
Abstract
This study demonstrates the peculiarities of the glial organization of the optic nerve head (ONH) of a fish, the tench (Tinca tinca), by using immunohistochemistry and electron microscopy. We employed antibodies specific for the macroglial cells: glutamine synthetase (GS), glial fibrillary acidic protein (GFAP), and S100. We also used the N518 antibody to label the new ganglion cells' axons, which are continuously added to the fish retina, and the anti-proliferating cell nuclear antigen (PCNA) antibody to specifically locate dividing cells. We demonstrate a specific regional adaptation of the GS-S100-positive Müller cells' vitreal processes around the optic disc, strongly labeled with the anti-GFAP antibody. In direct contact with these Müller cells' vitreal processes, there are S100-positive astrocytes and S100-negative cells ultrastructurally identified as microglial cells. Moreover, a population of PCNA-positive cells, characterized as glioblasts, forms the limit between the retina and the optic nerve in a region homologous to the Kuhnt intermediary tissue of mammals. Finally, in the intraocular portion of the optic nerve there are differentiating oligodendrocytes arranged in rows. Both the glioblasts and the rows of developing cells could serve as a pool of glial elements for the continuous growth of the visual system.
T
The ONH possesses variations among vertebrates, because whereas some animals, such as primates, present a lamina cribrosa (Radius and Gonzales 1981), others, such as teleosts, are devoid of it and this portion is occupied by an elevated concentration of glial cells that helps to organize the ganglion cell axons (Fujita et al. 2000). Although such divergences among species suggest that the cytoarchitecture of the ONH intraocular portion should also be different in teleosts, this part of the visual system has been little analyzed (Easter et al. 1984; Vecino et al. 1997; Lara et al. 1998; Ott et al. 1998; Petrausch et al. 2000).
The main goal of the present study was to characterize the glial cells of the tench optic disc and the ONH intraocular portion. We describe their organization by using immunohistochemistry (IHC) and electron microscopic analyses and we look for some differences with the glial cells located in the ONH of mammals. We used anti-S100 and anti-GFAP antibodies, previously used in the visual system of teleosts, to identify glial cells (Dahl et al. 1985; Maggs and Scholes 1986; Levine 1989; Nona et al. 1989, 1992; Vecino et al. 1997; Velasco et al. 1997; Jimeno et al. 1999). We employed anti-PCNA (Bravo et al. 1987) to analyze the proliferative potential of a population of possible glial cell precursors that we detected in the intraocular portion of the ONH. We also used the anti-glutamine synthetase (GS), which is expressed exclusively in Müller cells in the neural retina of all vertebrates (Riepe and Norenburg 1977; Linser 1991), to try to elucidate the cellular environment in which the new axons, which we labeled with the N518 antibody (Ott et al. 1998), grow throughout the visual system of fish.
Materials and Methods
All procedures used in this work were in accordance with the guidelines of the European Communities Council Directive (86/609/EEC) and current Spanish legislation for the use and care of animals.
We employed 14 tench (Tinca tinca), 14–16 cm in body length, obtained from a commercial fish hatchery (Ipescón; Salamanca, Spain). Ten of them were used for IHC analysis and the rest for the ultrastructural study. All the animals were deeply anesthetized by using an aqueous solution of 0.03% tricaine methane sulfonate (MS-222; Sigma, St Louis, MO).
For the ultrastructural analysis, the fish were perfused transcardially with 150 ml of a solution containing 2.5% glutaraldehyde, 2% paraformaldehyde, 0.05% CaCl2 in 0.1 M cacodylate buffer, 0.18 M sucrose (pH 7.4). The eyes were removed, cut into small pieces, and stored in the same solution overnight at 4C. The pieces were washed in 0.1 M cacodylate buffer, 0.18 M sucrose (pH 7.4) and postfixed with 1% OsO4 in distilled water containing 1% potassium ferricyanide for 2 hr. Before dehydration, the pieces were subjected to block staining with 1% uranyl acetate in distilled water. Dehydration was performed using a graded series of cold acetone. Epon 812 (Taab; Reading, UK) was employed as embedding resin. Semithin sections (1 μm thick) were stained with toluidine blue and ultrathin sections were mounted on formvar-coated one-hole grids, contrasted with uranyl acetate and lead citrate, and studied using a ZEISS EM-900 electron microscope. Four different antibodies were used to examine the glial population of the ONH and retina. Rabbit anti-S100 (Dako; Carpinteria, CA), previously used in teleosts to label mature astrocytes, labeled part of the Müller cell population and horizontal cells (Vecino et al. 1997; Velasco et al. 1997). The mouse anti-GS antibody (Chemicon; Temecula, CA), previously used in fish (Mack et al. 1998; Peterson et al. 2001) labeled the entire population of Müller cells. The mouse anti-GFAP antibody (Sigma) has been previously used to label the Müller cells' and astrocytes' processes in the fish retina (Jimeno et al. 1999) while the mouse anti-PCNA (clone PC10; Santa Cruz Biotechnology, Santa Cruz, CA) antibody has been widely used to identify proliferating cells in fish (Negishi et al. 1991; Mack and Fernald 1995; Velasco et al. 2001). We employed the anti-PCNA antibody as a label for proliferating cells because it binds to the nuclear PCNA protein, a co-factor of DNA polymerase δ, usually detectable in G1, S, and G2 cells (Bravo et al. 1987). We have also used a mouse monoclonal antibody, N518 (a generous gift from Dr. C.A.O. Stuermer), to specifically detect the fish protein neurolin localized in the newly formed RGC axons. Electron microscopy enabled us to describe ultrastructural differences among the diverse glial cell types and particularly to characterize microglial cells.
The specimens were anesthetized and quickly decapitated. The eyes were removed and fixed for 12 hr at 4C in a solution of 95% ethanol and 40% formaldehyde (9:1). The whole eyes were then rinsed in phosphate buffer 0.1 M, pH 7.4 (PB) and soaked in 30% sucrose in PB overnight. For the single labeling with the anti-GFAP antibody and for the double labeling with GS-S100, GFAP-S100, and N518–S100, we employed a different fixative. After decapitation the eyes were immersed in a solution of 4% paraformaldehyde in PB at 4C overnight. The eyes were then rinsed in PB and soaked in 30% sucrose in PB overnight. Then the eyes were frozen in liquid nitrogen and cut on a cryostat in 16-μm-thick nasotemporal sections. These were mounted on gelatin-coated slides.
The sections were rinsed three times in PBS 0.1 M, pH 7.4, with 0.2% Triton X-100 (PBS-Tx) before IHC processing. Endogenous peroxidase activity was blocked with 2% H2O2 in 100% methanol for 10 min. For the immunolabeling, the sections were first incubated with 2% normal horse serum in PBS-Tx (3 hr). Some slides were then incubated with the anti-PCNA or with the anti-GFAP monoclonal antibodies diluted at 1:500 and at 1:400, respectively, in a solution of PBS-Tx-serum with 1% dimethylsulfoxide (DMSO) overnight in a humidity chamber. To achieve double immunolabeling of GS-S100, GFAP-S100, and N518–S100, the same slides were incubated in the same buffered solution containing a mixture of the anti-S100 polyclonal rabbit antibody at 1:500, the anti-GS monoclonal antibody at 1:1000, the anti-GFAP antibody at 1:400, or the anti-N518 monoclonal antibody at 1:10.000 overnight in a humidity chamber.
After the latter incubations, all the slides were washed three times in PBS-Tx and, for the PCNA and GFAP labeling, sequentially incubated at room temperature (RT) with a biotinylated anti-mouse IgG antibody (Vector; Burlingame, CA) at a dilution of 1:200 and an avidin-biotin-peroxidase complex (Vector) (1:250). Detection of both immunoreactions were carried out with a solution of 0.05% 3.3′-diaminobenzidine tetrahydrochloride (DAB) and 0.025% H2O2 in 0.1 M Tris-HCl (pH 7.5) for 10 min at RT.
The sections for the double immunolabeling with GSS100, GFAP-S100, and N518–S100 were incubated with a mixture of the Cy3-conjugated fluorescent anti-rabbit IgG antibody (Jackson Immunoresearch; Avondale, PA) at a dilution of 1:1000 for detection of the S100 protein, and the Cy2-conjugated fluorescent anti-mouse IgG antibody (Jackson Immunoresearch) at a dilution of 1:1000 for detection of the GS, GFAP, or the neurolin protein. Finally, some of the sections were stained with DAPI (Sigma) at a dilution of 1:10,000 in distilled water to label all the cell nuclei. They were then dehydrated and mounted in Entellan (Merck; Darmstadt, Germany).
Some of the images from light microscopy were obtained with an Olympus Apogee digital camera coupled to an Olympus AX-70 photomicroscope with the appropriate fluorescent filter settings. The Cy3 fluorescent dye was visualized with a BP 510–550-nm wavelength filter, the Cy2 dye with a BP 470–490 filter, and the DAPI labeling with a BP 360–370 filter. The rest of the fluorescent images were obtained with a laser scanning spectral confocal microscope (Leica TCS SP2) using simultaneous excitation of fluorochromes with laser wavelengths of 488 and 543 nm and a filter-free prism spectrophotometer. Original pictures were further processed with Adobe Photoshop 5.5 software (Tucson, AZ) to obtain the optimal contrast within the same figure plate.
Results
We analyzed the glial organization of three different zones of the tench ONH, which is composed of the optic disc and the intraocular portion. For Zone 1, in the optic disc we studied the portion where the RGC axons coming from the NFL of the retina turn to go into the optic nerve (Figure 1A). For Zone 2, in the intraocular portion of the ONH we analyzed the interface between the retina and the optic nerve at the level of the limit with the choroid membrane (Figures 1A and 7), which in mammals is denominated the Kuhnt and Jacoby intermediary tissue. For Zone 3, in the intraocular portion of the ONH, we analyzed the portion of optic nerve at the interface with the sclera, which corresponds to the lamina cribrosa in mammals. In this zone, we also studied the rows of cells located among the RGC axons (Figures 1A and 7).
The Optic Disc: Zone 1
The analysis of the semithin sections from the optic disc (Figures 1A–1C), together with the GS, S100, and GFAP IHC labeling (Figures 2A, 2B, 3A, and 3B) demonstrated that the Müller cells within this area present some variations in their morphology and arrangement compared to those that they present in the rest of the neural retina. The somata and cytoplasmic processes of the entire population of the Müller cells of the tench retina were GS positive (GS+) (Figure 2A). The GS labeling of this population at the level of the optic disc (Figures 2A and 2C) demonstrated that the Müller cells' vitreal processes form a barrier at the interface between the retina and the optic nerve. At the same site, some of these long Müller cells' processes were strongly labeled with anti-S100 (Figures 2C, 3A, and 3B) and anti-GFAP (Figures 2B and 3A).
The semithin and ultrathin sections showed that the vitreal processes of the Müller cells located around the optic disc were quite thick and were surrounded by several cell somata that were directly apposed to them (Figures 1B and 1C). This is a feature that is not observed in the rest of the Müller cells of the tench retina. The ultrastructural analysis showed that the vitreal processes of the Müller cells were composed of several cytoplasmic prolongations (Figure 4B), while in the rest of the retina they were not grouped. They were not electron-dense and had many bundles of densely packed intermediate filaments, some light mitochondria, many cisternae of smooth endoplasmic reticulum, and many light vesicles adjacent to the cytoplasmic membrane (Figure 4B).
The IHC and ultrastructural analyses also permitted the characterization of the glial cell types found adjacent to the Müller cells' processes in the optic disc (Figures 1C, 4A, 4C, 5A, and 5B). Their cell bodies were arranged along the entire length of the Müller cells' vitreal processes, but most of them were located close to the inner limiting membrane (ILM) (Figure 4C). The morphological differences suggest the existence of at least two different glial types. Ultrastructurally, some of these cells had a very euchromatic kidney-shaped nucleus and contained abundant perinuclear cytoplasm, more electron-dense than the adjacent Müller cell cytoplasm. They possessed large somata from which various cytoplasmic prolongations emerged in several directions. Some of these processes divided the RGC axons into small fascicles at the level of the ONH and others ran parallel to the Müller cell processes, perpendicularly to the RGC axons in the NFL (Figure 4C). The last were joined by desmosomes and contained many bundles of intermediate filaments in the cytoplasm, typical features of astrocytes. The S100 labeling of this region also demonstrated the presence of S100-positive (S100+) mature astrocytes directly apposed to the Müller cells' arched processes (Figure 4A). This close relationship between astrocytes and Müller cells occurs only at the level of the optic disc, not in the rest of the retina. The other glial cell type found around the Müller cells' processes was S100-negative (S100-) and has been ultrastructurally characterized. They had small fusiform somata (more electron-dense than the astrocytes' somata) from which some short thin processes emerged (Figures 5A and 5B). These cytoplasmic prolongations were arranged parallel to the RGC axons. Their nuclei were very heterochromatic and were surrounded by a thin rim of cytoplasm that contained free ribosomes, short and dilated cisternae of rough endoplasmic reticulum, a developed Golgi apparatus, small mitochondria, and some clear vesicles (Figure 5B). These ultrastructural characteristics are those of resident microglial cells, which are also arranged in a similar way in the NFL of the rest of the neural retina.
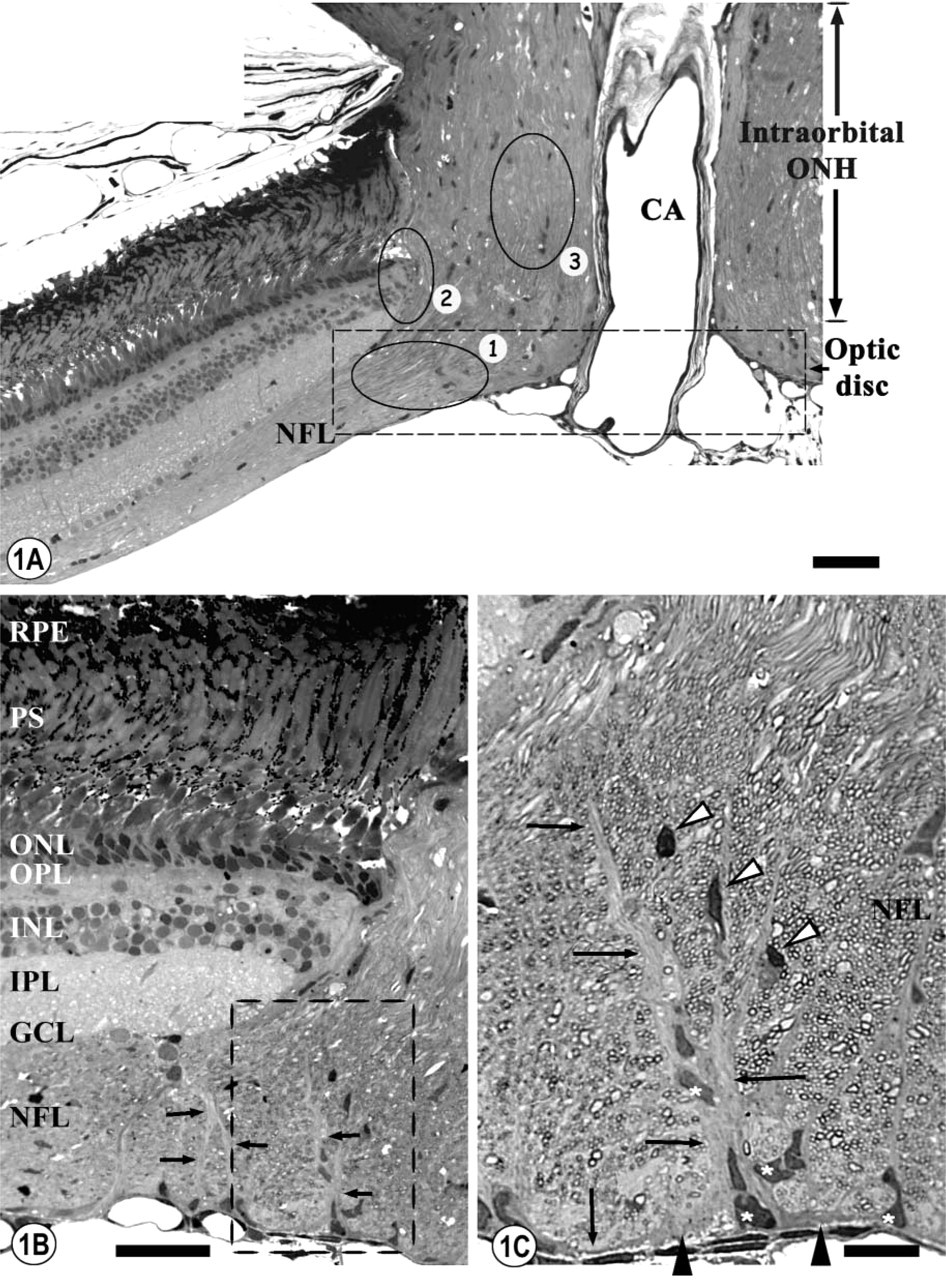
Semithin sections from the ONH.
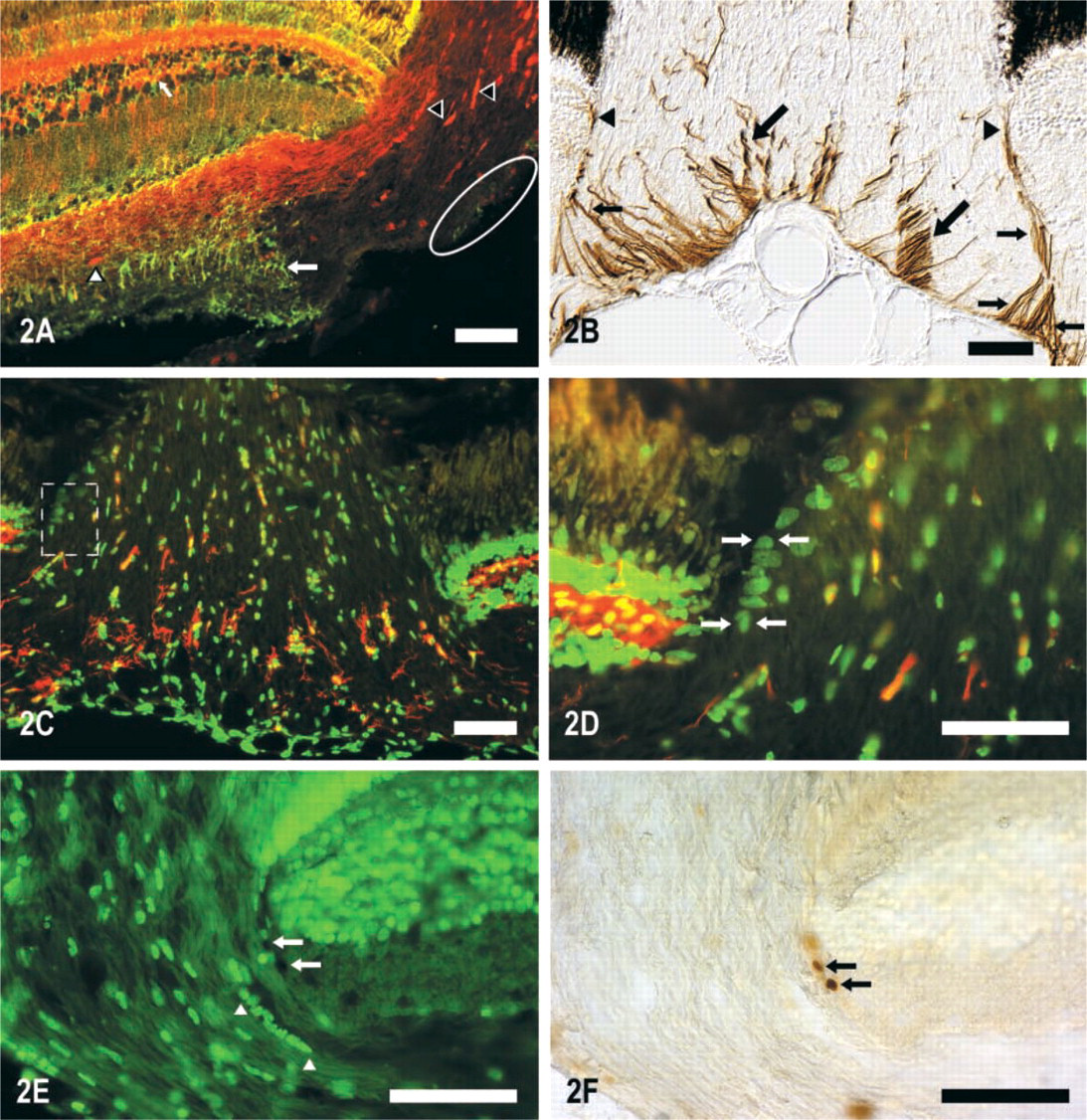
Cryostat sections of the ONH immunolabeled with different antibodies.
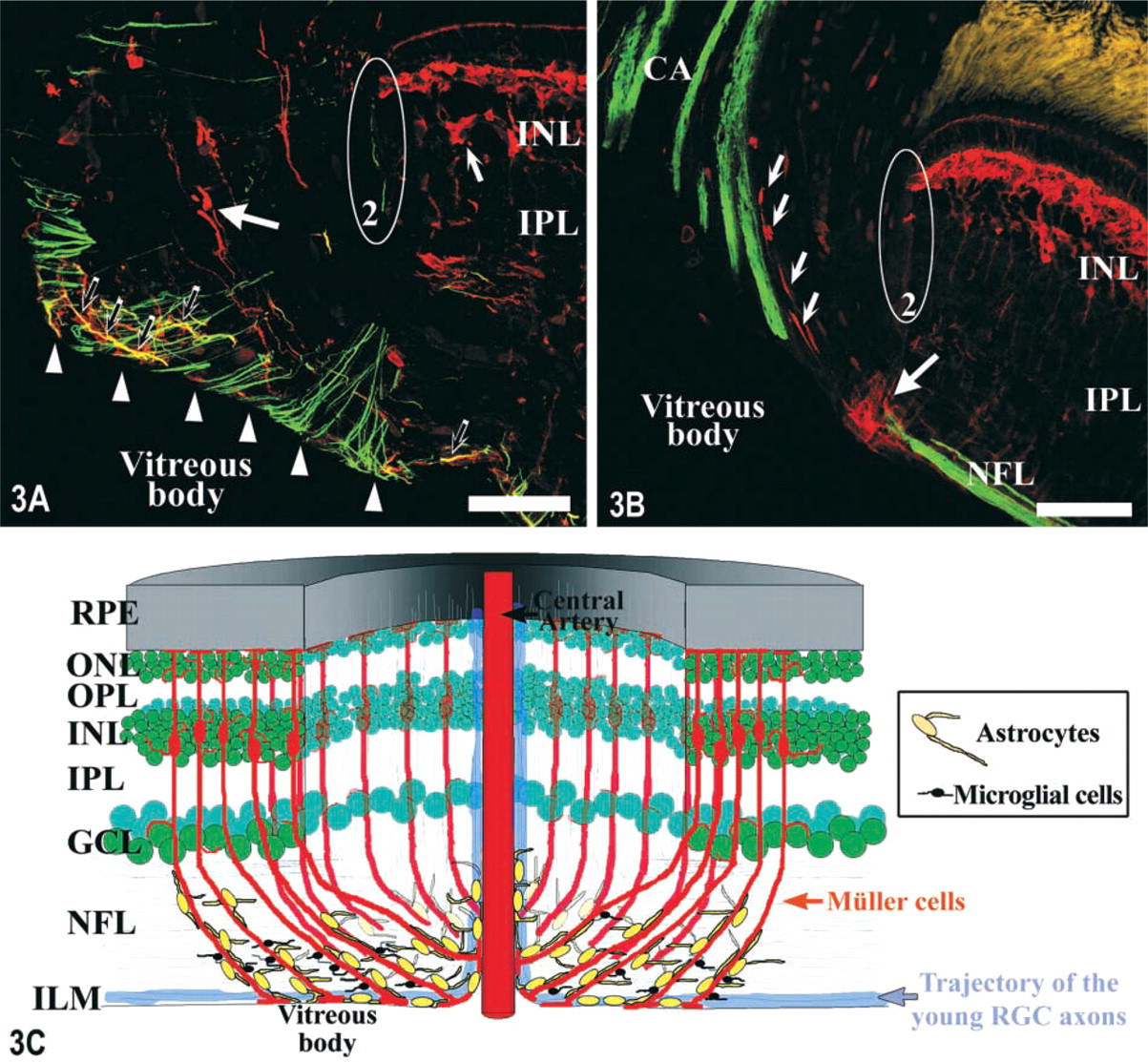
Cryostat sections of the ONH immunolabeled with different antibodies and visualized with a confocal microscope.
The S100 (Figures 2A, 3A, and 3B) and GFAP (Figure 2B) immunolabelings also demonstrated the presence of many S100+ and GFAP-positive (GFAP+) mature astrocytes in the center of the ONH that did not appear to be associated with the Müller cells' vitreal processes of the optic disc. The GFAP labeling showed that the gliofilaments of the cytoplasmic processes located in this region of the optic disc, probably from the astrocyte population located in this zone, presented a stronger labeling than the rest of the retina (Figures 2B and 3A). The GFAP-S100 double immunolabeling (Figure 3A) also detected GFAP+-S100+ processes in the optic disc (yellow in Figure 3A) that seemed to belong to astrocytes located close to the ILM. The N518–S100 double labeling permitted the detection of some of the S100+ astrocytes from this zone of the optic disc. These were closely associated with the new RGC axons, which were N 518-positive (N518+) and ran through the most vitreal part of the NFL at the level of the optic disc (Figure 3B).
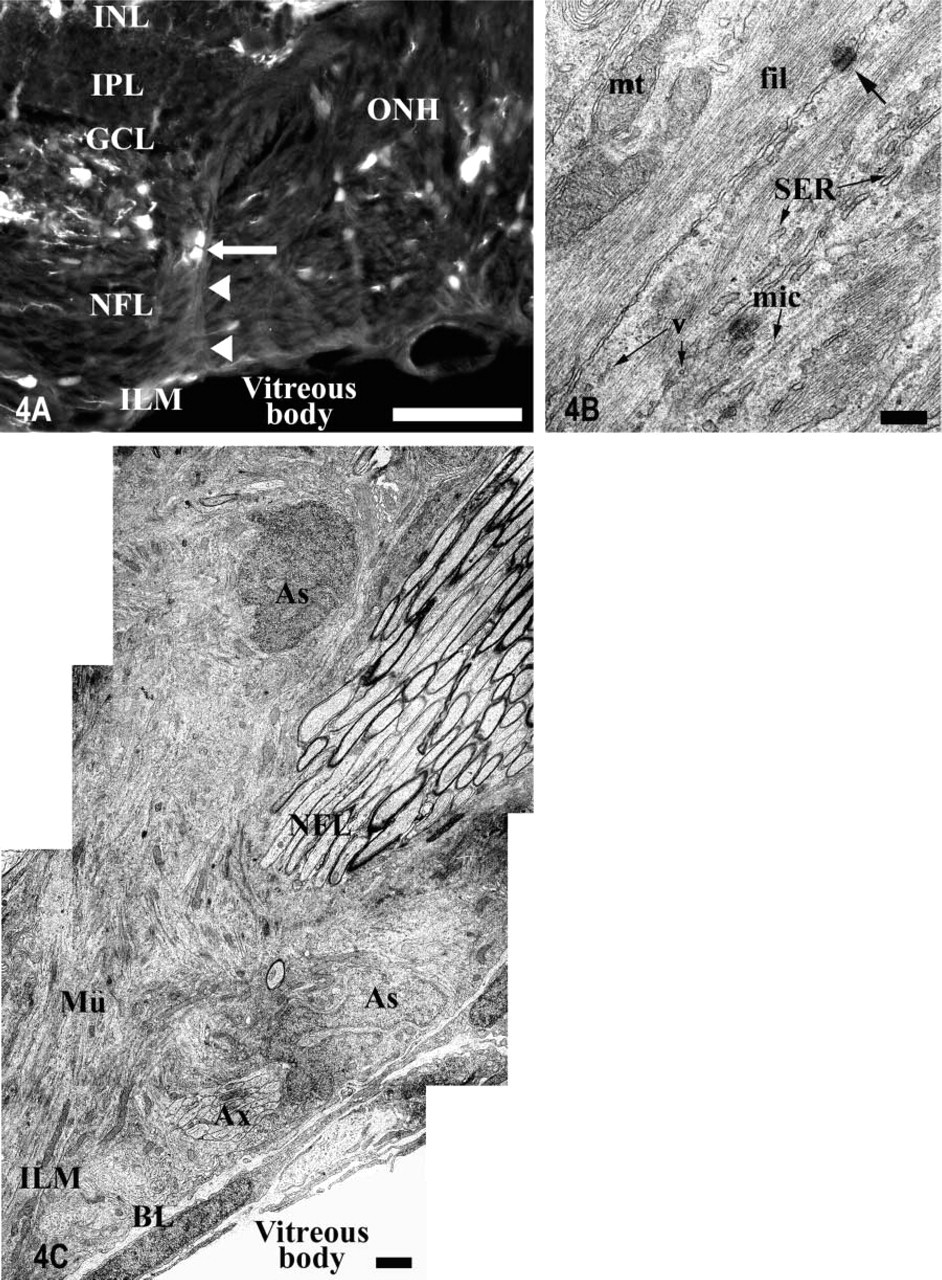
Light and electron micrographs of the glia limitans showing the association between Müller cells and astrocytes at the level of the optic disc (Zone 1 in Figure 1).
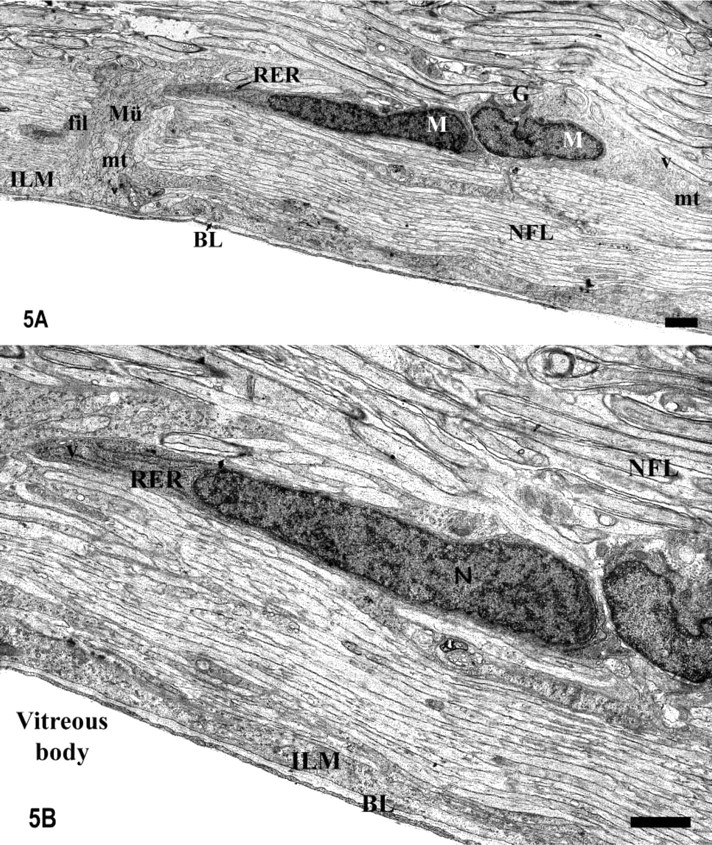
Electron micrographs of the glia limitans at the level of the optic disc.
The organization of the Müller cells, astrocytes, and microglial cells in the optic disc is summarized in Figure 3E. It shows a complete view of the spatial distribution of these glial elements and their relationships with the newly formed RGC axons.
Intraocular ONH
The intraocular ONH is the region where the RGC axons have already changed their orientation by 90° and start to run parallel to the central artery of the optic nerve (Figure 1A).
Zone 2. In this zone there was a group of densely packed cells and many cytoplasmic prolongations located at the level of the choroid of the retina that constituted a real barrier between the neural retina and the optic nerve (Figures 1A and 6A). The cell bodies were in close contact with the retinal layers. They were fusiform and were aligned in a sclero-vitreal orientation and had a very euchromatic nucleus with the same orientation (Figure 6B). Some of these cells were labeled with the anti-PCNA antibody, indicating that they possessed the ability to proliferate (Figures 2E and 2F). Moreover, these cells did not express S100 (Figures 2C, 2D, and 3B) or GFAP in contrast to the mature astrocytes previously shown in the optic disc (Figures 2C, 2D, 3A, and 3B). Their ultrastructural analysis showed that the nuclear envelope had many ribosomes attached to it and the perinuclear cytoplasm contained free ribosomes, many microtubules, short cisternae of rough endoplasmic reticulum with a flocculent material, some gliofilaments, and they were joined by desmosomes (Figure 6C). These are features of immature glial cells, presumably astroblasts. The cytoplasmic processes located between the astroblasts and the RGC axons running through the optic nerve were GFAP+ (Figures 2B, 3A, and 6B). These processes were more electron-dense than the cytoplasm of the astroblasts and had many bundles of intermediate filaments, some pale and elongated mitochondria, and were joined by desmosomes and tight junctions (Figures 6B and 6D). These features may show that the cytoplasmic processes observed in the interface between the retina and the optic nerve tissue do not belong to the adjacent astroblast but appear to emerge from the astrocytes that form the glia limitans between the meninges and the neural tissue. It is noteworthy that there was no basal membrane to separate these glial cells from the retinal cells, so they were in direct contact with them (Figure 6B). Moreover, there was no sheet of connective tissue to divide the retina from the optic nerve (Figures 6B and 6D). Therefore, the limit between the intraocular portion of the optic nerve and the retina at the level of its choroid membrane is composed of astroblasts and mature astrocytes' processes.
Zone 3. Regarding the typology of the remainder of the glial cells located in the intraocular portion of the tench ONH, we have shown in previous works (Lara et al. 1998; Lillo et al. 1998) that most of these glial cells are arranged in rows, parallel to the RGC axons, which are composed of mature oligodendrocytes and reticular astrocytes. Although, as described above, we found rows composed by different types of glial cells, the majority of them were constituted of glial cells with the same morphological features (Figure 7A). The IHC and ultrastructural analyses showed that some of the homogeneous rows were composed of S100+ reticular astrocytes (Figures 2A, 3B, and 7D) (Maggs and Scholes 1990). They possessed a very euchromatic nucleus, thick bundles of intermediate filaments within their cytoplasm, and were joined by desmosomes (Figure 7D). Moreover, we found some other rows composed of several medium oligodendrocytes that possessed medium electron-density and a different ultrastructure from astrocytes. These cells had a heterochromatic and polygonal nucleus; they possessed some microtubules and lacked intermediate filaments (Figures 7B and 7C). It was possible to differentiate the medium oligodendrocytes from the astrocytes because they were surrounded by astrocytic processes filled with intermediate filaments and joined by desmosomes, which ran parallel to the RGC axons (Figures 7B and 7C).
Discussion
In this work, ultrastructural and immunohistochemical analyses have permitted the demonstration of several interesting and previously unreported peculiarities of the glial cell populations in the optic nerve head of the tench.
The GS and GFAP labelings, and also the ultrastructural analysis of the Müller cells' vitreal end-feet, showed a specific spatial arrangement of these cytoplasmic processes around the optic disc with which many cell bodies of astrocytes (S100+) and microglial cells are associated. The PCNA labeling of proliferative cells has enabled us to identify a population of glioblasts forming a glial barrier between the retina and the optic nerve at the level of the intraocular portion of the optic nerve head. Finally, we detected rows of glial cells within the optic nerve head whose ultrastructural analysis has demonstrated that most of them are composed of differentiating oligodendrocytes.
The present work adds new data to those obtained in previous works related to the glial cells located within the fish ONH. These works suggest that these cells possess a pattern of protein expression that differs from that of the rest of the optic nerve, mainly those proteins related to the glial cytoskeleton, such as GFAP (Jimeno et al. 1999) and cytokeratin expression (Maggs and Scholes 1990; Scholes 1991). These observations indicate that the glial elements from the visual system of fish are a heterogeneous population and also that the glial cells of the ONH are different from those located in the same region in mammals.
Although the GS is present in the CNS astrocytes of vertebrates (Norenberg and Martinez-Hernandez 1979; Linser 1985), our results demonstrate that GS expression in the tench retina is restricted to the Müller cells, supporting previous works that showed the same pattern of expression in other fish retinas (Mack et al. 1998; Peterson et al. 2001). Within the NFL, the vitreal Müller cell processes separate the axon bundles by interfascicular septa (Prada et al. 1989), and in addition the GS staining and the ultrastructural analysis of the optic disc of the tench reveal that within this area they acquire specific morphological modifications. The analysis of the Müller cells' vitreal processes, as well as astrocytes and microglial cells associated with them, by using IHC labeling against GS, GFAP, and S100 proteins and also by electron microscopy, allowed us to determine that they create an elaborate and arched network around the optic disc that appears to be a real barrier between the retina and the ONH through which the RGC axons can pass (Figure 3E). A few studies have analyzed the ONH in teleosts (Easter et al. 1984; Bastmeyer et al. 1995; Vecino et al. 1997; Ott et al. 1998; Petrausch et al. 2000), but none of them describes the particular distribution of the vitreal processes of the Müller cells that we have shown in the optic disc of tench. This area appears to be exposed to several mechanical forces because it is a transition zone between the neural retina and the optic nerve. The network of the Müller cell processes could act as a resistant support for the mechanical movements (Reichenbach and Robinson 1995). To date, there is only one study (in chicken) that describes a similar regional adaptation of the Müller cells in the optic disc, where they form an organized arched structure (Prada et al. 1989). The cytoplasmic content of the Müller cells in the optic disc is similar to that of these cells in the rest of the retina, so they should perform the same function, allowing the diffusion of substances from the extraretinal space to the retinal cells. Furthermore, they could act as a barrier between the elements in the retina and the optic nerve. All these observations would support the hypothesis that Müller cells modify their morphology and functional behavior depending on the region at which they are located, as has been demonstrated in the retina of other animals (Reichenbach and Wohlrab 1986; Prada et al. 1989).
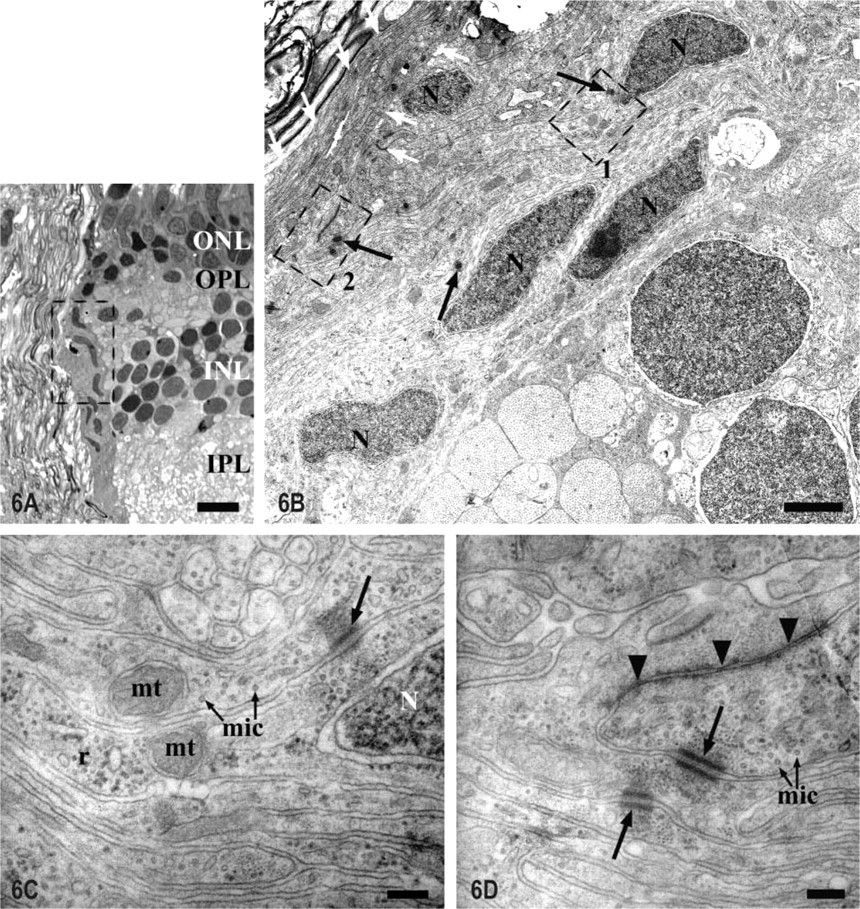
Light and electron micrographs of the ONH intraocular portion (Zone 2 in Figure 1).
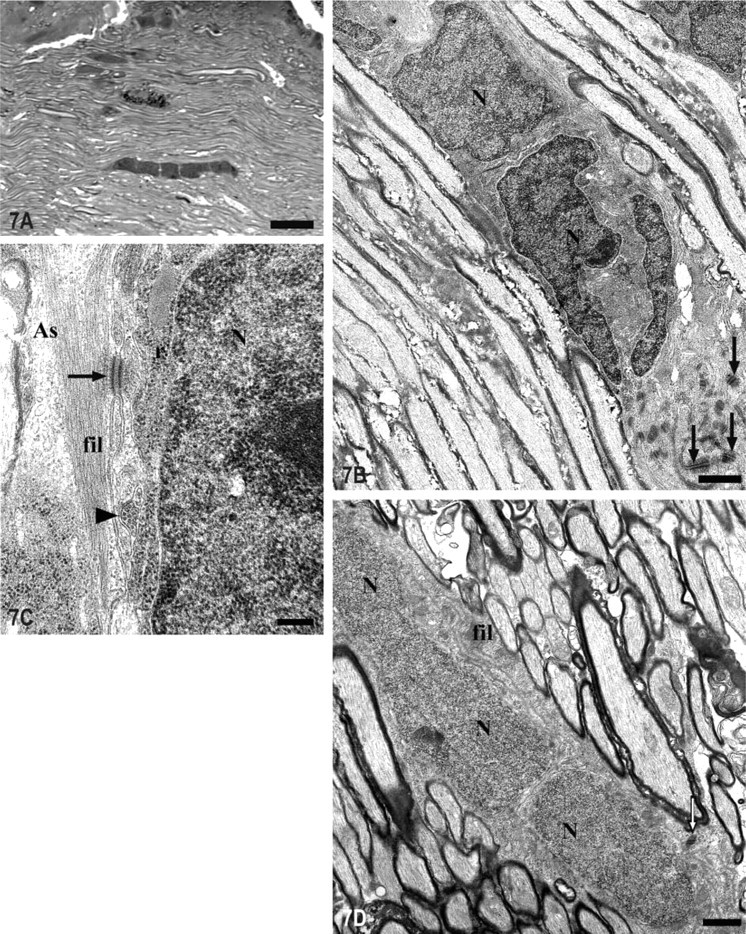
Light and electron micrographs of rows of cells formed by homogeneous glial somata in the intraocular portion of the ONH (Zone 3 in Figure 1).
Ultrastructural analysis of the optic disc of tench demonstrates the presence of astrocytes and microglial cells in close relation to the Müller cell processes. This special cell grouping surrounding the Müller cells' processes is exclusive to this region and, to our knowledge, has not been described before in the ONH of vertebrates. The astrocytes that we detected in the tench ONH share their arrangement and features with the reticular astrocytes that Maggs and Scholes (1990) first described in the optic nerve of other teleosts, although this is the first time they have been characterized in the ONH of fish.
Because RGC axons are continually added to the optic nerve (Johns and Easter 1977), there is always a pool of growth cones in the ONH that necessarily must cross the glial network that we have described here. IHC studies carried out in the goldfish ONH demonstrated the existence of a netrin-positive ring of cells around the optic disc, which is probably related to the RGC axons' growth (Petrausch et al. 2000; Stuermer and Bastmeyer 2000). Our IHC and ultrastructural analyses of the tench optic disc show the presence of many S100+ astrocytes located around the optic disc, most of them in the vitreal part of the NFL in the same area where the ring of cells exists in goldfish. As shown in Figures 3C and 3D, it is suggested that the S100+ astrocytes could wrap the N518+ new ganglion cells' axons that run along this retinal layer, so they could be responsible for guiding and orientating them to the ONH. The above-mentioned population of astrocytes has also been observed in the optic disc of goldfish (M. Bastmeyer, personal communication).
Previous works carried out by our group have described microglial cells in the NFL and optic disc of the tench by using specific markers for this cell type (NDPase and tomato lectin) (Jimeno et al. 1999; Velasco et al. 1999), but none of them had detected microglial cells apposed to the Müller cell processes at the optic disc. We consider, taking as a reference those previous studies and the ultrastructural analysis of this glial cell type carried out in the present work, that we are able to precisely identify these elements as microglial cells. In the NFL of the quail retina a similar association has been observed between the microglial cells and the Müller cells' processes (Navascués et al. 1994). These observations, and the proximity of the microglial cells to the optic nerve central artery, lead us to suggest that these cells may come from this vessel and enter the neural retina. In relation to this, it is known that the number of astroglia in the NFL in other species is proportional to the thickness of this layer in the different regions of the retina (Anderson 1969; Büssow 1980; Triviño et al. 1996), and their occurrence also coincides with the presence of blood vessels (Schnitzer 1985; Schuck et al. 2000). The large number of astrocytes in the optic disc surrounding the central artery seems to support the theory that, in mammals, the astrocytes of the retina also originate from the blood vessels of the optic nerve (Stone and Dreher 1987; Watanabe and Raff 1988).
In the intraocular portion of the ONH of animals that lack or present a poorly developed lamina cribrosa, as in mouse (Fujita et al. 2000), rat (Wolburg and Buerle 1993) and turtle (Dávila et al. 1987), this area is occupied by several thick astrocytic processes. In a previous IHC study of the tench ONH, the authors described many cells around the optic disc that are S100+ (Vecino et al. 1997) and suggested that these cells could form an interface between the retina and the optic nerve. Moreover, in a previous study of the GFAP labeling pattern in the visual pathway of the goldfish (Levine 1989), it was shown that both the astrocytes of the ONH and those of the optic tract are GFAP+, although the astrocytes of the rest of the optic nerve are GFAP-negative (GFAP-). This observation, together with the knowledge that the anti-GFAP antibody labels the Müller cell processes (Levine 1989; Nona et al. 1989), permits a greater comprehension of the description of the glial arrangement in the ONH.
In the intraocular region of the ONH, at the interface between the retina and the optic nerve, we detected a group of PCNA-positive (PCNA+) proliferating cells that had not been previously described, whose ultrastructure indicates that they are probably astroblasts (Trimmer et al. 1982; Kalnins et al. 1984; Monzón-Mayor et al. 1990a). Attached to this group of GFAP- astroblasts we have also identified bundles of GFAP+ glial processes that correlate with the cytoplasmic prolongations of the astrocytes that organize the glia limitans of all the optic nerve. This is in accordance with their content in gliofilaments, which is more prominent than in the neighboring astroblasts. We infer that these astroblasts could be a pool of young astrocytes that could migrate from this location to the optic nerve. It is possible that these astroblasts constitute a source for the maintenance of the glial population in the optic nerve because they are constantly needed to support the newly formed ganglion cells' axons. In animals with a lamina cribrosa, there is a group of cells known as Kuhnt intermediary tissue, formed only by mature astrocytes, at the same location where the pool of astroblasts has been found (Okinami et al. 1976; Triviño et al. 1996; Schuck et al. 2000). Similarly to the astroblasts of tench, these astrocytes also form a barrier between the retina and the optic nerve. Although in some animals these astrocytes appear surrounding the blood vessels and form a basal lamina that separates the neural from the connective tissue (Schuck et al. 2000), we did not observe that the astroblasts in tench form any kind of basal lamina because they are not related to blood vessels in this region. Schuck et al. (2000) reported that, at an early stage of the development of the chicken visual system, there are immature glial cells that typically express vimentin at the same region of interface between the optic nerve and the retina. These authors demonstrated that, during further embryonic development, these cells are special spindle-shaped glial cells that represent a heterogeneous population because some of them begin to express GFAP at the same time as others are still expressing vimentin or both (Schuck et al. 2000). This observation could support the idea that, in species that contain a continuous immature region in their visual systems, this region, in the limit between the retina and the optic nerve, could remain in an immature state throughout life.
In the intraocular portion of the ONH we also detected and ultrastructurally characterized rows of glial cells. Many of these were composed of several oligodendrocytes in the process of differentiation. Because there are no antibodies to specifically detect the oligodendrocyte population in fish tissue sections, in the present work we have verified their existence taking as a reference their ultrastructural features (Mori and Leblond 1970; Monzón-Mayor et al. 1990b; Peters et al. 1991; Lillo et al. 1998). These immature oligodendrocytes have been recognized in the developing optic nerve of mammals (Skoff et al. 1976; Raff et al. 1984) and anamniote vertebrates (Stensaas 1977), but they have never before been observed in the optic nerve of an adult animal. We infer that the continuous formation of new oligodendrocytes in the continuously growing optic nerve of fish may be an important process for the myelination of the constantly added new ganglion cell axons. Although several studies have detected maturing oligodendrocytes in the regenerating optic nerve of teleosts after a lesion (Wolburg 1978, 1981; Bastmeyer et al. 1991, 1993), and although it is known that continuous gliogenesis occurs in the adult optic nerve of fish in the zone where the new axons run (Maggs and Scholes 1986, 1990), the identification of these rows of maturing oligodendrocytes has been ambiguous until now.
Footnotes
Acknowledgments
Supported by Junta de Castilla y León and F.S.E (grant numbers SA103/01, SA30/99), by the Spanish MCyT, and by DGI.
We thank Mr. G. H. Jenkins for revising the English version.