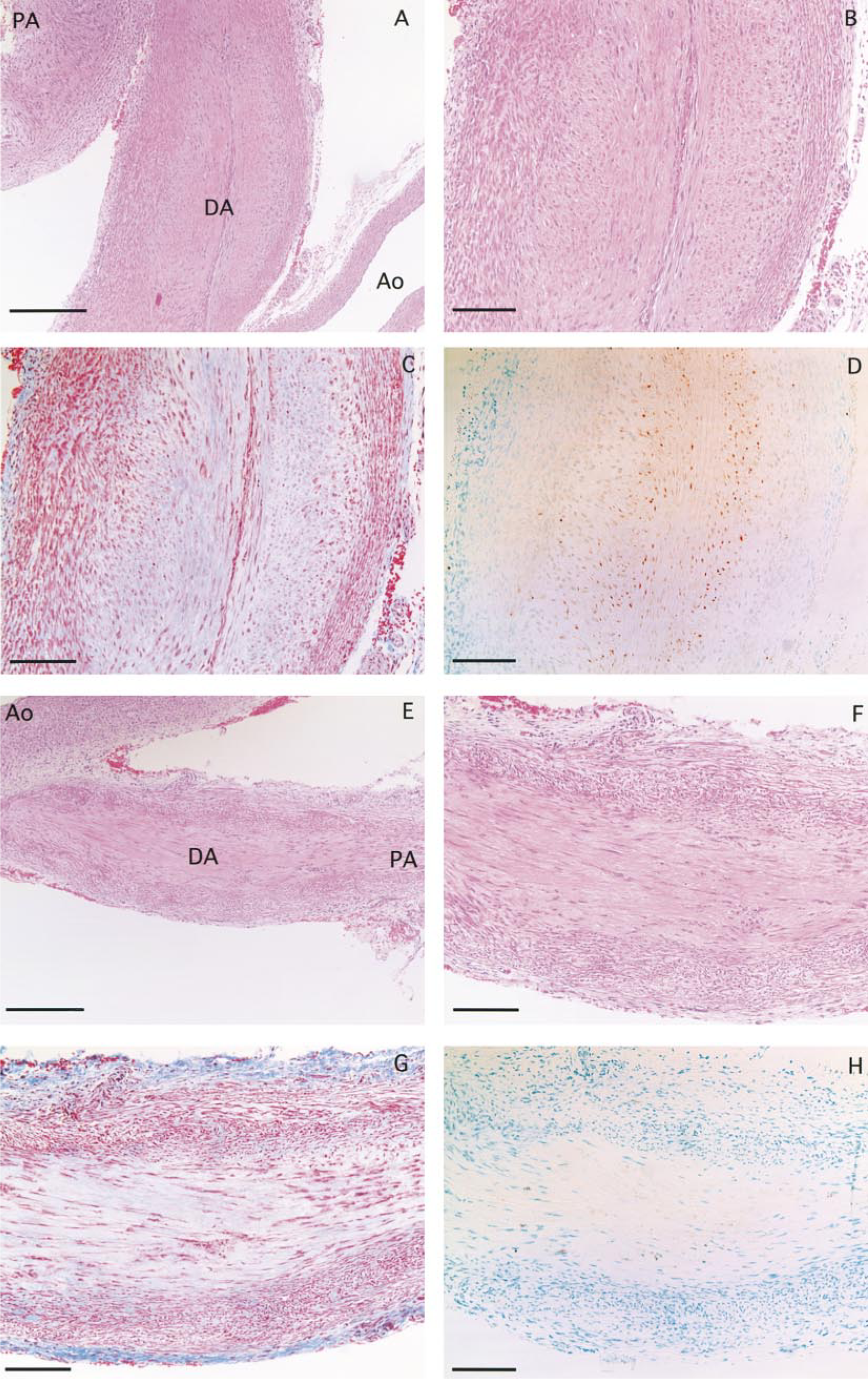
A longitudinal section of a ductus arteriosus (DA) with a pulmonary artery (PA) and an aortic artery (Ao) taken at 3 days (
Discussion
In this study we confirmed the localization of SM2 MHC mRNA expression in the DA just before birth. It is especially important to note that strong SM2 MHC mRNA expression was detected in the longitudinally oriented SMCs and in the inner layer of the circularly oriented SMCs (Figures 1A, 1B, and 2). We have already confirmed that the ratio of SM2/(SM1 + SM2) MHC mRNA expression is higher in the DA than in the PA and Ao at the fetal stage, and this ratio conversely becomes low compared with the PA and Ao at the neonatal stage (Matsuoka et al. 1995). In the present study we found no direct evidence showing that the SM2 phenotype provides contractile function for the SMCs. However, the SM2 is generally believed to be associated with the contractile phenotype of the SMCs, because SM2 MHC mRNA expression is abundant in the SMCs of esophagus, stomach, small intestine, and urinary bladder (Sakurai et al. 1996). These smooth muscle organs are often stretched and exhibit vigorous contractile activity. In addition, we have confirmed that cultured rabbit aortic SMCs undergoing cyclic mechanical stretching show an increase in SM2 MHC mRNA expression, but no changes were observed in the SM1 and non-muscle MHC gene expression in the same study (Imamura et al. 1992). Colbert et al. (1996) reported that retinoic acid may play a role in inducing, maintaining, and promoting the SM2 isoform in the developing DA, indicating the important role of the SM2 at duct closure. On the basis of the results presented here and of previous findings, the longitudinally oriented SMCs and the inner layer of the circularly oriented SMCs of the DA may directly play an important role in the closure of the DA after birth. Although their function is still unclear, the SMCs of the outer layer of the circularly oriented area are also believed to be associated with duct closure.
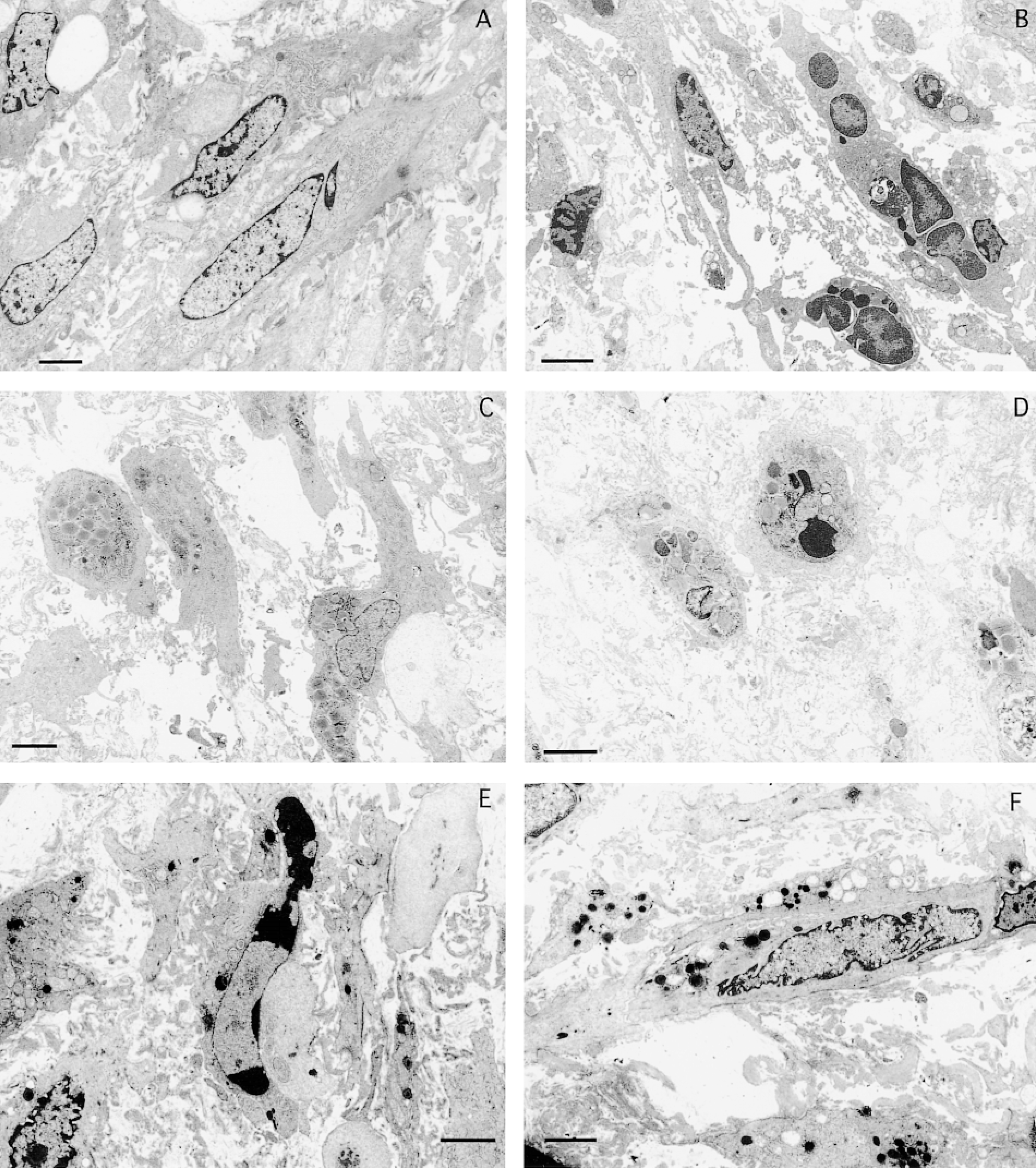
Transmission electron micrographs of the smooth muscle cells of the DA on gestational Day 29 (
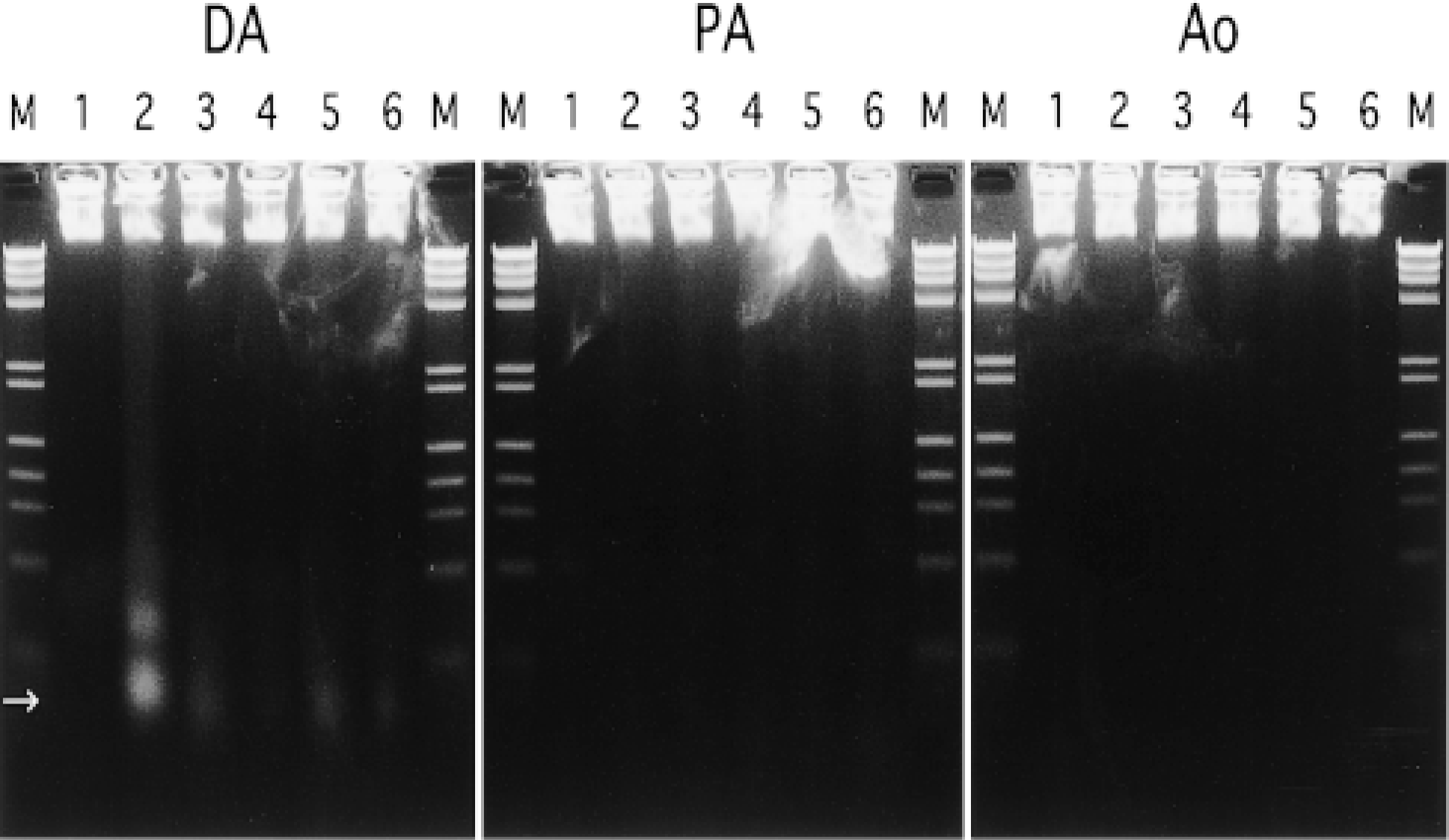
Agarose gel electrophoresis of DNA extracted from the ductus arteriosus (DA), pulmonary artery (PA), and aortic artery (Ao). Lane 1, gestational Day 29; Lane 2, Day 1; Lane 3, Day 3; Lane 4, Day 5; Lane 5, Day 7; Lane 6, Day 9; M, molecular markers; →, mononucleosome. In the DA, a faint but visible ladder is observable from 1 day after birth. However, no ladder formation can be detected in the PA and Ao.
At 1 day after birth, a significant number (>90%) of TUNEL-positive nuclei were detected in the longitudinally oriented SMCs and the inner layer of the circularly oriented SMCs of the DA (Figure 4H). Furthermore, the Masson-stained section revealed that the TUNEL-positive area in the DA was quickly replaced by connective tissue (Figure 4G). At 3 days after birth, the number of TUNEL-positive nuclei observed in the inner layer of the circularly oriented SMCs increased compared with 1 day after birth (Figure 5D). These findings indicate that the remodeling of the DA after birth is rapidly induced from the media of the constricted DA, especially from the longitudinally oriented and the inner layer of the circularly oriented SMCs. Slomp et al. (1997) reported that in the human neonatal DA (between 1 and 3 weeks of age), apoptotic cells were found in the areas of cytolytic necrosis in the inner media. However, in our study apoptotic cells were clearly detected in all areas, from the SMCs under the endothelial cells to the inner layer of the circularly oriented SMCs. It may therefore have taken a shorter time to replace smooth muscle with connective tissue in rabbit than in human.
Interestingly, increases in SM2 MHC mRNA expression and induction of apoptosis were observed at the same site in the media of the DA. This strongly suggests that the SMCs located in the longitudinally oriented and the inner layer of the circularly oriented area play an important role in duct constriction and remodeling of the vessel wall. Several investigators (Slomp et al. 1997; Mason et al. 1999) suggest that neointimal cushion formation is essential for closure of the DA after birth. The neointimal cushion is believed to be important as a prerequisite for complete anatomic closure of the DA after its constriction at birth (Silver et al. 1981; Gittenberger-de Groot et al. 1985). In addition, duct constriction itself may require not only the longitudinally oriented SMCs but also the inner layer of the circularly oriented SMCs because SM2 MHC mRNA expression was abundant in these SMCs. Furthermore, it is interesting that a single SMC may be led to contract at first, then to die by the apoptotic process, and finally to be replaced by connective tissue. Surprisingly, these phenomena occur one after another within a day after birth and may be associated with increased blood oxygen tension induced by the commencement of breathing after birth. Some investigators (Fay 1971; Kovalcik 1962; Roulet and Coburn 1981; Nakanishi et al. 1993) have demonstrated that hyperoxygen tension induces contraction of the DA. In addition, we reported that apoptotic cell death was induced by hyperoxygen tension in cultured SMCs of the DA but was not induced in the SMCs of the PA and Ao (Imamura et al. 1998). Therefore, both contraction and apoptosis in the SMCs of the DA are induced by hyperoxygen tension. However, ischemia after duct closure may also lead to apoptosis in the SMCs of the DA. More detailed analyses of the intracellular mechanisms that lead to smooth muscle contraction and apoptotic cell death in the DA will be important to elucidate the process of duct closure and the pathogenesis of the patent DA.
Footnotes
Acknowledgements
Supported by a Grant-in-Aid for Encouragement of Young Scientists (1995, 1996) from the Ministry of Education, Science and Culture, by an open research grant (1997, 1998) from the Japan Research Promotion Society for Cardiovascular Diseases, and by the Japan Shipbuilding Industry Foundation (1997, 1998).
We thank Ms Barbara Levene for reading the manuscript. We greatly appreciate the technical advice of Dr Takehiko Koji at Nagasaki University School of Medicine in establishing in situ hybridization using a nonradioactive T-T dimerized oligo-DNA probe. We also thank Yoshiyuki Furutani, BSc, at Tokyo Women's Medical University for technical help with densitometric analysis.