
Abstract
The goal of the present studies was to optimize mRNA detection with radioisotopic in situ hybridization histochemistry (ISHH). Test experiments performed on sections of rat brain tissue used computer-assisted image analysis to compare autoradiographic signals resulting when varying concentrations of 35S-labeled cRNA probes, dextran sulfate (DS), and dithiothreitol (DTT) were used for ISHH. We found that greatly enhanced corrected signal density (total density of signal area minus background density) was obtained using concentrations of probe and/or DS that were several-fold higher than those widely recommended in published ISHH procedures (probe concentration >4 × 104 cpm/μl; DS concentration >10%). Extended hybridization reaction (>16 hr) also significantly augmented the corrected signal density. Finally, nonspecific probe binding was greatly reduced and corrected signal density enhanced by including 750–1000 mM, rather than the widely used 10–200 mM DTT, in the hybridization buffer. These observations indicate that the low efficiency of hybridization and the formation of high background may largely compromise the sensitivity of routine ISHH procedures. We suggest that the new method using increased concentrations of 35S-labeled cRNA probe, DS, and DTT will be especially important for the cellular localization of rare mRNA species.
Keywords
D
The detection of rare mRNA species in individual neurons of the brain requires ISHH techniques that provide high sensitivity and also allow single-cell resolution. Recently, we observed that increasing concentrations of 35S-labeled cRNA hybridization probes, dextran sulfate (DS), and dithiothreitol (DTT) in the hybridization solution could noticeably enhance the autoradiographic hybridization signal for mRNAs encoding estrogen receptor isoforms. Application of this reformulated, but not the standard, hybridization solution enabled us to detect low expression levels of estrogen receptor-β (ERβ) mRNA in the majority of luteinizing hormone-releasing hormone neurons (Hrabovszky et al. 2000). It is critical to note that several control approaches, including recent immunocytochemical studies (Hrabovszky et al. 2001), confirmed the specificity of ERβ hybridization signal generated by the modified ISHH method. The unexpected dependence of hybridization signal on probe concentration conflicted with the common assumption that the conditions of the ISH reaction should satisfy the criterion to “saturate” target mRNA molecules in the tissue specimen (Nunez et al. 1989; Davenport 1998). Our findings raised the possibility that suboptimal hybridization conditions represent a general, albeit commonly overlooked, source of sensitivity loss in hybridization experiments.
The present studies were conducted to formally establish whether routine hybridization procedures provide submaximal hybridization signals. In addition, strategies were developed to improve the detection sensitivity of the ISHH method. To accomplish these aims, we assessed the impact of increased radioisotopic probe (>40,000 cpm/μl), DS (>10%), and DTT (>200 mM) concentrations in the hybridization solution, and also evaluated the influence of an extended hybridization time (>16 hr) on levels of autoradiographic hybridization signal and background.
Materials and Methods
Tissue Preparation
Adult female Sprague-Dawley rats (n = 3; 225 g bw) were maintained and treated in accordance with the NIH Guide for the Care and Use of Laboratory Animals and the University of Massachusetts IAUCC. After sacrifice of animals with CO2, brains were removed and snap-frozen on pulverized dry ice. Twelve-μm coronal sections through the cerebral cortex were prepared on a cryostat, thaw-mounted onto double gelatin-coated microscopic slides, dried, and stored in slide boxes at −80C as described previously (Petersen and McCrone 1994).
Preparation of cDNA Templates for In Vitro RNA Transcription
Both sense and antisense RNA probe sequences were used for hybridization assays. To ensure that results obtained were not probe-specific, we used several cRNA probes, each of which produced hybridization signal in the cerebral cortex. The cDNA template for androgen receptor (AR; Lubahn et al. 1988) was a 1016-bp fragment corresponding to bases 2821–3838 (generously provided by Dr. R. Handa). To prepare probes to the γ2 subunit of GABA-A receptor mRNA, a 304-bp cDNA template (gift from Dr. C.D. Carpenter), corresponding to nucleotides 1530–1833 (Shivers et al. 1989), was used. The 678-bp galanin cDNA was kindly provided by Dr. M.E. Vrontakis (Vrontakis et al. 1987). Finally, the 548-bp BamH1-Sal1 fragment of the rat progesterone receptor (PR) cDNA [rPR-1; generously provided by Dr. O.K. Park-Sarge (Park-Sarge and Mayo 1994)], was used as template for in vitro RNA transcription.
In Vitro Transcription of Antisense and Sense RNA Probes
The 10-μl transcription reactions were composed of the following ingredients: [35S]-UTP (NEN Life Science Products; Boston, MA), 120 pmol; linearized cDNA template, 1 μg; 5 × transcription buffer, 2 μl; 100 mM DTT (Sigma Chemical; St Louis, MO), 1 μl; 10 mM ATP, CTP, and GTP, 0.5 μl of each; 20 U/μl RNasin (Promega; Madison, WI), 0.5 μl; appropriate RNA polymerase (T3, T7, or SP6; Promega), 10 U. The reaction was allowed to proceed for 30 min at 37C. Then a second aliquot of RNA polymerase (10 U) was added and the mixture incubated for a second period of 30 min. After incubation, the volume was brought up to 90 μl with nuclease-free water, and we added 5 μl 1 M Tris-HCl buffer (pH 8.0), 1 μl tRNA, (25 mg/ml; Sigma), 1 μl 1 M MgCl2, 0.5 μl 20 U/μl RNasin (Promega), and 0.5 μl 10 U/μl DNase I (Roche Diagnostics; Indianapolis, IN). The template DNA was digested for 30 min at 37C, and finally the probe was purified by extraction with phenol/chloroform/isoamyl alcohol, then with chloroform/isoamyl alcohol. Unincorporated nucleotides were removed by two sequential NaCl/ethanol precipitations. The probe pellets were finally dissolved in 0.1% sodium dodecyl sulfate (SDS; Sigma) and radioactivity concentrations (in terms of cpm/μl) determined using a Beckman LS 6000 SC β-counter.
Preparation of Hybridization Buffer
The standard hybridization buffer was modified from a previously described formulation (Petersen and McCrone 1994) and contained the following: 50% ultra-pure formamide (Sigma), 2 × standard saline citrate (SSC) solution (1 × SSC = 0.15 M NaCl/0.015 M sodium citrate, pH 7.0), 10–30% DS (500,000 MW; Sigma), 1 × Denhardt's solution (0.02% each of bovine serum albumin, Ficoll, and polyvinylpyrrolidone; Sigma), 500 μg/ml yeast tRNA (Sigma), 500 μg/ml heparin sodium salt (Sigma), and 0.1% sodium pyrophosphate (Sigma). Immediately before use, the hybridization buffer was warmed to 56C and 200–1000 mM DTT (Sigma) was added and dissolved by vortexing. Changes in various components of this buffer are described for single test experiments.
Prehybridization Tissue Treatment Steps
Before ISHH, the slides were removed from the −80C freezer, warmed to RT (10 min), and loaded into RNase-free metal racks. The racks were placed in RNase-free plastic containers and sections were processed through the following prehybridization steps: 30-min fixation in phosphate-buffered (pH 7.4) 4% formaldehyde solution; 2-min rinse in 2 × SSC solution; 10 min acetylation in 0.25% acetic anhydride (Sigma)/0.9% NaCl/0.1 M triethanolamine (pH 8.0; Sigma) (Tecott et al. 1987); a brief rinse in 2 × SSC solution; dehydration in 70, 80, 95, and 100% ethanol (2 min each); delipidation in chloroform (5 min), then partial rehydration in 100% followed by 95% ethanol (2 min each). The slides were finally air-dried on trays.
In Situ Hybridization
The slides were placed in Nalgene boxes in which the air was humidified using caps of 50-ml centrifuge tubes containing distilled water. Twenty-five μl of hybridization solution was pipetted onto each section and covered with a glass coverslip. The standard hybridization reaction was carried out at 56C for 16 hr (overnight).
Posthybridization Tissue Treatment Steps
The coverslips were floated off the slides in 1 × SSC solution and the excess of hybridization solution was rinsed off in three changes of 1 × SSC solution. Then the slides were loaded into metal racks and the following incubation steps were carried out under agitation: 1 × SSC (RT), twice for 20 min; 50% formamide/2 × SSC mixture (52C), twice for 30 min; 2 × SSC (RT), 10 min; 50 μg/ml RNase A (Roche Diagnostics) dissolved in 500 mM NaCl/10 mM Tris-HCl/1 mM EDTA, pH 7.8, 37C), 30 min; rinse in 2 × SSC (RT), 10 min; 50% formamide/2 × SSC (52C), 30 min. Finally, the slides were rinsed in 2 × SSC buffer at RT (10 min), dipped briefly in distilled water (2 sec), rinsed in 70% ethanol (10 min), and air-dried on slide trays.
X-ray Autoradiography
For X-ray autoradiography, the slides were aligned in exposure cassettes and apposed to Kodak β-Max autoradiographic films for 2–14 days. The autoradiographs were developed with a Konica SRX-101A automated film processor.
Emulsion Autoradiography
After the development of film autoradiographs, the slides were dipped in Kodak NTB-3 nuclear emulsion (diluted at 1:1 with distilled water) and exposed for 1–4 weeks. The images were developed using Dektol developer (Kodak; Rochester, NY) for 2 min, rinsed with distilled water for 30 sec, and fixed with Kodak fixer for 5 min. Then the sections were rinsed in distilled water for 5 min and dried on slide trays.
Coverslipping of Sections
The sections were immersed in xylenes for 2 min, then coverslipped using DPX mounting medium (Fluka Chemie; Buchs, Switzerland) and used for light microscopic evaluation.
Computerized Semi-quantitative Image Analysis of X-ray Film Autoradiographs
X-ray films were placed on a light box and images were captured and digitized using a BioQuant Windows image analysis system (R and M Biometrics; Nashville, TN) interfaced with a CCD video camera (Hitachi Denshi; Tokyo, Japan) and an AF MicroNikkor 60-mm objective. The average gray level of highlighted pixels (on a 0–255 scale) was measured over a similar region of the cingulate cortex or the retrosplenial cortex in each section to determine the total density of the signal area (“total signal density”). In addition, “background density” measurement was taken over the white matter of the corpus callosum in each section. The background density measurement was subtracted from the total signal density to obtain the “corrected signal density” value. The mean of replicate measurements was determined for each test group, and means were compared by ANOVA. Newman-Keuls tests were performed post hoc when indicated.
Processing of Photographic Images
Representative film autoradiographs from each group were used for photographic illustrations. In addition, emulsion autoradiographs were examined using a Nikon light microscope and photographed with the CCD video camera. Digital images were processed using Adobe Photoshop 4.0 software (Tucson, AZ). In each experiment, representative photographs were merged into a single table before editing to maintain a reliable visual comparison of individual panels.
Preparation of RNase A-pretreated Sections for Control Studies
The slides were removed from the −80C freezer, placed in Coplin jars, and fixed for 10 min in acetone. The sections were air-dried briefly, then incubated in RNase A solution (50 μg/ml; Roche Diagnostics) for 30 min, as described above. To remove residual RNase A from the sections, the slides were rinsed abundantly with 2 × SSC solution (five times for 5 min). Formaldehyde fixation and further prehybridization treatment steps of the RNase-treated sections were performed in the same way as for untreated sections.
Results
Application of 1000 mM Instead of 200 mM DTT to Reduce Background Density of X-ray Film Autoradiographs
In practice, the use of high amounts of radioisotopic probe is limited by the formation of unacceptably high nonspecific background. In an attempt to overcome this problem, we addressed the advantage of increasing the concentration of DTT from 200 mM to 1000 mM in the hybridization solution. A sense-strand RNA transcript to the GABA A γ2 receptor subunit was used for the first hybridization experiment in the presence of either 200 mM or 1000 mM DTT. Because the pattern of background generated by this probe was not similar to the distribution of neuronal cells (Figures 1A-1D), it was unlikely that cross-hybridization of the probe to tissue nucleic acids contributed to the background we observed. Test experiments were performed using our standard hybridization solution containing 40,000 cpm/μl probe and 10% DS, as well as a modified hybridization solution that included a largely elevated probe concentration (200,000 cpm/μl) and 20% instead of 10% DTT, which further increased the effective concentration of the probe (Wahl et al. 1979). For research design, see overlays in Figures 1A–1D and table in Figure 2. Background levels over both the white matter (corpus callosum) and the gray matter (cingulate cortex) were analyzed and compared using three-way ANOVA, with main effects of DTT concentration (200 mM vs 1000 mM), probe DS contents (40,000 cpm/μl-10% vs. 200,000 cpm/μl-20% DS) of the hybridization solution, and tissue characteristics (white matter vs gray matter).
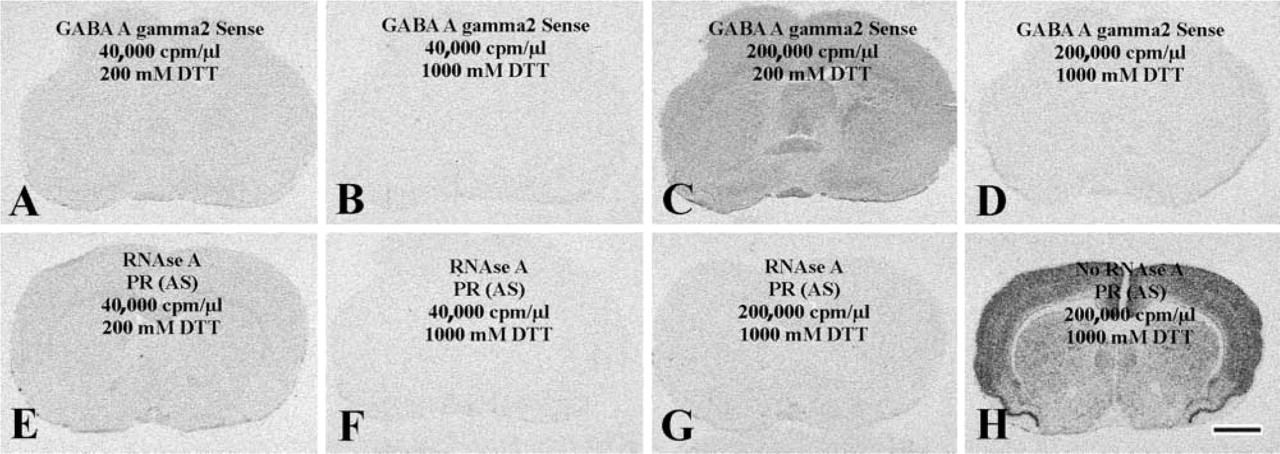
Reduced nonspecific probe binding after application of 1000 mM instead of 200 mM DTT in the hybridization solution. (
We found that the use of DTT at 1000 mM instead of 200 mM significantly reduced background density (F = 1463.18; p<0.05). Compare Figures 1B to 1A and Figures 1D to 1C and blocks 2B to 2A and 2D to 2C in Figure 2. The extent of reduction was high when increased amounts of probe (200,000 cpm/μl) and DS (20%) were used (32% in the white matter; compare blocks 2D to 2C in Figure 2), as opposed to the slight (but significant; p < 0.05) reduction observed using the standard probe (40,000 cpm/μl) and DS (10%) concentrations of our procedure (18% in the white matter; compare blocks 2B to 2A in Figure 2).
The inclusion of enhanced probe/DS concentrations in the hybridization solution significantly augmented background density (F = 549.64; p < 0.05), as shown by the comparison of Figure 1C to Figures 1A and 1D to Figure 1B and blocks in Figure 2C to 2A and Figure 2D to 2B. Background density enhancements were robust if the hybridization buffer contained 200 mM DTT (30%; compare blocks 2C to 2A in Figure 2) and slight if the hybridization solution contained 1000 mM DTT (7%; compare blocks 2D to 2B in Figure 2).
Tissue characteristics (white or gray matter) also exerted a statistically significant effect on background (F = 9.94; p < 0.05). Newman-Keuls test showed that film density over the white matter was significantly higher (p<0.01) than over the gray matter (Figure 1C; compare the two columns in block C of Figure 2) if high probe/DS were combined with low DTT. In contrast, background was evenly distributed between the white matter and the gray matter when we used the low probe/DS combination with low DTT (p = 0.18; Figure 1A; compare the two columns in block A of Figure 2), low probe/DS with high DTT (p = 0.59; Figure 1B; two columns in block B of Figure 2) or high probe/DS in the presence of high DTT (p = 0.79; Figure 1D; two columns in block D of Figure 2).
A second experiment used RNase A-pretreated sections for hybridization with an antisense PR probe. This approach was designed to prevent any hybridization to occur between the probe and tissue RNA molecules (Tecott et al. 1987; Höfler et al. 1998; Sunday 1998). The effects of DTT concentration (200 mM vs 1000 mM), probe-DS concentrations (40,000 cpm/μl probe-10% DS vs 200,000 cpm/μl probe-20% DS) and tissue characteristics (white vs gray matter) on background density were significant (p<0.05; analysis not detailed) and entirely reminiscent to the observations made using the sense-strand GABA A γ2 receptor subunit probe (compare Figures 1E to 1A, 1F to 1B, and 1G to 1D in representative photomicrographs obtained using identical probe, DS, and DTT concentrations). In addition, the modified hybridization procedure simultaneously using high DTT (1000 mM), high probe (200,000 cpm/μl), and high DS (20%) concentrations in RNase A-untreated control sections generated autoradiographic images characterized by high signal and low background labeling (compare signal density in gray matter vs background density in white matter structures in Figure 1H).
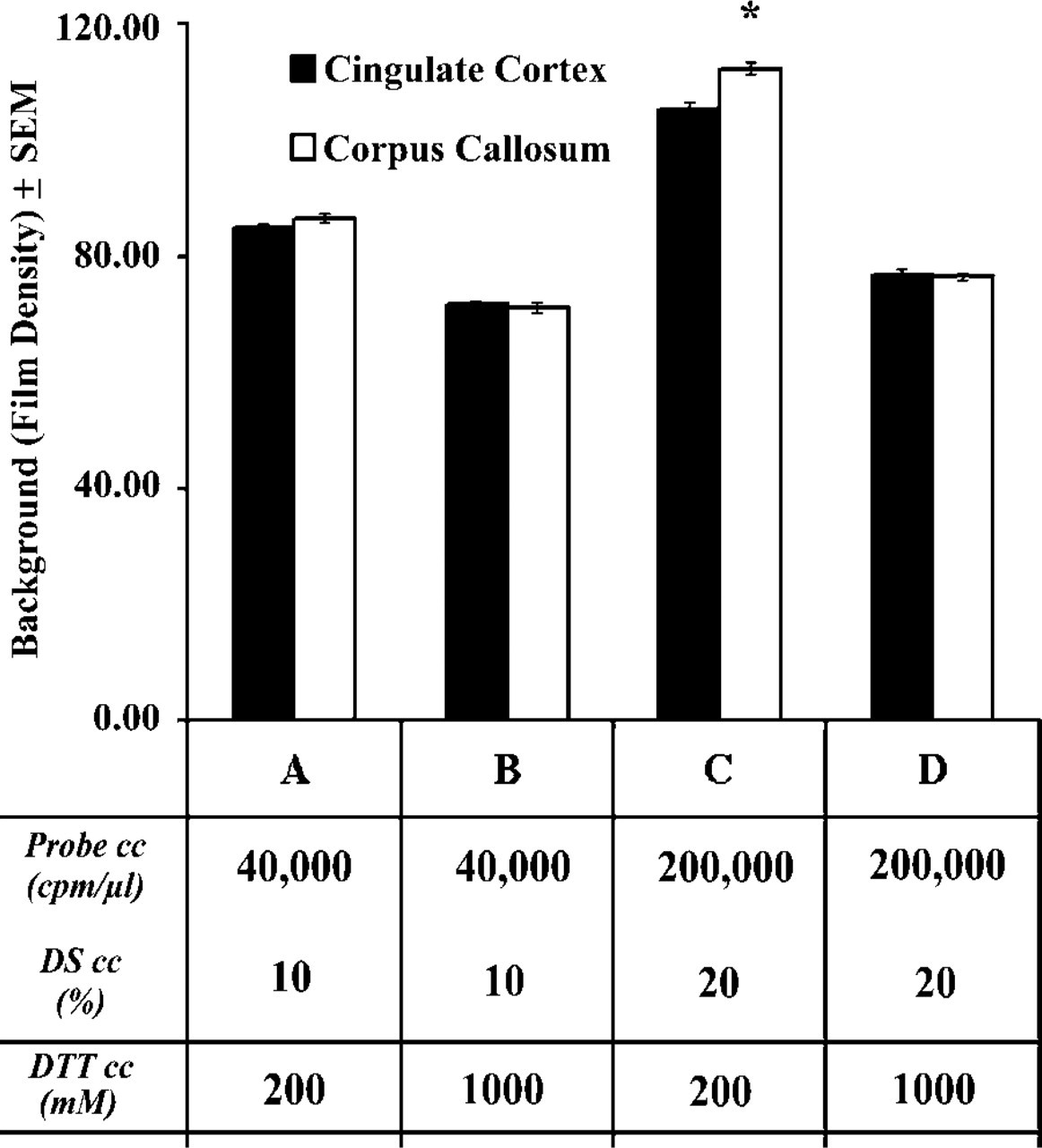
Quantitative analysis of autoradiographic background as an effect of elevated DTT concentration, increased (sense) probe-DS concentrations, and tissue characteristics. The application of 1000 mM (
Effects of Increased Probe, Increased DS, or Both, on Hybridization Signals
To address the potential advantage of using increased probe and increased DS concentrations in the hybridization solution, consecutive sections were divided into nine groups (six to eight sections/group) and hybridized for 16 hr with the antisense probe to the γ2 subunit of the GABA-A receptor, using three different concentrations of probe (40,000, 80,000, or 120,000 cpm/μl) and DS (10, 20, or 30%) in the hybridization solution. High amounts of DTT (750 mM) were added to the hybridization buffer to suppress nonspecific probe binding. After posthybridization treatments, the slides were exposed to X-ray films for 50 hr, then the autoradiographs were developed (Figure 3). Corrected signal density was determined in each treatment group (Figure 5) and two-way ANOVA, with DS concentration and probe concentration as main effects, was performed. Autoradiographs derived from 14C standards (using empty film density for background correction) were also analyzed to determine the 14C radioisotope concentrations causing identical corrected density to individual groups of hybridized sections. Finally, the full set of sections used in this experiment was coated with nuclear track emulsion. The emulsion autoradiographs were developed after 7 days of exposure and were used for light microscopic evaluation of hybridization signals in the cingulate cortex (Figure 4).
The distribution pattern of hybridization signal for the GABA A γ2 mRNA was identical using any combination of DS and probe concentrations, but film density differed largely in each group (Figure 3). Corrected signal density of the cingulate cortex showed robust enhancements as probe and/or DS contents of the hybridization solution increased (Figure 5). The influences of probe (F = 63.59; p<0.01) and DS (F = 864.80; p<0.01) concentrations were statistically significant. Furthermore, they showed significant interaction (F1,2 = 3.67; p<0.01) (Figure 5). The corrected signal density produced by our routine hybridization procedure was increased approximately fourfold in this experiment by raising probe and DS concentrations (compare the last column to the first column in Figure 5 and Figures 3I to 3A). A similar enhancement of corrected density could be generated by a tenfold increase in 14C radioisotope concentration (radioactivity/area), as we established by the analysis of co-exposed 14C standards. In addition, emulsion autoradiographs of the cingulate cortex (Figures 4A-4I) demonstrated that individual neurons gained improved definition by the clustering of silver grains when probe and/or DS was used at increased concentrations. Diffuse background, represented by homogeneously scattered grains in the molecular layer of the cingulate cortex (middle portion of individual panels in Figure 4), was negligible in all groups.
Use of a Series of Different Test Probes to Extend the Validity of the Concept that Routine Hybridization Conditions Produce Submaximal Hybridization Signals
We used a series of test probes to various targets (galanin, PR, and AR mRNAs) and of different lengths to address the issue of whether increased concentrations of probe and/or DS can further increase hybridization signals (corrected signal density). Results of ISHH studies using the galanin (Figures 6A-6D), the PR (Figures 6E and 6F), and the AR (Figures 6G and 6H) probes under various test conditions uniformly indicated that increased signal intensities could be achieved as probe and/or DS concentrations were increased in the presence of 800 mM DTT. For each test probe, the effects of increased probe (in Figures 6B vs 6A, Figures 6F vs 6E) and increased DS (in Figures 6D vs 6B and Figures 6H vs 6G) concentration on corrected signal densities were statistically significant by one-way ANOVA (p<0.05). These findings corroborated the idea that the lower probe and DS concentrations did not allow target mRNA saturation to occur.
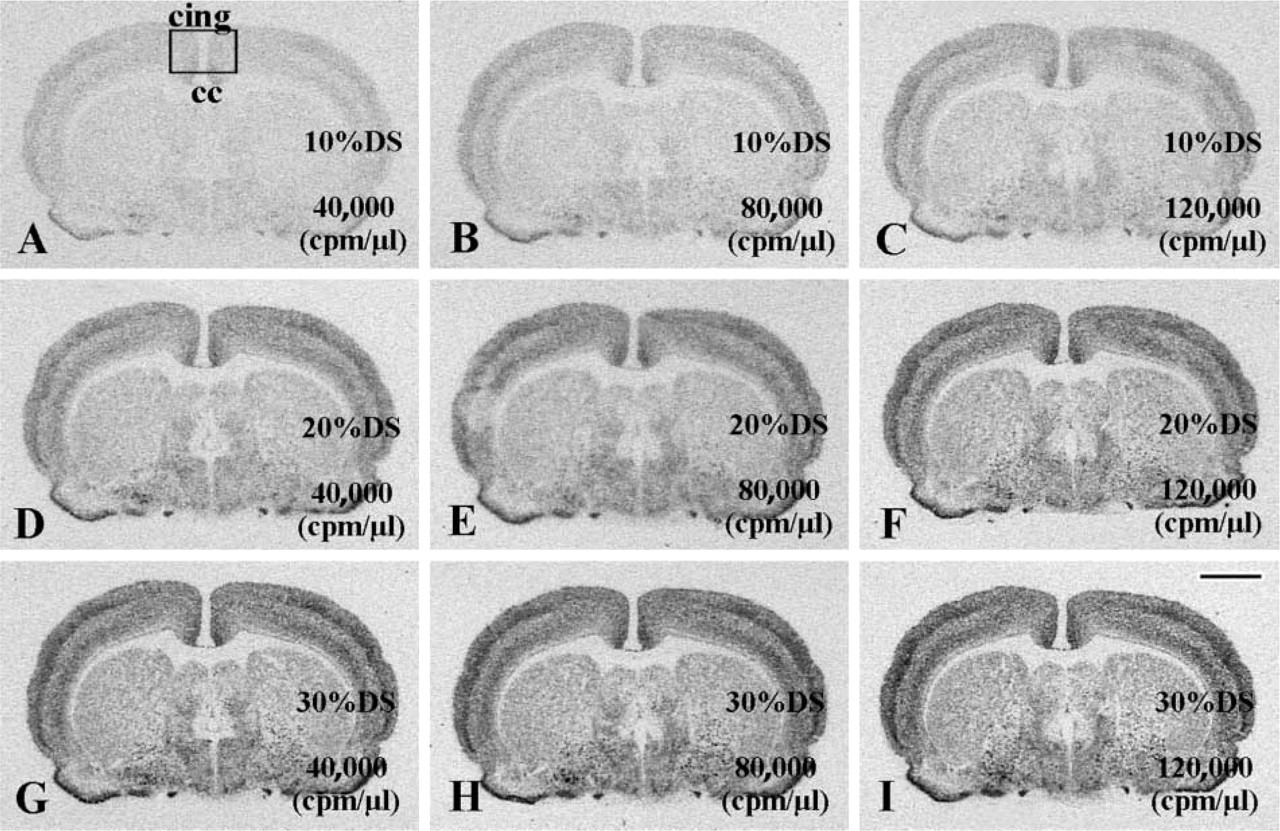
X-ray film autoradiographs illustrating enhanced hybridization signals after application of increased GABA-A γ2 probe and DS concentrations. The lowest signal produced by the standard formulation of hybridization solution (40,000 cpm/μl probe and 10% DS) in
Use of Extended Hybridization Time as an Alternative to Increased Probe to Further Verify the Concept that Regular Hybridization Reactions are Incomplete
On the basis of the concept that incomplete hybridization reactions should provide higher signals if allowed to proceed longer, the hybridization time was extended from the commonly used 16 hr (overnight hybridization; Angerer and Angerer 1981) to 40 hr. High DTT (800 mM) in the hybridization solution was used to suppress nonspecific probe binding. The first group of slides was removed from the incubator after 16 hr of hybridization with the antisense PR probe, then processed through posthybridization treatments. The second set of slides was hybridized for 40 hr and then processed through identical posthybridization steps. Using any probe-DS combinations, the extended hybridization reaction (40 hr vs 16 hr) augmented significantly (one-way ANOVA; p<0.05) the corrected signal density of the retrosplenial cortex (compare Figures 7E to 7A, 7F to 7B, 7G to 7C, and 7H to 7D). This observation was in harmony with the concept that hybridization reactions did not come to completion after 16 hr.
Titration of The Probe Concentration that Generates Maximal Corrected Signal Density
A broad range of probe concentrations were tested to establish hybridization conditions that provide maximal corrected signal density in a 16-hr hybridization reaction. To minimize background associated with high amounts of radioisotope in the hybridization solutions, the test experiment used a PR probe that was labeled to low specific activity in the presence of 108 μM UTP and 12 μM [35S]-UTP. Using this approach, the “plateau” phase of the hybridization reaction was expected at tenfold lower radioisotope concentration than using a probe labeled to maximal specific activity (in the absence of unlabeled UTP). Probe concentrations ranging from 4,000 cpm/μl to 176,000 cpm/μl were tested in a hybridization buffer that contained 20% DS and 1000 mM DTT. For each section group hybridized, mean total density, background density, and corrected signal density values were determined from film autoradiographs. For experimental design, see columns in Figure 8. Results of this study established that the highest value of corrected signal density (asterisk in Figure 8) was obtained using 20% DS and 40,000–64,000 cpm/μl probe. When compared with results of the standard procedure (10% DS and 4,000 cpm/μl of the low specific-activity test probe), this represented a threefold total increase of corrected signal density, of which 80% was already achieved by simultaneously raising DS from 10% to 20% and probe from 4000 to 16,000 cpm/μl. We also found that increasing the concentration of probe from 64,000 cpm/μl to 88,000 cpm/μl or further tended to slightly decrease the computable value of corrected signal density. This phenomenon was a consequence of a sudden rise in background density (Figure 8).

Emulsion autoradiographs showing enhanced hybridization signals after application of increased GABA-A γ2 probe and DS concentrations. (
Discussion
The results of these studies demonstrate that the sensitivity of conventional ISHH procedures can be improved significantly by increasing the concentrations of 35S-labeled cRNA probe (>40,000 cpm/μl), DS (>10%), and DTT (>200 mM) in the hybridization solution, or by using an extended hybridization reaction time (>16 hr). These modifications will facilitate the cellular localization of low-abundance mRNAs in brain and other tissues.
The Concept of Target mRNA Occupation (Saturation)
A plateau phase in hybrid formation was demonstrated in vitro using immobilized nucleic acid targets (Galau et al. 1977a, b). Although the kinetics of the ISH reaction are far more complex, various hybridization protocols used in the literature are based on the assumption that most targeted mRNA molecules in the tissues would be bound to probe by the end of the hybridization period (saturation phenomenon) (Nunez et al. 1989; Davenport 1998). In contrast to this concept, the results of the present studies demonstrate that the corrected signal density is still submaximal when 35S-labeled cRNA probe and DS are used at the highest concentrations we encountered in the literature. It is important to point out that the enhancement of corrected signal density by increased probe and/or increased DS required an improved protection against background formation, which we were able to achieve by the inclusion of 750–1000 mM instead of 10–200 mM DTT in the hybridization mixture. Without this modification we found that the use of increased probe and/or DS generated extremely high background over both signal and background regions, often reducing the corrected signal density value. Although specific probe binding (hybridization) to the signal area was probably increased in the presence of low DTT concentrations, the enhancement of background density appeared to exceed the enhancement of total cortical density (resulting from enhanced specific and nonspecific probe binding), with a net result of a reduced corrected signal density. Studies showing the non-linear optical density response of X-ray films to radiation (film grayness increases to a lesser extent than does radioactivity in the corresponding tissue area; Kuhar et al. 1985; Davenport 1998) provide a partial explanation of this phenomenon. It also follows that the “maximal corrected signal density” and the “highest signal-to-background ratio” in a saturation experiment using gradually increasing concentrations of probe for hybridization indicate mRNA target saturation only if background noise can be kept at a constant level. Because the simultaneous use of 1000 mM DTT and a low specific activity PR probe in our saturation experiment generated background levels that were unaffected by a broad range of probe concentrations, it is probable that the probe concentrations approaching the corrected signal density plateau, were close to “saturation” of most of the hybridizable mRNA targets. The formation of a corrected signal density plateau at stable background levels was a fundamental observation in this study, supporting the basic concept that a finite number of target molecules were present in the tissue for hybridization with the PR probe.
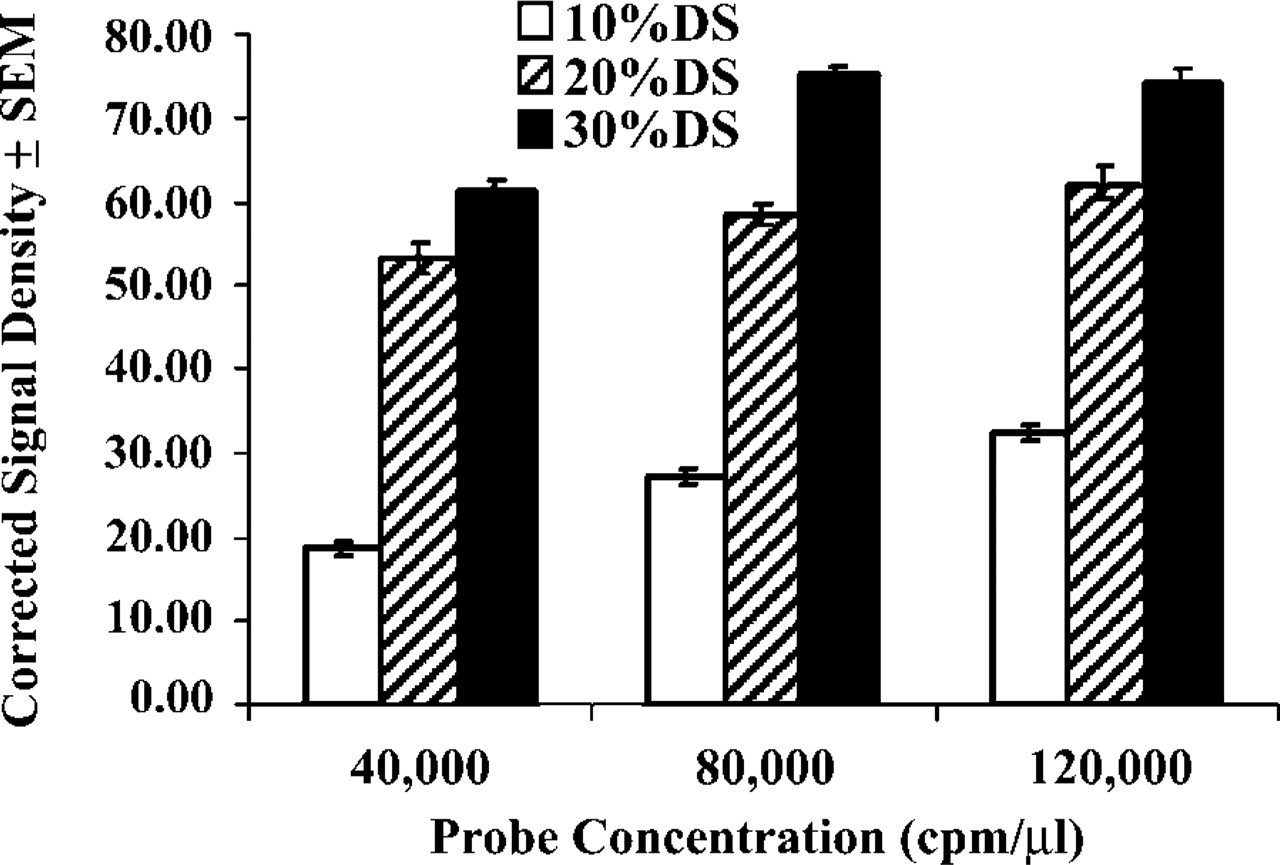
Effects of varying GABA-A γ2 probe (40,000, 80,000, or 120,000 cpm/μl) and DS (10, 20, or 30%) concentrations on corrected signal density of the cerebral cortex. Increased probe and increased DS concentrations significantly (p<0.01) enhanced the computable values of corrected signal density, and their effects were synergistic (p<0.01). Representative sections of each group were used for photographic illustrations in Figures 3 and 4. For detailed statistics, see text.
DTT Concentration
Although the mechanism by which DTT reduces nonspecific background is only partially understood (Zoeller et al. 1997), DTT at 10–200 mM represents a basic component of the hybridization solution. Our hybridization experiments using either sense-strand RNA transcripts or antisense probes on RNase A-pretreated sections established that the ability of DTT to reduce background can be largely enhanced by raising its concentration from 200 mM up to 1000 mM in the hybridization solution. Increased DTT was able to reduce background at any probe and DS concentrations we tested, but its use became critically important when elevated probe and/or DS concentrations were present in the hybridization solution. These data encourage the routine application of 750–1000 mM DTT in the hybridization solution as an efficient tool to prevent high background.
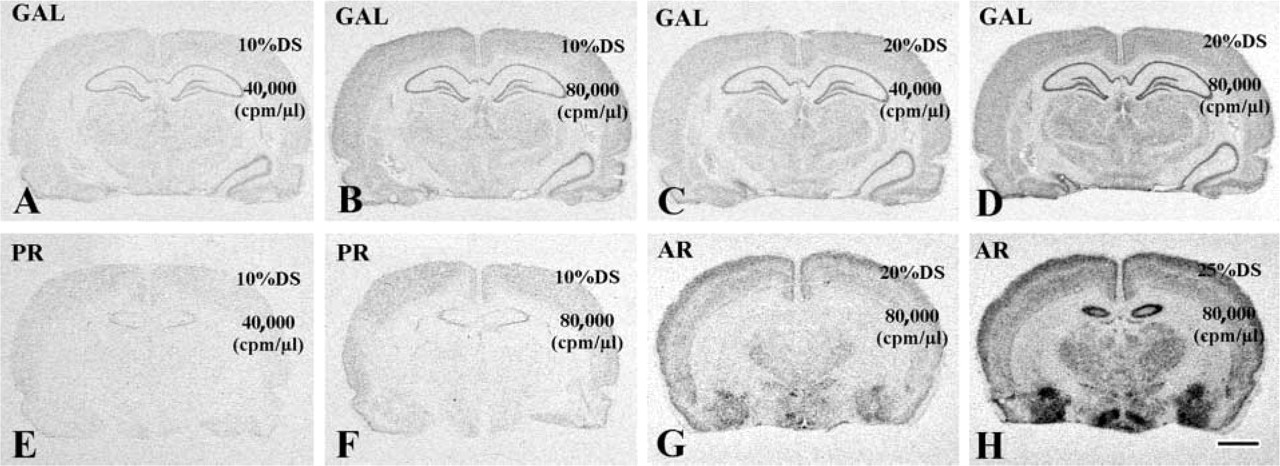
Enhanced ISHH signals after use of increased probe or DS concentrations. Use of test probes to galanin (
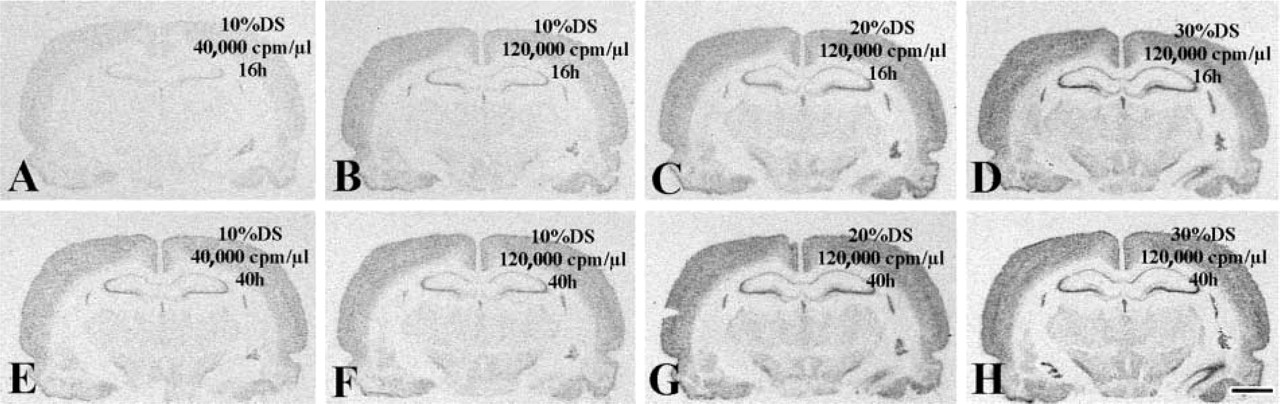
Use of extended hybridization reactions to enhance the ISHH signals. (
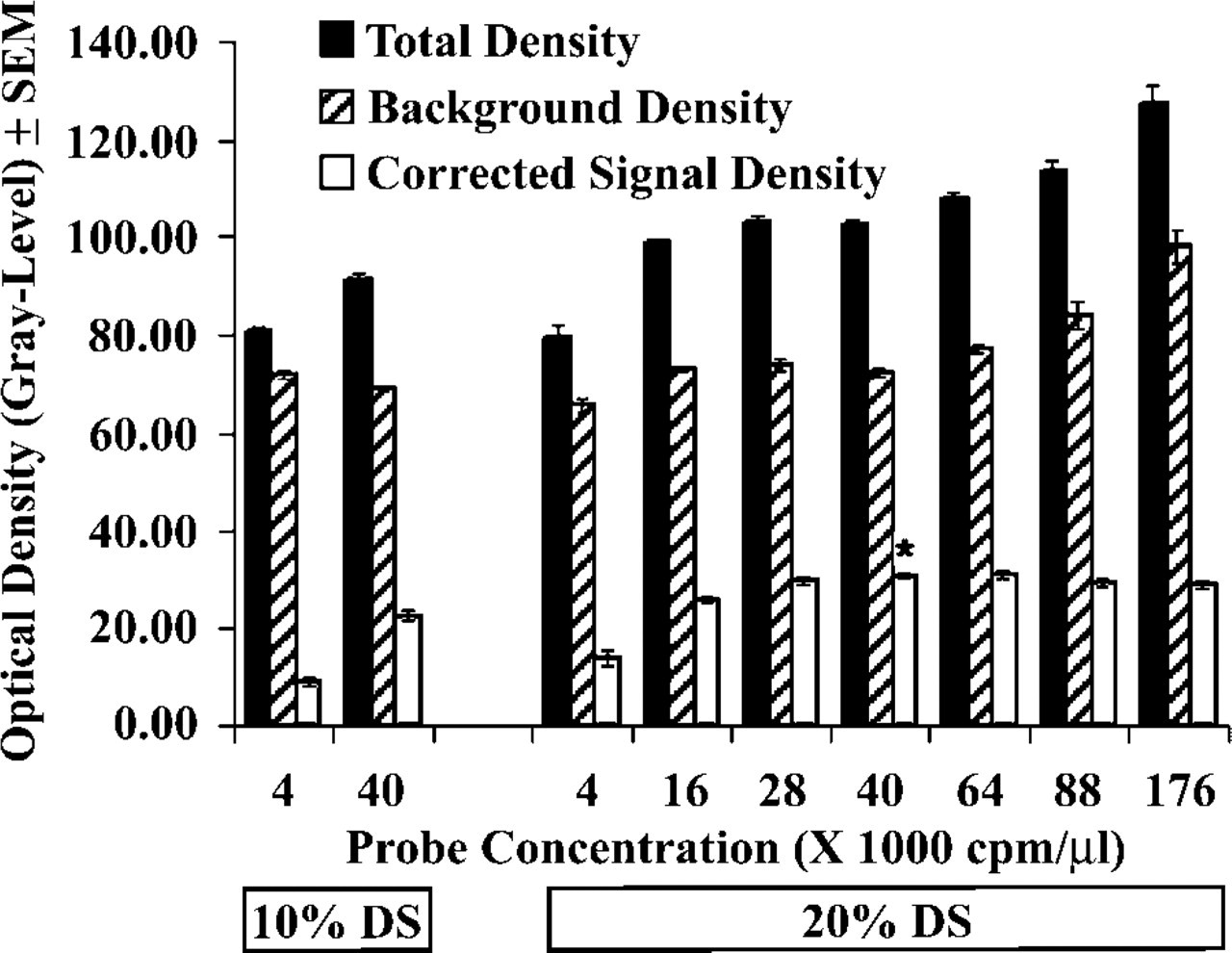
Total cortical density, background density and corrected signal density of ISH autoradiographs using increasing amounts of a low specific-activity PR probe for a saturation experiment. Compared to the standard procedure (here: 4,000 cpm/μl probe and 10% DS; first column), the maximal corrected signal density (asterisk) required the use of a tenfold probe excess (40,000 cpm/μl) and 20% instead of 10% DS in the hybridization solution. Note that background remained relatively stable below 64,000 cpm/μl probe, but then it suddenly increased, thus producing a slight reduction of corrected signal density at the highest probe concentrations.
The Rate of Hybrid Formation
Nucleic acid hybridization to immobilized targets follows pseudo-first-order kinetics if probe is in large access (Galau et al. 1977a, b). Despite its higher complexity, the ISHH reaction has been postulated to follow similar reaction kinetics (Tecott et al. 1987). It is interesting to note that the strong impact of probe concentration on hybridization signal, which we report here, is incompatible with pseudo-first-order reaction kinetics.
Probe Concentration
The enhancements of corrected signal density after application of increased probe concentrations were robust and reproducible using a variety of cRNA probes to different mRNA targets and of various lengths. Application of the low specific-activity PR probe in the presence of 1000 mM DTT revealed that a corrected density plateau was approached only with probe concentrations an order of magnitude higher than those recommended in published ISHH procedures and with 20%, instead of 10% DS, in the hybridization buffer, which further enhanced the effective probe concentration (Wahl et al. 1979). The use of increased probe and DTT concentrations represents a promising and novel approach to enhancing the sensitivity of in situ mRNA detection. Nevertheless, the high cost of the radioisotope and the increased problem of radioactive waste management are important issues that must be considered.
DS Concentration
Consistent with the concept that DS increases the effective probe concentration by excluding probe molecules from the volume occupied by the polymer (Wahl et al. 1979), when the same probe concentration was used the application of 20% or 30% DS in the hybridization buffer yielded consistently higher autoradiographic signals (corrected signal densities) than did 10% DS. Therefore, high amounts of DS or substitute macromolecules in the hybridization mixture offer efficient and inexpensive alternatives to the use of high-radioisotopic probe concentrations.
Detection Sensitivity
The practical value of using probe, DS, and DTT at increased concentrations will be the enhanced detection sensitivity of the isotopic ISHH procedure. This expectation is supported by our recent observation of ER-β mRNA expression in the majority of LHRH neurons, which required the simultaneous application of high probe, high DS, and high DTT in the hybridization solution (Hrabovszky et al. 2000). In our present studies using the GABA A γ2 cRNA probe, the advantage of these modifications was confirmed by the markedly improved clustering of silver grains over individual cortical neurons. In the same experiment, the difference we found between the lowest and the highest corrected signal density of film autoradiographs was comparable to a 10-fold difference in 14C radioisotope concentration on co-exposed radioisotope standards. Although without the use of appropriate tissue paste calibration standards (Davenport et al. 1988; Davenport 1998; Palfi et al. 1998; Vizi and Gulya 2000) and precise background correction methods, these data did not enable us to precisely determine the extent to which target mRNA molecules remained unoccupied using the standard hybridization procedure. Similarities between 14C plastic standards and 35S-labeled brain tissue paste standards (Miller 1991) indicate that the lowest probe and DS concentrations left most target mRNA molecules unhybridized in this study.
Autoradiographic Background
One must be aware that increased probe and DS may amplify hybridization artifacts, in addition to the advantage of enhancing detection sensitivity. Two distinct categories of nonspecific probe-tissue interactions we have to consider (Höfler et al. 1998) include the binding of probe to non-nucleic acid tissue constituents and the cross-hybridization of probes with nucleic acids exhibiting partial sequence homology with the target (Crabbe 1985). It has been proposed that the first type of background is largely contributed by the oxidation of tissue sulfhydryls, followed by the interaction of the resulting disulfides with the phosphorothioate moiety (O3P = 35S) of the probe (Zoeller et al. 1997). Whereas most hybridization procedures in the literature use DTT at 10–100 mM to minimize this type of background, results of our present studies indicate that DTT above 200 mM can further reduce nonspecific probe binding without any obvious disadvantage. The most dramatic effects of elevated DTT (750–1000 mM) could be observed when elevated concentrations of probe and/or DS were used.
The second important source of nonspecific probe binding is cross-hybridization of probes with rRNA and mRNA sequences that are partially homologous with the targeted mRNA (Crabbe 1985; Höfler et al. 1998; Sunday 1998). If hybridization stringency allows such cross-hybridization to occur, we reason that improved hybridization conditions will amplify nonspecific as well as specific hybridization events because these reactions are driven by identical chemical forces. In the light of this reasoning, it was important that high probe and DS concentrations did not generate cross-hybridization artifacts, as evidenced by the lack of neuronal-type density distribution on X-ray films when we used high concentrations of the sense-strand GABA-A γ2 RNA transcript. However, whenever high probe and DS concentrations are used one should consider the increased importance of specificity controls (Tecott et al. 1987; Davenport 1998; Höfler et al. 1998; Sunday 1998).
Use of Corrected Signal Density to Indicate the Completeness of Hybridization Reaction
Digital subtraction of background (e.g., background produced by a sense control probe; Davenport et al. 1988; Molenaar et al. 1993) from the total signal density is a commonly applied correction method to approximate the specific hybridization signal and, at the same time, to compensate for variations in autoradiographic background. Although this procedure has the great practical value of simplicity, it is important to recognize that variable background can introduce a new error to the quantification procedure. If nonspecific probe binding increases similarly within the signal area and within the background area, the enhancement of background density will exceed the increase of total density in the signal area, thus reducing the corrected signal density value as a net effect. This phenomenon results from the non-linear optical density response of the X-ray film to radiation (Kuhar et al. 1985; Davenport 1998). A second factor that causes corrected signal density to decrease in the presence of high background is the preferential accumulation of background in white matter structures (e.g., the corpus callosum) that we used here for background correction. In the light of the above two phenomena, we reason that as long as increasing amounts of probe cause a growing corrected signal density in spite of the opposite effects of rising background, the hybridization reaction is incomplete. Although we can use this argument to justify the application of “growing corrected signal density” to indicate that free probe binding sites are available for hybridization in the tissues, it is also important to recognize that this index is not very sensitive, despite the practical value of its simplicity. Unless background levels are stabilized using a wide range of increasing probe concentrations, the corrected signal density usually peaks much before the plateau phase of hybridization reaction (Nunez et al. 1989; Davenport 1998) is reached. This phenomenon may become a pitfall in studies attempting to correlate the maximal corrected signal density (or the maximal signal-to-background ratio) with the saturation of mRNA targets. In our saturation experiment, the simultaneous application of a low specific-activity PR probe and 1000 mM DTT in the hybridization solution ensured stable background levels that were unaffected by a broad range of probe concentrations. Therefore, probe concentrations generating a corrected signal density plateau were probably close to occupying most mRNA targets. It is interesting to note that saturation of hybridization signal in this experiment required 20% instead of 10% DS and probe concentrations one order of magnitude higher than those used in standard hybridization procedures.
To summarize our data, we demonstrated that the low efficiency of hybrid formation and the generation of high oxidative background may largely compromise the sensitivity of in situ mRNA detection using 35S-labeled cRNA probes. We suggest that the use of increased concentrations of 35S-labeled cRNA probe, DS, and DTT will greatly facilitate the cellular localization of rare mRNA species in brain and other tissues.
Footnotes
Acknowledgments
Supported by grant HD27305 from the National Institutes of Health (Bethesda, MD).
We wish to thank Drs C.D. Carpenter, R. Handa, O.K. Park-Sarge, and M.E. Vrontakis for their kind donation of cDNAs.