
Abstract
The human tissue kallikrein 13 gene (KLK13), encoding for hK13 protein, was recently cloned and characterized. Here we describe the immunohistochemical (IHC) localization of hK13 in normal human tissues and compare it with the expression of two other kallikreins, hK6 and hK10. We performed the streptavidin-biotin IHC method on 204 paraffin blocks from archival, current, and autopsy material prepared from almost every normal human tissue, using a polyclonal and a monoclonal hK13 antibody. The staining was cytoplasmic and both antibodies yielded similar results. The hK13 protein was revealed in a variety of tissues, mainly in glandular epithelia. Other epithelia that expressed hK13 included the urothelium, the spermatic epithelium, and the epithelium of the choroid plexus. hK13 was intensely immunoexpressed by some endocrine organs, such as the adenohypophysis, the thyroid gland, the parathyroid glands, the adrenal medulla, the Leydig cells of the testis, and the cells of the endocrine pancreas. Immunoreactivity was also observed in the primordial follicles, the corpus luteum, and sparse luteinized cells in the stroma of the ovary, the trophoblastic cells of the placenta, the Hassall's corpuscles of the thymus, and chondrocytes. Nerves and ganglia of the peripheral nervous system, and both neurons and glial cells in the central nervous system, were positive. In short, hK13 was expressed by many glandular epithelia, some endocrine organs, and some specialized epithelia and cells. Comparison of these data with hK6 and hK10 expression suggests that the three kallikreins have a similar IHC pattern in normal human tissues.
Keywords
H
Very few studies have examined human kallikrein expression by IHC in normal or diseased tissues. In our previous reports, we described hK6 and hK10 expression in a variety of normal tissues by IHC (Petraki et al. 2001,2002) and observed that both kallikreins had a similar IHC pattern of expression. Here we describe the IHC localization of hK13 in diverse human tissues and correlate these data with the expression of hK6 and hK10.
Materials and Methods
Representative samples from almost all normal human tissues were studied by IHC. Parts of an organ with different histology (e.g., stomach fundus, body, antrum) were examined separately. Tissues that exist in several organs (e.g., fat, muscle, vessels, peripheral nerves, ganglia) were not studied separately. A total of 204 paraffin blocks were examined. From these, 170 represented archival or current material from 137 cases and the other 34 samples were autopsy material from two cases. The archival and the current material were biopsy or surgical specimens (Table 1). Most but not all of these samples were examined previously for hK6 and hK10 immunoexpression (Petraki et al. 2001,2002).
Tissues studied for the immunohistochemical expression of human kallikrein 13
The IHC staining was performed on 4-μm-thick paraffin sections of tissues fixed in buffered formalin, according to a streptavidin-biotin–peroxidase protocol using the DAKO LSAB+Kit Peroxidase (DAKO; Glostrup, Denmark). An hK13-specific rabbit polyclonal antibody (1:500) and two hK13 (2–17 and 11C1) mouse monoclonal antibodies (MAbs: 1:150 and 1:50, respectively) raised against full-length recombinant hK13 protein produced in yeast were used. The recombinant hK13 protein was produced and purified by HPLC as previously described for hK10 protein (Luo et al. 2001b). None of these antibodies crossreacted with other kallikreins in ELISA assays (Kapadia et al. in press). Staining procedures included deparaffinization in warm xylene for 5 min with two changes of xylene at room temperature (RT), followed by rehydration by transfer through graded alcohols. Endogenous peroxidase activity was blocked with 0.5% H2O2 in methanol for 10 min. The sections were pretreated with 10 mmol/liter citrate buffer (pH 6.1) in a microwave oven for 5 min and were incubated overnight at 4C with the hK13 primary rabbit polyclonal (PAb) and mouse MAb in 3% BSA. After two washes of the sections in 50 mM Tris buffer (pH 7.6), the biotinylated link (DAKO; Carpinteria, CA) was applied for 15 min and a streptavidin-peroxidase conjugate followed for another 15 min. The enzymatic reaction was developed in a freshly prepared solution of 3,3′-diaminobenzidine tetrahydrochloride using DAKO Liquid DAB Substrate-Chromogen Solution for 10 min (brown color). The sections were then counter-stained with hemalum, dehydrated, cleared in xylene, and mounted. The staining pattern, the distribution of the immunostaining in each tissue, and the intensity of the staining were studied in detail.
In selected tissues the primary polyclonal antibody was replaced by a nonimmune rabbit serum (1:500) in 3% BSA to evaluate nonspecific binding. For the same reason, an immunoabsorption test was also performed in which the primary hK13 antibody was incubated for 1 hr at RT with an excess of recombinant hK13 antigen.
Results
The hK13 immunoreactivity, using the polyclonal antibody and the two MAbs, was generally localized in the cytoplasm. All antibodies revealed the same distribution of the antigen in all tissues. Replacement of the primary polyclonal antibody with nonimmune rabbit serum and the immunoabsorption test resulted in disappearance of the immunostaining in all tissues examined.
In our previous studies we did not find any major differences between the immunolocalization of hK6 and hK10 in normal human tissues. It is worth mentioning that, in the present study, hK13 had, in general, a similar IHC distribution as the other two kallikreins. The protein was revealed in a variety of tissues, indicating that it is not tissue-specific.
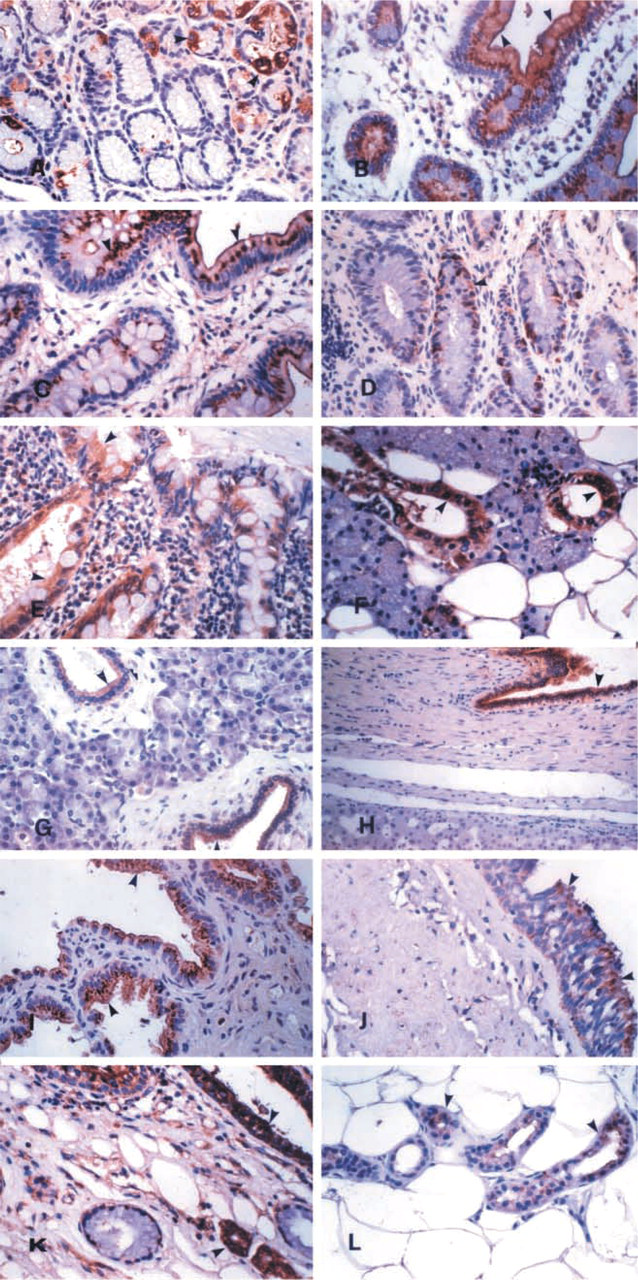
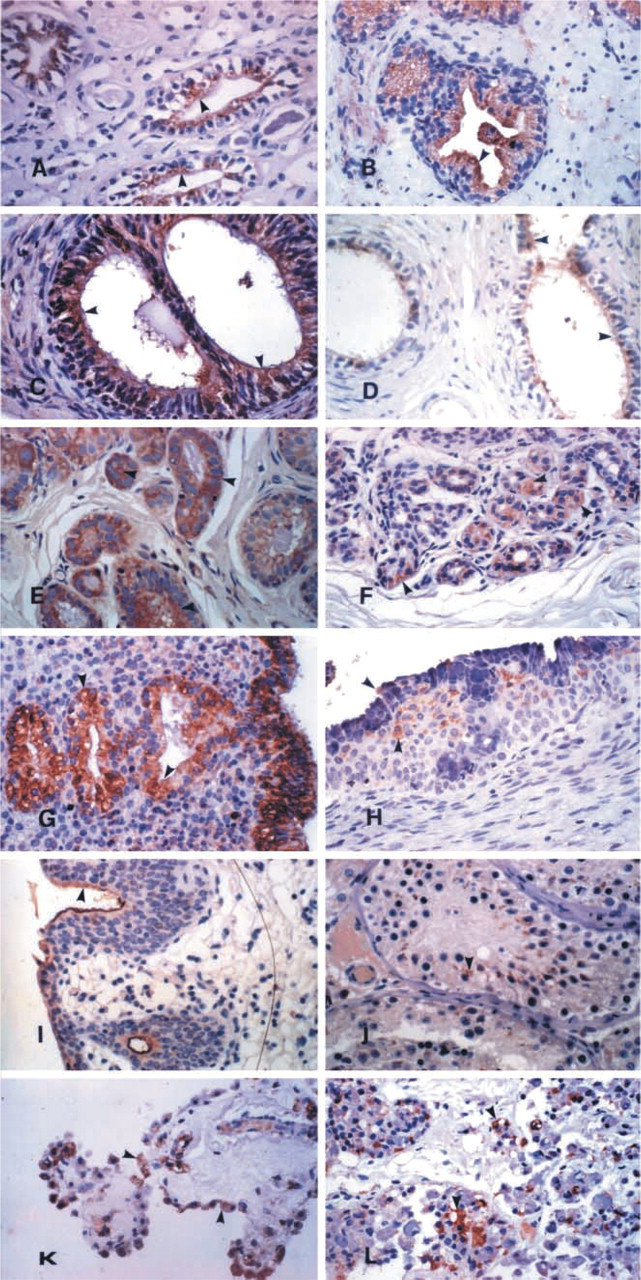
The hK13 protein was immunoexpressed mainly by glandular epithelia. The gastric mucosa expressed the kallikrein focally in all cell types (Figure 1A). A characteristic supranuclear, droplet-like, and brush border localization was observed in the duodenum (Figure 1B) and the other parts of the small intestine (Figure 1C). All parts of the large intestine (Figure 1D) and the appendix (Figure 1E) showed strong and diffuse immunoexpression in both the enterocytes and the goblet cells. The major and minor salivary glands (Figure 1F), as well as the submucosal glands of the pharynx and the esophagus, expressed the antigen. The epithelium of the pancreatic (Figure 1G) and bile duct system (Figure 1H), and also of the gallbladder (Figure 1I), was positive. Strong cytoplasmic staining was observed in the epithelium of the bronchial tree (Figure 1J). All glands of the upper and lower respiratory system (nasal, sinusoidal, laryngeal, tracheal, and bronchial) were strongly positive (Figure 1K). The epithelium of the eccrine and apocrine glands of the skin was also strongly positive (Figure 1L). The glandular epithelia in the genitourinary tract and in the male and female reproductive systems expressed hK13. The urinary tubuli of the kidney expressed the protein diffusely (Figure 2A). Immunoexpression in the epithelium of the prostate (Figure 2B), the ejaculatory ducts, the seminal vesicles, the rete testis, the epididymis (Figure 2C), and the spermatic duct (Figure 2D) was diffuse and strong. The staining in the surface epithelium of the ovary, epithelium of the fallopian tubes, breast (Figures 2E and 2F), and endometrium (Figure 2G) was strong, but in some cases focal. No differences were observed during the course of the menstrual cycle. The columnar epithelium of the endocervix showed weaker and more focal distribution, and foci of immature squamous metaplasia also expressed the antigen (Figure 2H).
(
(
Weak and diffuse immunoexpression of hK13 by the squamous epithelia in the different organs of the human body was not considered as positive. Other than glandular epithelia, epithelia that expressed hK13 were the urothelium, with immunolocalization mainly in the superficial cells (Figure 2I), the spermatic epithelium (Figure 2J), and the epithelium of the choroid plexus (Figure 2K).
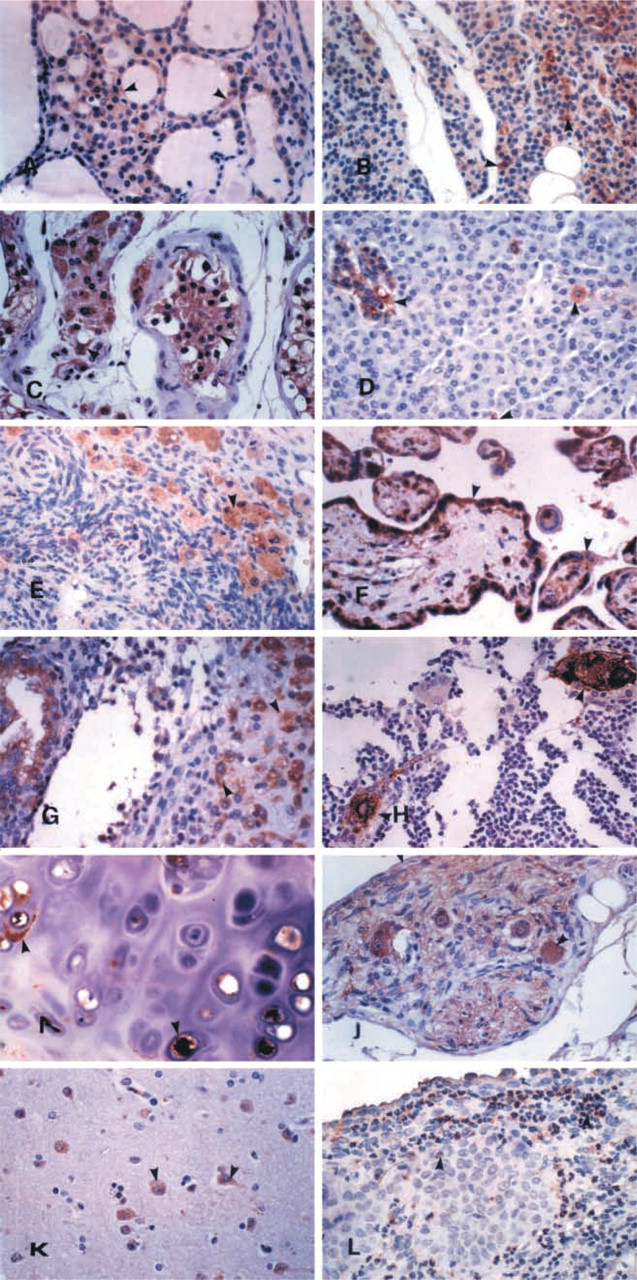
The hK13 protein was intensely immunoexpressed by some endocrine organs and showed a similar distribution to that of the other two kallikreins. The lactotrophs and the corticotrophs of the anterior lobe of the pituitary (Figure 2L), the epithelium of the thyroid follicles, mainly in foci of hyperplasia oxyphilic metaplasia (Figure 3A), the cells, mainly oxyphilic, in the parathyroid glands (Figure 3B), the adrenal medulla, the Leydig cells of the testis (Figure 3C), and the cells of the endocrine pancreas islets of Langerhans and scattered endocrine cells in the exocrine pancreas (Figure 3D) also expressed hK13.
(
A clear immunoreaction was also observed in the primordial follicles, the corpus luteum, and in sparse luteinized cells in the stroma of the ovary (Figure 3E). The trophoblastic cells of the placenta expressed this kallikrein in all phases of their development (Figure 3F). The decidual cells in the endometrium during pregnancy and in certain endocrine disorders were also positive (Figure 3G).
The Hassall's corpuscles of the thymus (Figure 3H) and the chondrocytes (Figure 3I) also expressed the kallikrein. Nerves and ganglia of the peripheral nervous system (Figure 3J) were strongly positive. In the central nervous system, both neurons and glial cells were positive (Figure 3K). Among the inflammatory cells, neutrophils expressed hK13 strongly (Figure 3L).
In short, hK13 was expressed by many glandular epithelia, by some endocrine organs, and by some specialized epithelia and cells.
Discussion
The human kallikrein 13 gene (KLK13) was cloned by using the positional candidate approach (Yousef et al. 2000). By employing reverse transcription-polymerase chain reaction analysis, it was reported that KLK13 is expressed in various tissues, including breast, prostate, salivary glands, testis, lung, heart, thymus, adrenal, colon, thyroid, and trachea (Diamandis et al. 2000b; Yousef and Diamandis 2001; Yousef et al. 2000). In addition, using ELISA-based methodologies for hK13, we confirmed that this protein is present in tissue extracts from esophagus, tonsil, salivary glands, prostate, kidney, skin, trachea, ureter, testis, breast, lung, thyroid, and stomach (our unpublished data). Furthermore, hK13 protein was found at relatively high concentrations in biological fluids such as seminal plasma, amniotic fluid, milk of lactating women, follicular fluid, and cerebrospinal fluid (our unpublished data). This combined information is in accord with the IHC data presented here. It is clear that the hK13 protein is expressed in many human tissues and cannot be considered a tissue-specific marker. Among all the kallikreins, some have tissue-restricted specificity, including hK2 and hK3, which are highly expressed in the prostate and at much lower levels in other tissues. In general, our data presented here correlate well with the expression of human kallikreins 6 and 10, two kallikreins that have been immunohistochemically localized by using similar techniques (Petraki et al. 2001,2002).
We report here that hK13 is expressed mainly by glandular epithelia, further suggesting that this protein is secreted. The finding of hK13 protein in seminal plasma, amniotic fluid, milk of lactating women, follicular fluid, and cerebrospinal fluid further supports this hypothesis. Among other kallikreins that have been identified, all are found in biological fluids, an indication that all members of this family are secreted serine proteases.
In the testis, we have demonstrated that Leydig cells and the spermatic epithelium express hK13 protein. In our previous work we described the identification of five new mRNA KLK13 splice variants, which are expressed only in human testis (Chang et al. 2001). We have also reported differential expression of the KLK13 gene in testicular cancer. Furthermore, the five splice variants appear to be downregulated or not expressed at all in testicular cancer. The physiological and pathobiological roles of hK13 protein in the testis warrant further investigation.
The hK13 protein is expressed at high levels in the pituitary, especially in lactotrophs and corticotrophs, and in epithelium-lined spaces of the intermediate lobe remnant, which are prolactin-immunoreactive. The same distribution was observed with hK6 and hK10 expression in our previous studies (Petraki et al. 2001,2002). It is worth mentioning that another kallikrein, KLK1, has been found in the lactotrophs of the rat and in the human anterior pituitary (Jones et al. 1990; Vio et al. 1990). Furthermore, we found hK13 along with hK6 and hK10 in the islet cells of the human pancreas. Moreover, these kallikreins were expressed in the epithelium of the pancreatic ducts but not in the acinar cells of the exocrine pancreas. Pancreatic kallikrein (KLK1) has also been found in pancreatic cells, including the β-cells of the pancreatic islets (Ole-MoiYoi et al. 1979; Pinkus et al. 1983). We have previously speculated that the presence of these proteolytic enzymes in the pituitary and the islet cells of pancreas may be related to their ability to degrade hormone precursors, an activity that has been attributed to many other proteolytic enzymes (Mason et al. 1983; Coughlin 1999). The expression of human kallikrein 13 in the salivary glands is expected, because all other identified kallikreins are also expressed in this tissue (Diamandis et al. 2000b; Yousef and Diamandis 2001).
The expression of a few kallikreins in the CNS has been previously reported (Diamandis et al. 2000b; Yousef and Diamandis 2001). Yamanaka et al. (1999) found KLK6 mRNA in mature oligodendrocytes and suggested that this protease may be important in the processes that occur after maturation of oligodendrocytes, such as myelination or turnover of the proteins in myelin. Furthermore, Little et al. (1997) detected positive hK6 immunostaining in monkey cortex cells lining the perimeter of cortical microvessels, in brains of human patients with Alzheimer's disease, and in microglial cells, indicating a role of this protease in brain disease. Furthermore, new knowledge about human kallikrein 6 suggests a role in neurodegenerative diseases (Diamandis et al. 2000c; Ogawa et al. 2000; Okui et al. 2001; Bernett et al. 2002; Blaber et al. 2002; Gomis-Ruth et al. 2002). It will be useful to examine the involvement of hK13 and other kallikreins in human neurodegeneration.
We have previously reported expression of hK6 and hK10 in oxyphylic cells in the thyroid and parathyroid glands. The expression of hK13 in these tissues is similar. Previously, the presence of hK2 and hK3 in the thyroid has been reported (Magklara et al. 2000).
Many kallikreins have found important applications as cancer biomarkers, including hK2 and hK3 for prostate cancer (McCormack et al. 1995; Chu 1997; Rittenhouse et al. 1998), hK6 and hK10 for ovarian carcinoma (Anisowicz et al. 1996; Diamandis et al. 2000d; Luo et al. 2001a), and hK11 for ovarian and prostate carcinoma (Diamandis et al. 2002). The availability of a sensitive method for measuring hK13 in biological fluids will allow examination of this protein as a disease biomarker. Moreover, the expression of multiple kallikreins in normal and cancerous tissues has prompted us to speculate that multiple members of this gene family may participate in cascade enzymatic reactions, similarly to the coagulation and apoptotic cascades. The localization of the three aforementioned kallikreins (hK6, hK10, and hK13) in the same types of human cells, as well as their hormonal regulation (Diamandis et al. 2000b; Yousef and Diamandis 2001), further supports this hypothesis.
In summary, we describe for the first time the IHC localization of hK13 in various human tissues. Further studies will be necessary to examine the physiological and pathobiological role of this protein in these tissues.