
Abstract
HMGN3 (Trip7) is a member of the high-mobility group N (HMGN) nucleosome-binding protein family, which binds specifically to nucleosomes, reduces the compactness of the chromatin fiber, and enhances transcription from chromatin templates. By Western blotting and Northern blotting analysis, we showed that HMGN3 is expressed in a tissue-specific manner, with the strongest expression in mouse brain. Here we analyzed the expression of HMGN3 in various regions of the mouse brain by histological techniques. Enhanced expression of HMGN3 was observed in the lateral olfactory tract, anterior commissure, corpus callosum, internal capsule, fornix, stria medullans, optic tract, and axon bundles. The expression patterns of HMGN3 in the mouse brain was significantly different from that of the related protein HMGN2 and was very similar to that of the glial fibrillary acidic protein (GFAP). We suggest that HMGN3 might play a role in astrocyte function.
T
The HMG proteins are divided into three subgroups: HMGB (formerly HMG-1/-2), HMGA (formerly HMGI/Y/C), and HMGN (formerly HMG-14/-17/Trip 7) (Bustin 2001a). All growing mammalian and most vertebrate cells contain non-histone chromosomal HMGN proteins (Bustin and Reeves 1996; Bustin 1999).
HMGN2 (HMG17) is a well-characterized protein that binds specifically to nucleosomes, reduces the compactness of chromatin fiber, and enhances transcription from chromatin templates (Bustin 2001b). A related protein named HMGN3 (Trip7) was recently detected in a HeLa cell library by a yeast two-hybrid assay through its ligand-dependent interaction with the thyroid hormone receptor TRβ1 (Lee et al. 1995). Both Northern and Western analysis demonstrated that HMGN3 is expressed in a tissue-specific manner in mice (West et al. 2001) and that the highest level of expression was found in whole mouse brain extracts (West et al. 2001). Interestingly, the closely related HMGN2 protein was expressed in most cells and its expression in the brain was not higher than in other tissues (West et al. 2001). On the basis of these observations, the aim of this study was to determine the distribution of HMGN3 proteins in normal mouse brain tissue sections.
Adult mice (20 weeks of age) were sacrificed by decapitation, the brain was removed, immediately immersed in fresh PBS containing 4% paraformaldehyde for 4 hr and 70% EtOH at 4C overnight, and then embedded in paraffin. For immunohistochemical staining, paraffin sections were first blocked with PBST-milk (PBS with 0.1% Tween-20 and 2.5% dry milk) solution and then incubated overnight at room temperature (RT) with either polyclonal rabbit anti-hHMGN3 (dilution 1:100; antibody 2752) (West et al. 2001), or rabbit anti-hHMGN2 (dilution 1:10) (Bustin et al. 1978), or rabbit anti-GFAP (dilution 1:500; Dako; Glostrup, Denmark) antibody solutions diluted in PBST-milk. After washing, the sections were treated with biotinylated horse anti-rabbit IgG (Vectastain ABC kit; Vector Labs, Burlingame, CA) for 1 hr at RT. After washing in the second antibody, the sections were incubated with avidin-biotin-peroxidase complex (Vectastain ABC kit) for 30 min at RT. After a wash with PBS, the bound antibodies were detected with the chromogen 3,3′-diaminobenzidine (DAB) and hydrogen peroxide in 0.1 M PB, as recommended by the manufacturer. For double immunofluorescence, rat anti-GFAP (Zymed Laboratories; San Francisco, CA) and rabbit anti-HMGN3 as primary antibodies, and anti-rat IgG rhodamine conjugate and anti-rabbit IgG fluorescein conjugate (dilution 1:20; CN Biosciences; San Diego, CA) as secondary antibodies were used. Fluorescence staining was observed by a Zeiss Axioplan fluorescence microscopy using the appropriate filters.
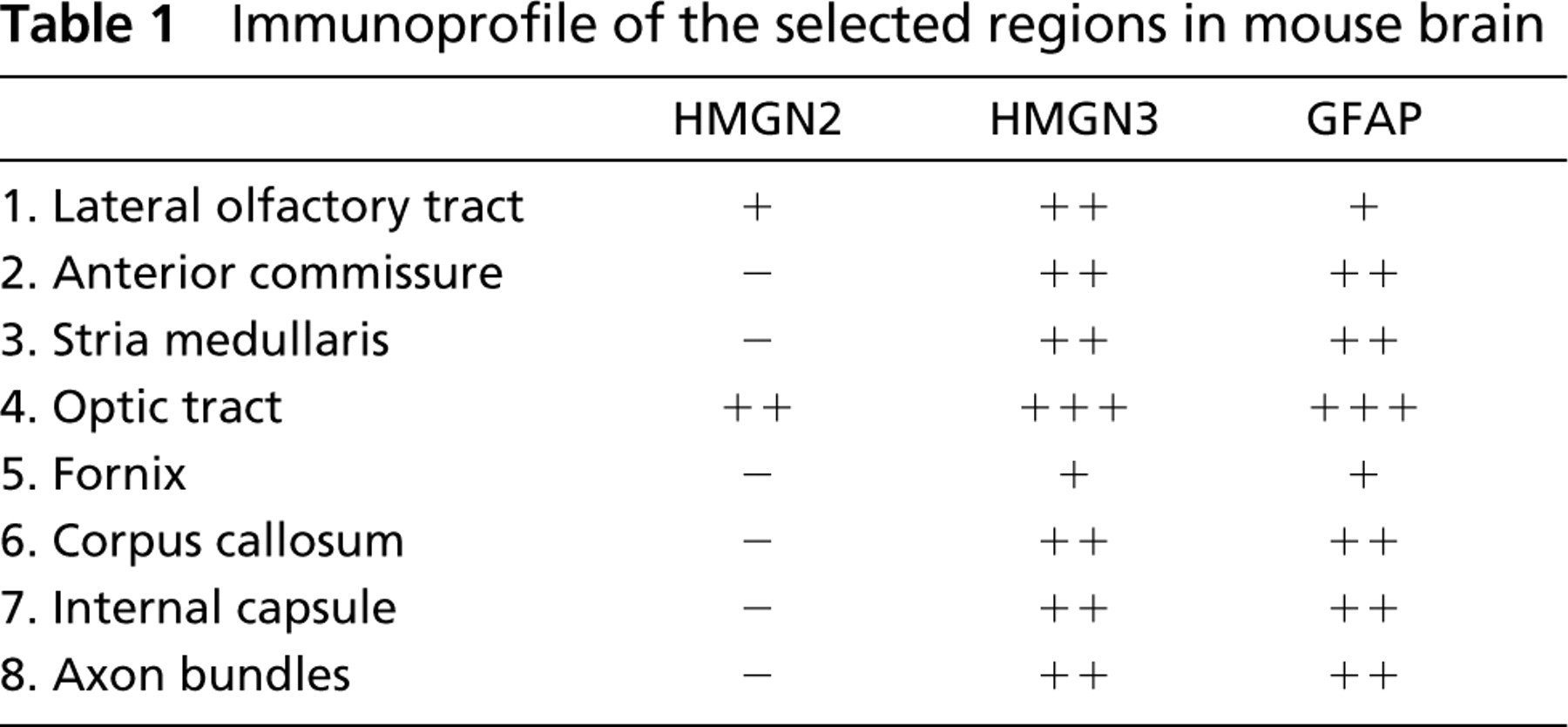
The immunohistochemical data revealed that the expression of HMGN3 was enriched in specific regions of the mouse brain (Figure 1B; Table 1). Relatively high expression levels were detected in fiber bundles including the lateral olfactory tract, anterior commissure, corpus callosum, internal capsule, fornix, stria medullans, optic tract, and axon bundles. Most of these regions are part of the thalamocortical sensory pathway (Alcantara et al. 1992). In contrast, the expression pattern of the closely related HMGN2 proteins did not vary among the various brain regions except that relatively high expression levels were detected in the optic tract (Figure 1A). In addition, HMGN2 was not detected in the fornix, corpus callosum, internal capsule, and in axon bundles.
We found that the expression patterns of HMGN3 in mouse brain are similar to that of GFAP (Figure 1C; Table 1). GFAP (glial fibrillary acidic protein) synthesis is considered an important factor in astrocyte differentiation and is part of the reactive response of the CNS to injury (Eng et al. 2000). In mature astrocytes of the CNS, GFAP is a major component of the 8-9-nm intermediate filaments. GFAP is rapidly synthesized during astrogliosis, the astrocytes' response to various injuries, such of trauma, disease, genetic disorders, or chemical insult, in the CNS of higher vertebrates (Eng et al. 2000). The GFAP gene promoter region has several putative growth factor-binding domains and is actively transcribed in mature brain (Gomes et al. 1999). Among the growth factor domains present in the GFAP promoter are thyroid hormone response elements (Gomes et al. 1999). Indeed, thyroid hormones are known to be important for normal brain development (Thompson and Potter 2000).
Immunoprofile of the selected regions in mouse brain
Thyroid hormones (T3) and thyroid hormone receptors (TRs) also act as chromatin-remodeling factors and are considered to be transcriptional co-activators (Collingwood et al. 1999). Interestingly, HMGN3 protein had been detected by a two-hybrid system as a protein that binds to the TR/retinoid × receptor (RXR) complex in a T3 dependent manner (Lee et al. 1995). It has been suggested that TR recruits HMGN3 to assist in chromatin unfolding and promote transcriptional activation.
To further study the co-localization of HMGN3 with GFAP at the cellular level, we performed double immunofluorescence (Figure 2) with cerebellum, a region in the brain where the distribution of HMGN3 is similar to that of GFAP (Figure 1). As indicated in Figure 2 (middle), expression of HMGN3 was detected in the purkinje cell layer (PC) and was especially prominent in the granular layer (GL). GFAP was also prominently detected in the GL layer (Figure 2, upper). In the molecular layer (MO), the expression of both proteins was not prominent (Figure 2, upper and middle). A merge of the two images (Figure 2, bottom) indicates that the distribution of GFAP (red) partially overlapped (yellow) with that of HMGN3 (green).
These observations, together with our present findings that the localization of chromosomal protein HMGN3 in the mouse brain is similar to that of GFAP, raise the possibility that the distribution of HMGN3 is related to the expression of GFAP. We find that many of the fiber tracts that are related to sensory pathways are strongly immunoreactive for HMGN3. However, we also observed differences in the expression patterns of HMGN3 and GFAP, especially in that the outer Purkinje cells in the PC layer and fibrous cells in the GL layer were prominently stained by HMGN3 (Figure 2). We speculate that HMGN3 might be expressed not only in astrocytes but also in other cells. Therefore, our results raise the possibility that HMGN3 protein plays a functional role in the astrocytes of mouse brain.
Immunohistochemical expression of HMGN2 (
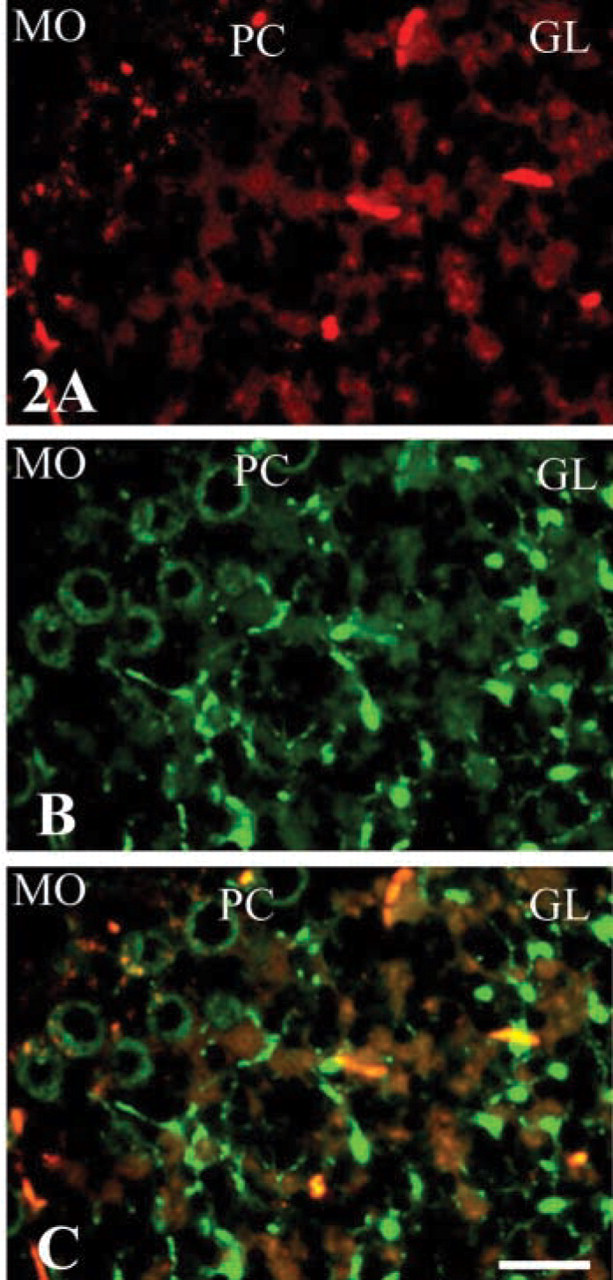
Double immunofluorescence with antibodies against GFAP (