
Abstract
We examined the distribution of annexin 6 (ANX6) in rat brain with immuno-histochemistry (IHC). Several neuronal cell populations were intensely labeled with the ANX6 monoclonal antibody (MAb), including layer 5 of neocortex, the lateral septum, the lateral hypothalamic area, the red nucleus, and the Purkinje cell layer in cerebellum. Neuronal immunolabeling was localized to the nucleus and the cytosol. Darkly stained ANX6-immunoreactive glia, with the morphology characteristic of astrocytes, were abundant in the hippocampus, substantia nigra reticulata, and cerebellum. Evidence suggests that ANX6 may function in neuronal and glial calcium-dependent processes.
A
One of the most abundant annexins in the mammalian brain, as determined by biochemical analyses, is annexin 6 (ANX6; Woolgar et al. 1990). Neuroanatomic studies of ANX6 immunoreactivity (IR) in the mammalian brain have been limited to the cerebellum, dorsal root ganglia, and spinal cord (Woolgar et al. 1990; Naciff et al. 1996). In the porcine cerebellum, ANX6-IR is present in Purkinje, basket, and stellate neurons but not in glial cells (Woolgar et al. 1990). Most sensory neurons in the rat dorsal root ganglia show immunostaining for ANX6 (Naciff et al. 1996). ANX6-IR has also been detected in large motor neurons in the ventral horn of the rat spinal cord (Nacif et al. 1996). In the present study we examined the distribution of ANX6-IR in rat brain.
All animal procedures in this study were approved by the Institutional Animal Care and Use Committee (IACUC) at the Indiana University School of Medicine. Twelve adult male Sprague-Dawley albino rats (Harlan; Indianapolis, IN), weighing 300-400 g, were maintained under a 12:12-hr light-dark cycle with food and water freely available. Each animal was deeply anesthetized with sodium pentobarbital (100-150 mg/kg, IP) and perfused through the ascending aorta with 150 ml of heparinized saline, followed by 500 ml of 4% paraformaldehyde in 0.1 M sodium phosphate buffer, pH 7.4. After fixation the brains were removed, allowed to sink overnight in a 30% sucrose solution, and cut into 30-μm sections on a freezing microtome.
A one-in-three series of sections was preincubated for 1 hr in 2% normal donkey serum (NDS), 2% bovine serum albumin (BSA), and 0.3% Triton-X (TX) in 0.01 M PBS, pH 7.4. The sections were incubated with mouse anti-ANX6 (ICN; Aurora, OH), diluted 1:500 in PBS-NDS-BSA-TX, with gentle agitation for 2-4 days at 4C. Next, the sections were rinsed in PBS, incubated in biotinylated SP-conjugated donkey anti-mouse IgG (Jackson; West Grove, PA), diluted 1:200 in PBS-NDS-TX for 2 hr, rinsed again, and reacted with ABC reagent (Vector; Burlingame, CA) for 2 hr. After several rinses in PBS, the sections were reacted in a solution of 0.035% 3-3′ diaminobenzidine tetrahydrochloride (DAB), 2.5% nickel ammonium sulfate, and 0.3% H2O2 in 0.1 M sodium acetate buffer, pH 6.0, for 1 min. The sections were rinsed in PBS and mounted on gelatin-coated slides. The slides were cleared in xylene and coverslipped with Permount (Fisher; Pittsburgh, PA).
The anti-ANX6 monoclonal antibody (MAb) (clone 73; available from both ICN and BD Transduction Laboratories, San Diego, CA) was generated against ANX6 isolated from human placenta and specifically recognizes ANX6 in a variety of tissues (Goulding et al. 1995; Barwise and Walker 1996; Babiychuk et al. 1999). The specificity of the antibody was further tested by omission of the primary antibody and substitution of normal mouse serum for the primary antibody. No specific staining was observed in either of these two control experiments.
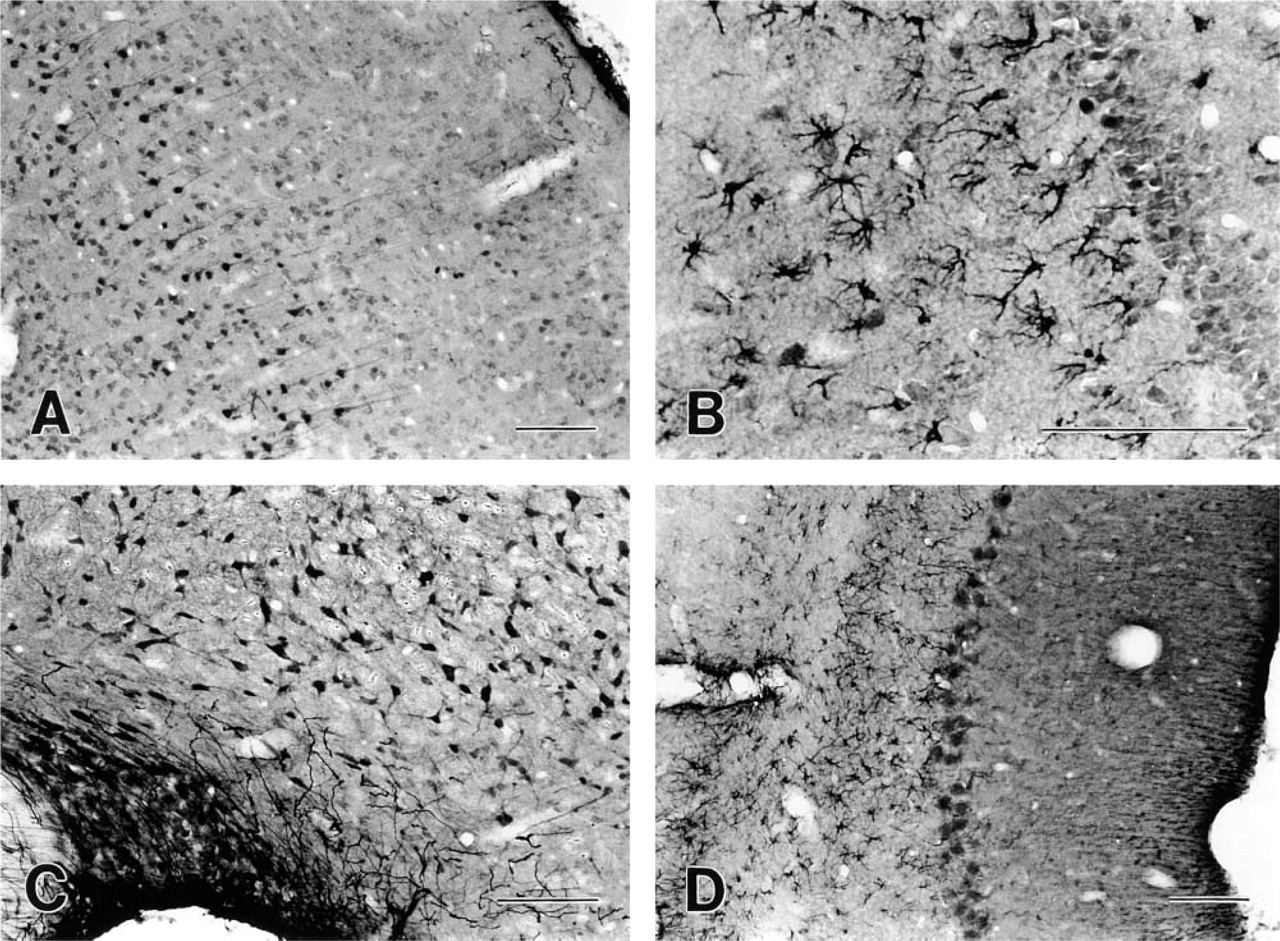
A subset of neurons in the rat brain showed dense immunolabeling with the ANX6 MAb. In general, neuronal immunolabeling was localized to both the nucleus and the cytosol, with some labeling present in the proximal dendrites and axon. ANX6-IR neurons were numerous in cortical areas, particularly in layer 5 of the neocortex (Figure 1A), entorhinal cortex, pre- and post-subiculum, and layers 2-6 of lateral occipital cortex, with the exceptions of the insular and piriform cortices, which were largely unstained. Dark-staining neurons were located in several basal forebrain areas, including the diagonal band, the lateral septum, and the magnocellular preoptic nucleus. In the amygdala and striatum, ANX6-IR neurons were most prominently labeled in the olfactory tubercle, the nucleus of the lateral olfactory tract, and the globus pallidus. In the hypothalamus, many ANX6-IR neurons were found in the anteroventral periventricular nucleus, the lateral anterior hypothalamic nucleus, and the ventrolateral portion of the lateral hypothalamic area (Figure 1C). The reticular thalamic nucleus contained the darkest-staining ANX6-IR neurons in the thalamus. In comparison, the caudate putamen and the dorsomedial hypothalamic nucleus showed a relative lack of immunostaining, whereas many other forebrain areas, such as the ventromedial hypothalamic nucleus and the paraventricular thalamic nucleus, contained lightly labeled neurons. In the rostral brainstem, ANX6-IR was particularly robust in neurons located in the cerebellum (Purkinje cells; Figure 1D); lateral vestibular nucleus, red nucleus, oculomotor nucleus, superior colliculus (intermediate and superficial layers), central gray (dorsal and lateral subdivisions), trapezoid nucleus, and lateral substantia nigra.
ANX6-IR in glial cells was evenly distributed throughout the cell soma and processes. Two types of ANX6-IR glial cells were observed: radial glia and stellate glia. ANX6-IR radial glia were abundant along the brain surface, with many perpendicular processes extending into the optic chiasm, lateral olfactory tract, cerebral peduncle, lateral lemniscus, middle and inferior cerebellar peduncles, spinal tract of the trigeminal nerve, and spinocerebellar and pyramidal tracts. ANX6-IR stellate glia were limited in their distribution to the hippocampus (Figure 1B), cerebellum (Figure 1D), substantia nigra reticulata, internal capsule, and portions of the thalamus. Immunopositive stellate glia displayed a morphology characteristic of astrocytes, with relatively large, rounded cell bodies and long, slender tapering cell processes (Figure 1B).
Our results show that ANX6-IR is expressed in many neuronal cell populations and in a small subset of glial cells in the rat brain. Woolgar et al. (1990) previously detected large amounts (up to 0.3% total protein) of ANX6 in porcine cerebrum and cerebellum. The relative abundance of this peptide in the mammalian nervous system was also noted by Naciff et al. (1996), who observed high concentrations of ANX6-IR in spinal cord motor neurons and sensory ganglion cells.
ANX6-IR was localized in both the cytosol and the nucleus of neurons, but with a nuclear predominance. Previous studies have found that the subcellular distribution of annexin proteins varies by the annexin and, for each annexin, by cell type and with physiological changes in intracellular Ca2+ (Raynal and Pollard 1994; Barwise and Walker 1996; Babiychuk et al. 1999). For example, ANX6 in rat skeletal muscle is associated with both the sarcoplasmic reticulum and the plasma membrane (Luckcuck et al. 1998). In human fibroblasts and smooth muscle cells, ANX6 is diffusely distributed in the cytoplasm but migrates to the plasma membrane in response to chemical stimulation (Barwise and Walker 1996; Babiychuk et al. 1999). Some annexins, such as annexins 1 and 5, show partial co-localization in the cell nucleus, either in the nucleolus or in chromatin-rich areas (Sun et al. 1992; Raynal and Pollard 1994). The subcellular localization of ANX6 in chromatin-rich areas of neuronal nuclei is a novel finding, suggesting a possible role for ANX6 in nuclear function.
Several cytoskeletal proteins present in the nervous system have been found to interact with ANX6 in a Ca2+- or phospholipid-dependent manner (Watanabe et al. 1994; Sheldon and Chen 1996; Garbuglia et al. 1998), suggesting a possible role for ANX6 in Ca2+-dependent regulation of the neural cytoskeleton. For example, S100-B protein has binding sites for both ANX6 and glial fibrillary acidic protein (GFAP), a major component of the astrocyte cytoskeleton (Garbuglia et al. 1998). S100-B inhibits the formation of GFAP intermediate filaments, whereas ANX6 binds to S100-B and blocks its inhibitory effects on GFAP assembly in a dose- and Ca2+-dependent manner (Garbuglia et al. 1998). ANX6 also binds to actin, a major constituent of the neuronal cytoskeleton, and may regulate actin assembly in the axon and nerve terminal (Tanaka et al. 1994; Sheldon and Chen 1996). Several different annexins, including ANX6, interact with spectrin-like peptides, which form a 2D cytoskeletal network underlying the plasma membrane (Watanabe et al. 1994). In the presence of Ca2+, ANX6 dissociates calspectrin (brain spectrin) from F-actin. Calspectrin is necessary for the crosslinking of F-actin microfilaments (Watanabe et al. 1994). Babiychuk et al. (1999) recently provided evidence that ANX6 acts as a linker protein connecting the plasma membrane with the actin-based cytoskeleton in smooth muscle cells. On the basis of these and similar findings, investigators have proposed that annexins act as membrane-cytoskeleton linkage molecules (Sheldon and Chen 1996; Babiychuk et al. 1999). We speculate that annexin 6 may contribute to the Ca2+-dependent membrane-cytoskeletal dynamics that underlie rapid changes in neuronal and glial morphology.
(