
Abstract
The KLK6 gene is a new member of the human kallikrein gene family and encodes for a secreted protease, human kallikrein 6 (hK6; also known as zyme/protease M/neurosin). No study has as yet reported detailed immunohistochemical localization of hK6 in human tissues. Our purpose was to examine the expression of hK6 in human tissues by immunohistochemistry. We have analyzed 199 paraffin blocks from archival, current, and autopsy material prepared from almost every normal human tissue. We employed an hK6-specific polyclonal rabbit antibody and avidin-biotin to localize hK6 by IHC. The staining pattern, the distribution of the immunostaining, and its intensity were studied in detail. The IHC expression of zyme was generally cytoplasmic. Various normal human tissues expressed the protein abundantly. Glandular epithelia constituted the main immunoexpression sites, with representative organs being the breast, prostate, kidney, endometrium, colon, appendix, salivary glands, bile ducts, and gallbladder. The small intestine, stomach, endocervix, Fallopian tube, epididymis, bronchus, and upper respiratory tract showed a focal expression as well. Choroid plexus epithelium, peripheral nerves, and some neuroendocrine cells (including the islets of Langerhans, cells in the anterior pituitary gland, and adrenal medulla) expressed the protein strongly and diffusely. A characteristic immunostaining was observed in the Hassall's corpuscles of the thymus, the oxyphilic cells of the thyroid and parathyroid glands, the primordial follicles of the ovary, dendritic cells mainly in the spleen, and in various cells of the placenta.
Keywords
U
New members of the human kallikrein gene family have recently been discovered (Diamandis et al. 2000a). This gene family now consists of 15 genes which all encode for trypsin-like or chymotrypsin-like serine proteases, show significant homology at both the DNA and amino acid level, and are localized at the chromosomal locus 19q13.4. This area of investigation has recently been reviewed (Diamandis et al. 2000a; Yousef and Diamandis 2001).
The KLK6 gene (encoding for human kallikrein 6, hK6) has been cloned independently by three groups of investigators and was previously given the names zyme (Little et al. 1997) (cloned from brain tissue), protease M (Anisowicz et al. 1996) (cloned from breast tissue), and neurosin (Yamashiro et al. 1997) (cloned from a colon carcinoma cell line). Recently, uniform nomenclature for all newly discovered and traditional kallikrein genes has been established (Diamandis et al. 2000b). The KLK6 gene encodes for a trypsin-like serine protease of 244 amino acids, of which 16 amino acids constitute the signal peptide and five amino acids the activation peptide. The mature enzyme consists of 223 amino acids. It has been previously predicted that hK6 is a secreted protein (Anisowicz et al. 1996; Little et al. 1997; Yamashiro et al. 1997; Yousef et al. 1999). This was recently verified by the finding of hK6 protein in various biological fluids, including cerebrospinal fluid, breast milk, nipple aspirate fluid, breast cyst fluid, male and female serum, seminal plasma, amniotic fluid and breast cancer cytosols (Diamandis et al. 2000c). Little et al. (1997) suggested that this enzyme has amyloidogenic potential in the brain and may play a role in the development and progression of Alzheimer's disease. Anisowicz et al. (1996) have cloned the same gene by the method of differential display, and found that it is downregulated in aggressive forms of breast cancer. The same gene was cloned by Yamashiro et al. (1997) from the human colon adenocarcinoma cell line COLO 201.
Recently, we have established the genomic organization and the hormonal regulation of the KLK6 gene and studied its tissue expression by rRT-PCR (Yousef et al. 1999). We found that this gene is abundantly expressed in many tissues, including the salivary gland, brain, uterus, heart, thymus, prostate, liver, breast, thyroid, spleen, placenta, trachea, testis, and kidney (Yousef et al. 1999). We have further raised polyclonal mouse and rabbit antibodies and developed an immunofluorometric procedure for measuring hK6 with high sensitivity and specificity (Diamandis et al. 2000c). Using this assay, we have recently found that hK6 is elevated in serum of ovarian cancer patients and in serum and cerebrospinal fluid of patients with Alzheimer's disease (Diamandis et al. 2000d; Diamandis et al. 2000e). However, no study has as yet described the immunolocalization of hK6 in human tissues. Here we describe immunohistochemical localization of hK6 in diverse human tissues.
Materials and Methods
This was an IHC study on almost all normal human tissues to establish the expression of hK6. Parts of an organ with different histology (e.g., stomach, fundus, body, antrum) were examined separately. A paraffin block of three different cases for every tissue (organ, all parts with different histology) was selected. Cases with malignancy in adjacent sites of the organ were excluded to avoid phenotypic changes that may be associated with cancer. Tissues that exist in several organs (e.g., fat, muscle, vessels, peripheral nerves, ganglia, and neuroendocrine cells) were not studied separately. A total of 199 paraffin blocks were examined. A total of 165 blocks were from archival or current material from 132 cases and the rest were autopsy material from two cases (Table 1).
The avidin-biotin complex IHC method was performed on 4-μm-thick sections using two hK6-specific antibodies, one rabbit polyclonal and one mouse monoclonal. Both antibodies were raised in-house against full-length hK6 produced recombinantly in a mammalian stable cell line system (Little et al. 1997). The recombinant hK6 protein was purified by HPLC as previously described (Little et al. 1997). The specificity of the antibodies was evaluated during development of the immunofluorometric assay, as previously described, and by Western blotting analysis (Diamandis et al. 2000c; and our unpublished data). We found no detectable crossreactivity from other closely related antigens such as hK2, hK3, and hK10 (Diamandis et al. 2000c). All tissues were fixed in buffered formalin. Staining procedures included deparaffinization in xylene and then two changes of xylene at room temperature, followed by rehydration by transfer through graded alcohols. Endogenous peroxidase activity was blocked with 0.5% H2O2 in methanol for 10 min. The sections were pretreated with citrate buffer (pH 6.1) in a microwave for 5 min and incubated overnight with the hK6 primary rabbit polyclonal antibody (1:500) and the hK6 primary mouse monoclonal antibody (1:150) in 3% BSA. After washing with Tris-buffered saline (TBS, pH 7.6) for 15 min, they were incubated with a biotinylated secondary antibody for 10 min. After washing with TBS, we incubated with the avidin-peroxidase complex for 10 min. The next steps included application of diaminobenzidine (DAB) substrate and incubation for 10 min, washing in running water, staining with hematoxylin for 1 min, washing in running water again, dehydration through graded alcohols, and covering by DPX. A negative control section was always included in which the primary antibody was substituted with non-immune rabbit or mouse serum. In addition, for selected tissues we performed immunoabsorbtion of the primary hK6 antibody by mixing it for 1 hr with excess recombinant hK6 before immunostaining.
The staining pattern, the distribution of the immunostaining in each tissue, and the intensity of the staining were studied in detail.
Results
The hK6 immunoreactivity using the polyclonal and the monoclonal antibody was generally localized in the cytoplasm. Both antibodies revealed the same distribution of the antigen in all tissues. Replacement of the primary antibody with non-immune serum or immunoabsorbtion of the primary antibody abolished the immunostaining in all tissues examined. These findings indicate that our staining procedure is specific for hK6. The protein was revealed in a variety of tissues, indicating that this is not a tissue-specific protein. hK6 is mainly expressed by glandular tissues, but our data suggested that it could also serve as a neuroendocrine marker. The distribution and the expression levels of hK6 in various tissues are described below and further summarized in Table 2.
Tissues studied for the immunohistochemical expression of human kallikrein 6
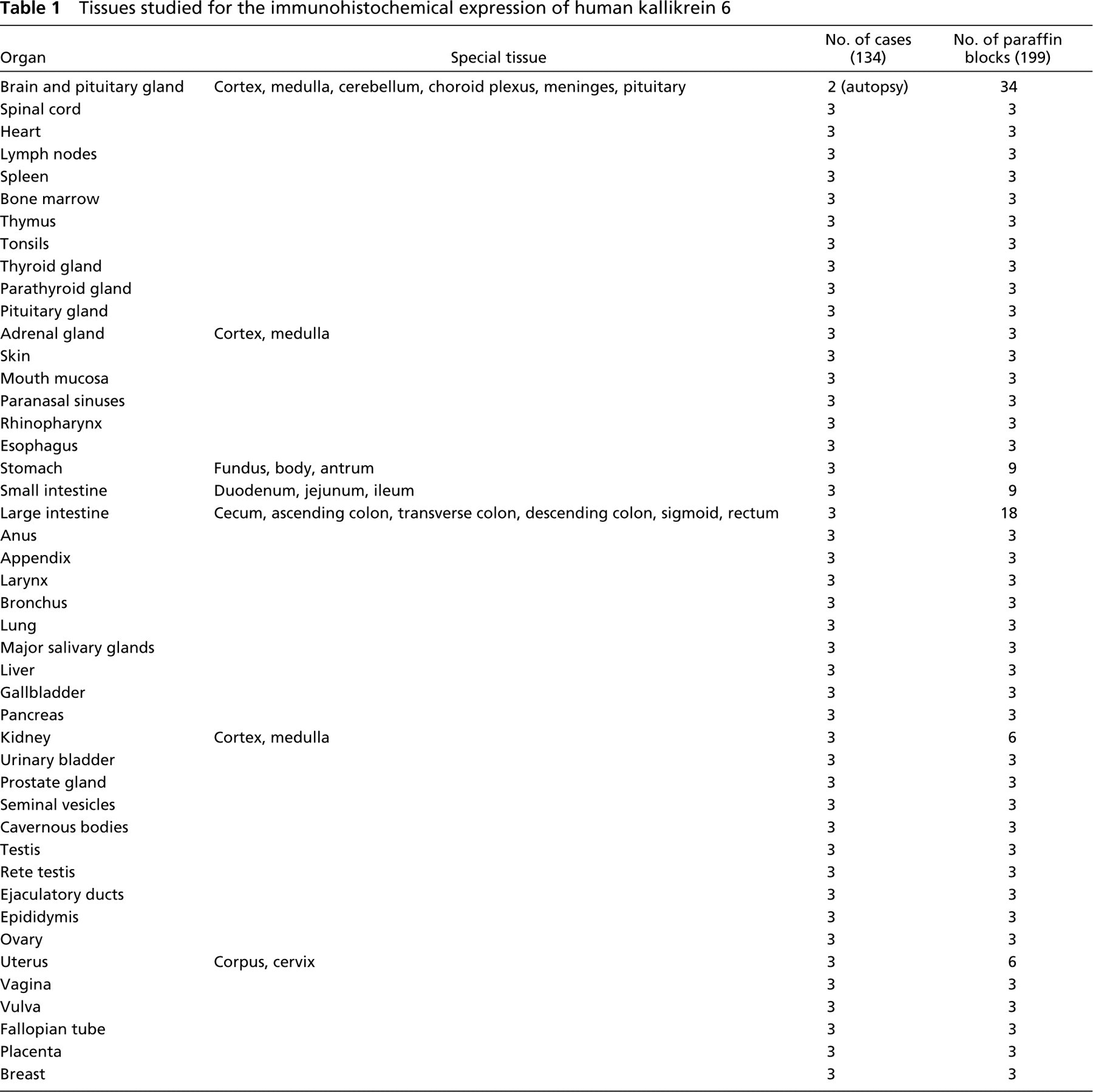
Central and Peripheral Nervous System
Strong and diffuse positivity in the epithelium of the choroid plexus was observed. (Figures 1A and 1B). In the cerebellum, the antigen was expressed weakly in the Purkinje cells and the stellate (basket cells), whereas the granular cells were negative. In the entire central nervous system the nerve cells showed weak immuno-staining as well (Figure 1C). Glial cells showed a weaker expression focally. Staining of the peripheral nerves was intense (Figure 1D).
Female Reproductive System
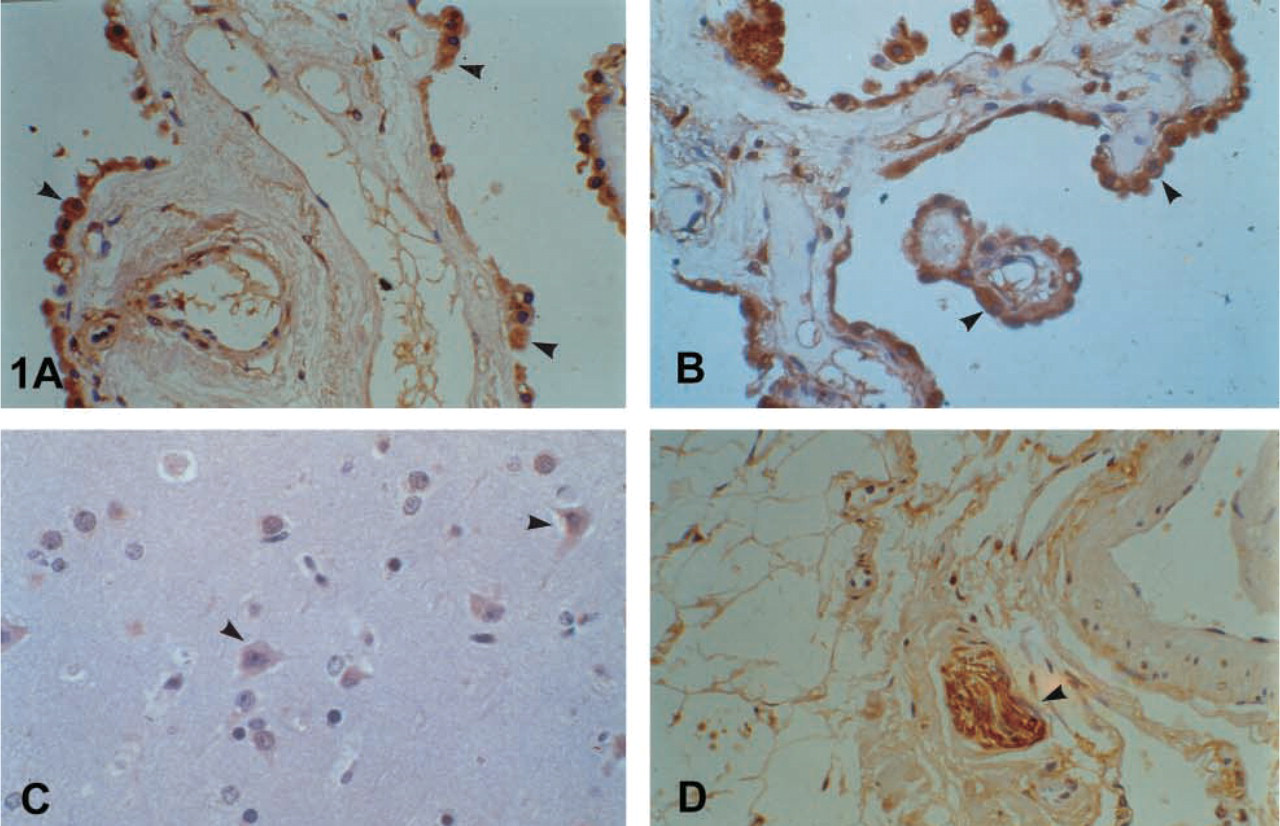
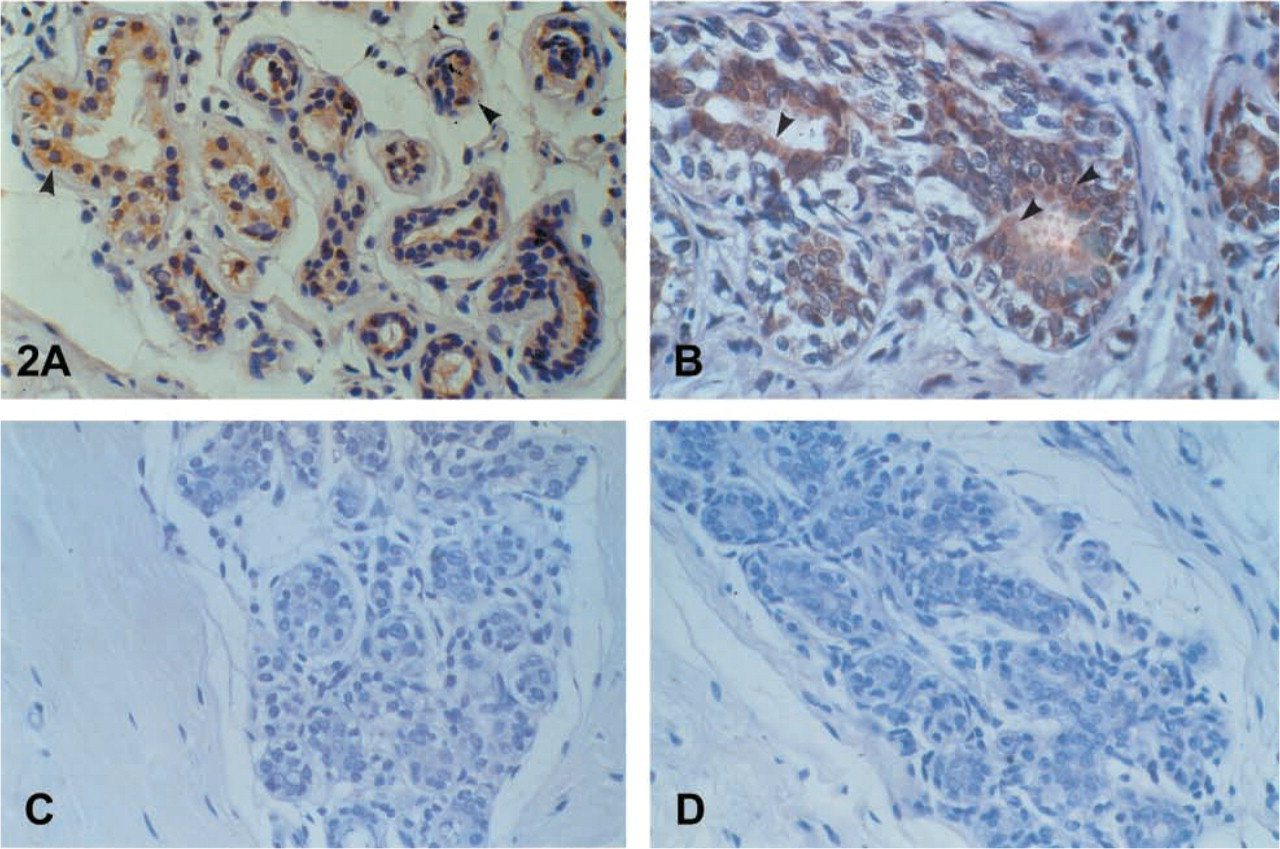
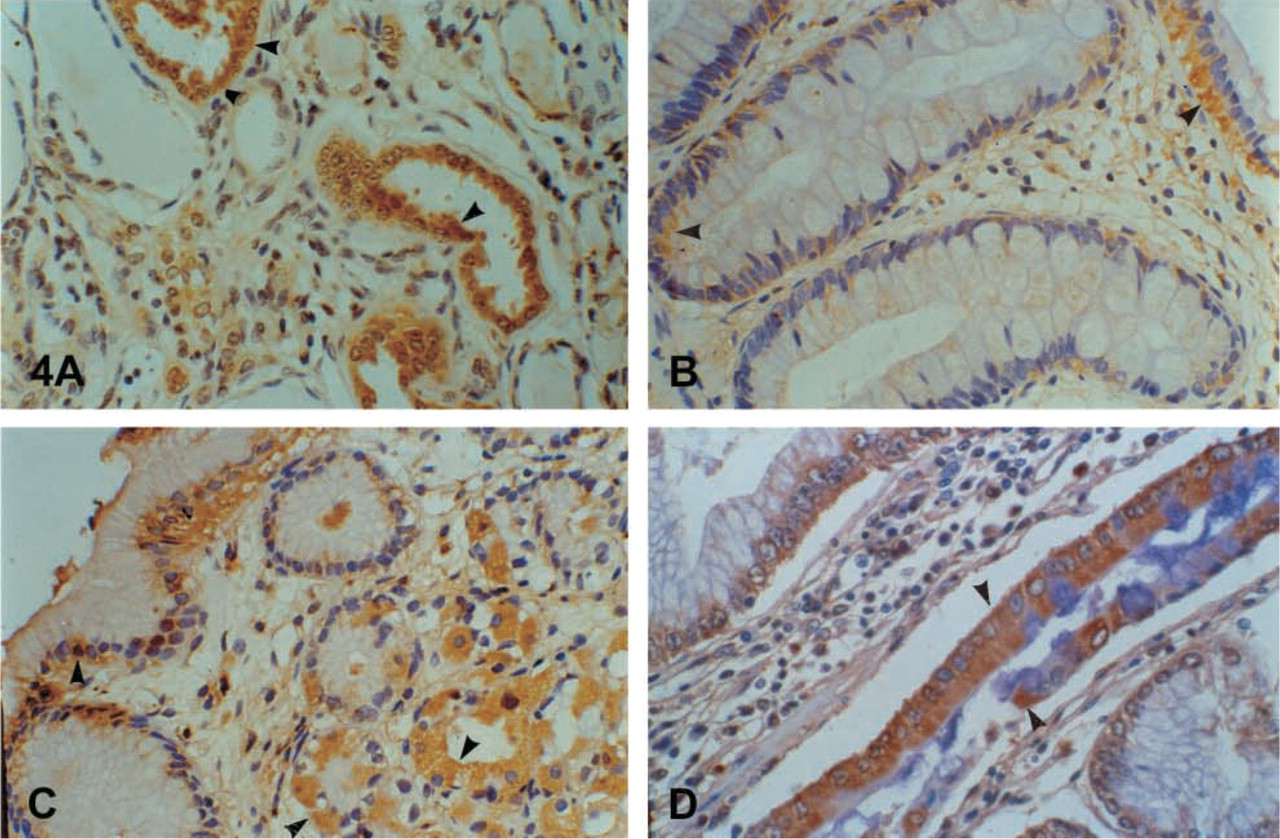
The epithelium of the breast stained positive. Cytoplasmic and brush-border distribution was observed, and luminal secretions were positive as well (Figures 2A-2D). A similar immunophenotype appeared in the columnar epithelium of the endometrium and the endocervix of the uterus. A characteristic droplet-like, widely distributed expression was noted in the endometrium (Figure 3A), but staining in the endocervix was focal and paranuclear. No major differences were seen during the menstrual cycle. The myometrium was negative. Weak and focal expression by the squamous cervical epithelium could not be safely considered as positive (see Discussion). The squamous epithelium of the vagina was negative. Supranuclear cytoplasmic, brush-border, and ciliary staining was revealed in the Fallopian tubes. Characteristic was the positivity in the primordial follicles of the ovary. In the placenta the protein was localized in the endothelia, in calcifications of the villi, as well as in “X” cells and, focally, in trophoblastic cells.
Tissues with a hK6 immunoexpression

D, diffuse; C, cytoplasmic; BB, brush border; LS, luminal secretions; PN, paranuclear; SpN, supranuclear; SbN, subnuclear.
Genitourinary Tract
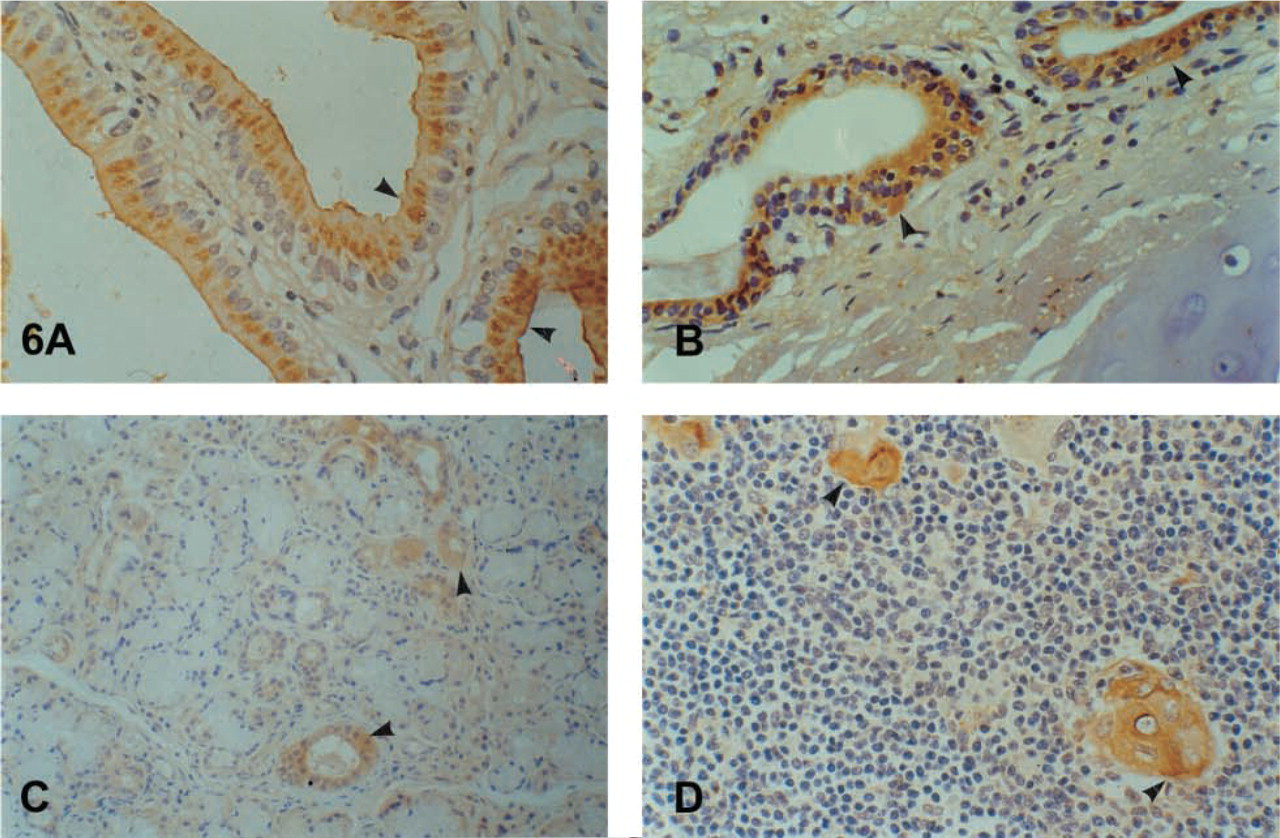
Strong diffuse immunostaining was observed in the prostate columnar cells. Basal cells remained unstained. Because we did not use a double-staining method, it was not possible to evaluate the positivity of the neuroendocrine cells. In general, hK6 had the same immunostaining pattern in the benign prostate gland as hK2 and hK3 (Figures 3B-3D). A focal, mainly suprabasal cytoplasmic, brush-border and ciliary staining in the epithelium of the epididymis, seminal vesicles, and spermatic duct was the rule. The urinary tubuli of the kidney (Figure 4A) showed immunoexpression as well. It is doubtful if weak focal expression by the umbrella cells of the urothelium and the Leydig cells of the testis could be considered as positive. Cavernous bodies were negative.
Gastrointestinal Tract
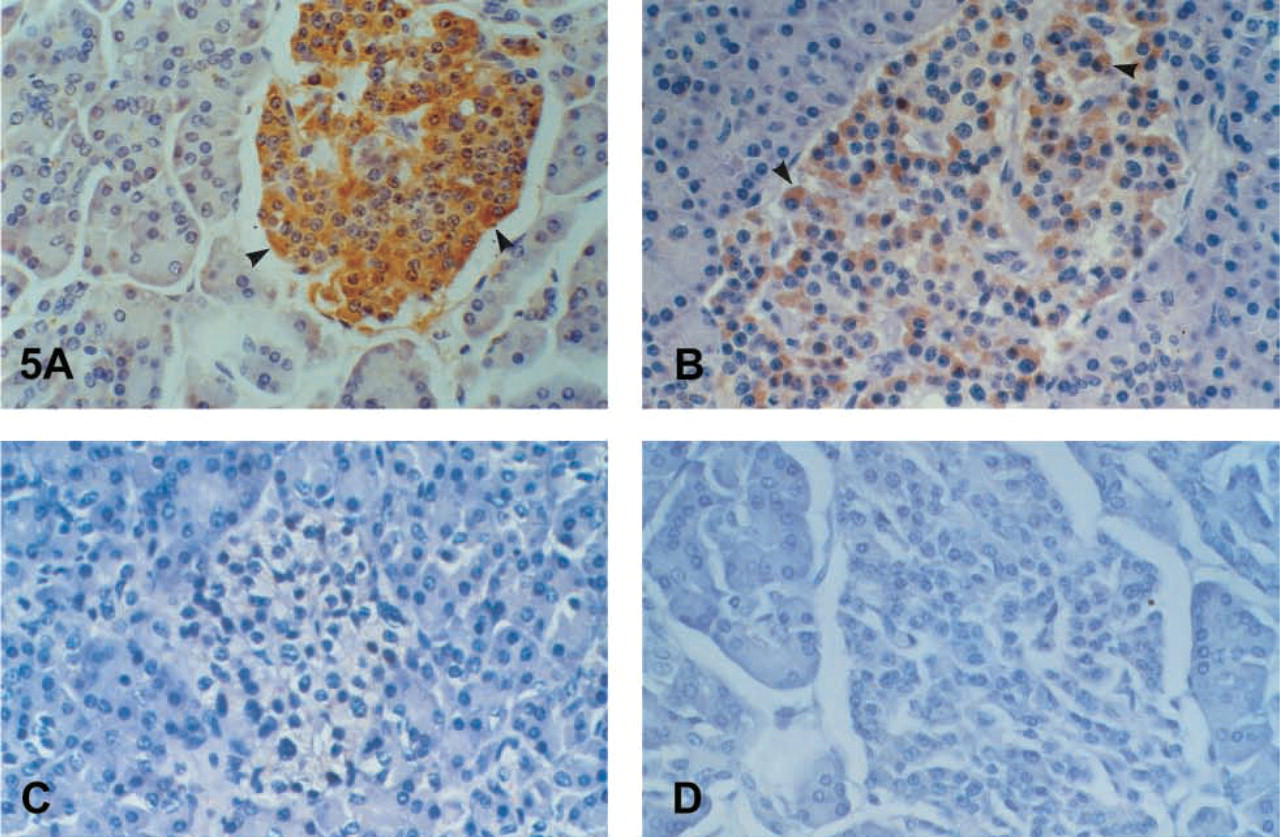
The glandular epithelium of the large bowel and the appendix showed strong, diffuse cytoplasmic, mainly subnuclear (large intestine) and supranuclear (appendix) immunostaining in the enterocytes (Figure 4B). No clear differences were noted among the different segments of the colon. Expression in the small intestine was focal cytoplasmic, mainly subnuclear in the duodenum and mainly peri- and supranuclear in the ileum. The antrum of the stomach showed a focal cytoplasmic, mainly subnuclear and brush-border staining; the body showed a brush-border and parietal cell expression (Figure 4C). Strong positivity in foci of intestinal metaplasia in the gastric mucosa was the rule (Figure 4D). The duct epithelium of the eosophageal glands expressed the antigen as well. Expression by neuroendocrine cells throughout the gastrointestinal tract was generally obvious. The reactivity by basal cells in the epithelium of the esophagus and the anus was considered to arise from neuroendocrine cells. We found strong positivity in the cells of the islets of Langerhans in the pancreas (Figures 5A-5D). The acinar cells of the exocrine pancreas were negative. Only some scattered positive cells were observed between them. The epithelium of the medium-sized pancreatic ducts showed a cytoplasmic and mainly brush-border immunostaining. Hepatocytes were negative. A cytoplasmic and brush border immunostaining was observed in the bile ducts and the gallbladder mucosa (Figure 6A).
(
(
Respiratory Tract
A mainly brush-border staining in the respiratory epithelium of the bronchus, larynx, trachea, rhinopharynx, and paranasal sinuses was observed (Figure 6B). The staining in the ducts was cytoplasmic. The alveoli of the lung were negative.
Salivary Glands (Major and Minor) and Skin Appendages
Cytoplasmic positivity in the ductal epithelium and scattered positive cells in the alveoli were noted (Figure 6C).
Spleen, Tonsils, Lymph Nodes, and Bone Marrow
Some positive cells, possibly dendritic, in the germinal centers of the follicles, mainly in the spleen, were found. Various inflammatory cells, mainly neutrophils but also some plasma cells, also showed positivity.
Thymus
Strong positivity in the Hassall's corpuscles was characteristic (Figure 6D).
Adrenal Gland
Weak to moderate positivity was observed in the medulla.
Thyroid Gland
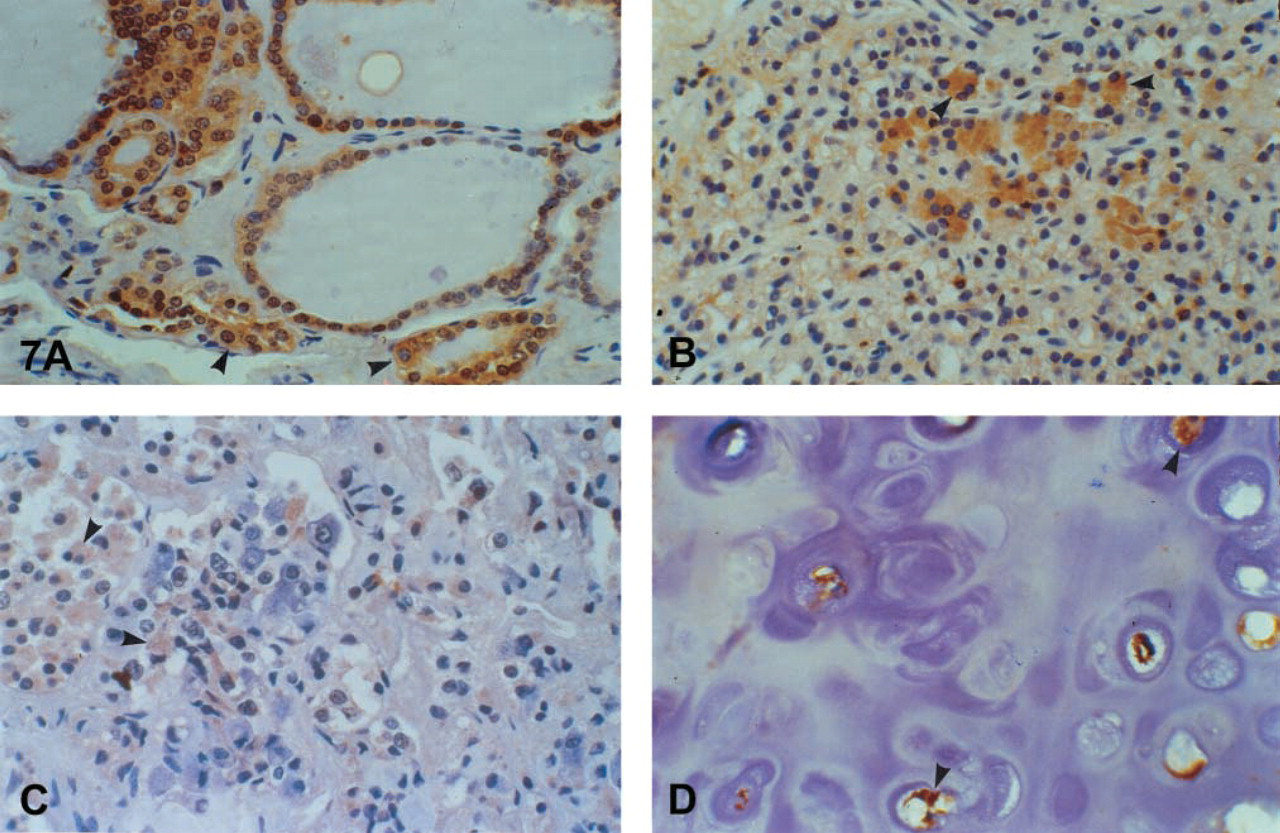
Focal protein immunoexpression was revealed by follicular cells, mainly in hyperplastic conditions and in oxyphilic cell metaplasia (Figure 7A).
Parathyroids
Immunoexpression by the oxyphilic cells was noted (Figure 7B).
Pituitary Gland
In the anterior pituitary, many cells expressed the protein strongly (Figure 7C). The pituitocytes of the pars nervosa were negative.
Mesothelium (Pleura)
The positivity was variable and was observed mainly in hyperplastic conditions.
Squamous Epithelia
Squamous epithelia were generally negative. In some cases weak focal expression by keratinocytes (cervix, mouth mucosa) was observed but could not be considered safely as positivity.
Mesenchymal Tissues
In general, mesenchymal tissues, except for neural tissue, were negative. Some weak positivity was observed in some myoepithelial cells in the walls of small arteries and in chondrocytes (Figure 7D).
Diffuse Neuroendocrine System
Neuroendocrine cells in several organs expressed hK6.
In short, hK6 is expressed by many normal human tissues. Glandular epithelia constitute the main hK6 immunoexpression sites, with representative organs being breast, prostate, kidney, endometrium, colon, appendix, salivary ducts, bile ducts, and gallbladder. The small intestine, the stomach, the endocervix, the Fallopian tube, the epididymis, the bronchus, and the upper respiratory tract showed focal expression. Choroid plexus epithelium, peripheral nerves, and neuroendocrine cells (including the islets of Langerhans and adrenal medulla) expressed the protein strongly and diffusely. Characteristic immunostaining was observed in Hassall's corpuscles of the thymus, oxyphilic cells in the thyroid and parathyroid glands, in the primordial follicles of the ovary, in dendritic cells mainly in the spleen, and in various cells of the placenta.
Discussion
The kallikrein 6 gene (KLK6) is one of the recently cloned members of the human tissue kallikrein gene family (Diamandis et al. 2000a). Messenger RNA encoding for hK6 protein (zyme/protease M/neurosin) has been previously detected in many tissues, including breast, brain, spinal cord, cerebellum, kidney, uterus, salivary gland, thymus, spleen, and testis in some mammalian species but not in mice, rats, or hamsters (Anisowicz et al. 1996; Little et al. 1997; Ya-mashiro et al. 1997; Yousef et al. 1999). Until recently, no methods were available for the quantitative measurement of hK6 in biological fluids and tissues. The development of polyclonal antibodies and of an immunofluorometric procedure for quantifying the hK6 protein has helped to define the organs that express this protein and its relationship to cancer and other pathological conditions (Diamandis et al. 2000c, d,e). We used recombinant hK6 protein for the development of polyclonal rabbit and mice antibodies and monoclonal mouse antibodies. The developed immunoassay for hK6 (Diamandis et al. 2000c) demonstrates good sensitivity and dynamic range and detects a single immunoreactive band in all biological fluids examined. This proteinase is present in its free form in serum, in contrast to PSA, which is mainly bound to α1-antichymotrypsin (Stenman et al. 1991; Diamandis et al. 2000c).
(
(
In our recent work, we found the highest concentration of hK6 in milk of lactating women, followed by cerebrospinal fluid, nipple aspirate fluid, and breast cyst fluid (Diamandis et al. 2000c). The protein was also detected in male and female serum, in the majority of seminal plasmas, and in a relatively small percentage of amniotic fluids and breast tumor cytosolic extracts, but not in urine. The examination of a number of human tissue extracts revealed high concentrations of hK6 in the salivary glands, lung, colon, Fallopian tube, placenta, breast, pituitary, and kidney. We further found that hK6 concentration in brain tissue extracts from patients with Alzheimer's disease was lower than that in normal controls; however, the CSF and serum hK6 concentrations were higher in patients with the disease (Diamandis et al. 2000e). Using the rabbit polyclonal antibody, we localized hK6 by IHC in normal epithelial cells of the choroid plexus. Therefore, hK6 may constitute a new biomarker for diagnosis and monitoring of Alzheimer's disease (Diamandis et al. 2000e).
This is the first report describing IHC localization of hK6 in a large variety of human tissues. Only Little et al. (1997) have previously studied hK6 immunohistochemically, and only in brain tissues. Because we used exclusively normal tissues, it will be interesting to follow up these studies with tissues from various pathological conditions, to establish if hK6 has value as a disease biomarker.
hK6 was localized in a large number of normal human tissues and therefore cannot be considered as a specific tissue marker, in contrast to the homologous proteins hK2 and PSA, which show prostate-restricted specificity (McCormack et al. 1995; Rittenhouse et al. 1998). With the availability of highly sensitive techniques, it has become apparent that these two kallikreins are also expressed in many other tissues, including breast, thyroid, and salivary glands (Black and Diamandis 2000; Maglara et al. 2000). The expression of hK6 mainly by glandular epithelia suggests that it is secreted. This is further supported by the finding of hK6 in many biological fluids (Diamandis et al. 2000c). The absence of hK6 in urine corresponds well with the lack of immunoreactivity by the urothelium. The positive immunoexpression of glial cells in our study is in accordance with previous findings (Little et al. 1997; Yamanaka et al. 1999). Yamanaka et al. (1999) found KLK6 mRNA in mature oligodendrocytes and suggested that this protease may be important in the processes occurring after maturation of oligodendrocytes, such as myelination or turnover of the proteins in the myelin. Little et al. (1997) detected positive immunostaining in monkey cortex cells lining the perimeter of cortical microvessels, in human brains of patients with Alzheimer's disease, and in microglial cells, indicating a role of this protease in brain disease. We localized hK6 also in the choroid plexus epithelium and correlated our findings with Alzheimer's disease (Diamandis et al. 2000e). Yamashiro et al. (1997) found strongest KLK6 mRNA expression in brain, followed by expression in spleen, in accordance with the immunoreactivity of the Hassall's corpuscles of the spleen in our study. An interesting finding is the expression of hK6 in oxyphilic cells in the thyroid and parathyroid glands. This accords with demonstration of hK2 and hK3 in oxyphilic cells of the thyroid (Maglara et al. 2000), suggesting another similarity among these three kallikreins. The detection of hK6 in human male and female serum (Diamandis et al. 2000c) is indicative that it is secreted by various tissues. Worth mentioning is the previous demonstration of other kallikreins, including hK3 (PSA) and hK2, in breast secretions, breast cancer cytosols, seminal plasma, and amniotic fluids (Yu and Diamandis 1995a,b; Black et al. 2000; Black and Diamandis 2000).
Another interesting finding was the presence of hK6 in some cells of the diffuse neuroendocrine system and in nerves. The staining was relatively strong, and we believe that it could be representative of neuroendocrine differentiation. The expression of hK6 is reminiscent of CD56 (natural killer cell-associated antigen, neural cell adhesion molecule) and CD57 (Leu-7, T-cell surface marker) which are sensitive but not specific for cells and neoplasms with neuroendocrine differentiation (Kaufmann et al. 1997). These suggestions need verification with a study of a variety of neuroendocrine tumors. Furthermore, we found strong hK6 expression in the islets of Langerhans and in the epithelium of the pancreatic ducts, but not in the acinar cells of the exocrine pancreas. It is possible that hK6 is involved in islet hormone processing, a property already attributed to many other proteolytic enzymes (Seidah and Chretien 1999).
(
(
(
The contribution of hK3 (PSA) and hK2 in the diagnosis and monitoring of prostate cancer suggests that other kallikreins may also have value as candidate biomarkers. We have already shown that serum hK6 concentration is increased in ovarian carcinoma (Diamandis et al. 2000d). Some other kallikrein genes are either underexpressed or overexpressed in certain carcinomas (Yousef and Diamandis 2001). The encoded proteases may serve to positively or negatively regulate cell growth or differentiation by cleavage of cell surface receptors (Coughlin 1999) or cell growth regulatory and angiogenic proteins, and by activation of other proteases for invasion and metastasis. These serine proteases are secreted into the extracellular space and therefore are excellent candidate circulating tumor markers. In addition to KLK2 and KLK3, other kallikrein genes have been studied in relation to oncogenesis and prognosis of malignant tumors (Diamandis et al. 2000a; Yousef and Diamandis 2001). Liu et al. (1996) reported downregulation of KLK10 in cancerous breast and prostate tissues and in cancer cell lines. Others reported upregulation of TADG-14/KLK8 and KLK7 in ovarian carcinomas (Tanimoto et al. 1999; Underwood et al. 1999). Breast, prostate, renal, and gastrointestinal cancer may reveal some relation to hK6 expression, as these organs strongly express this protein.
KLK6 has been shown to be regulated by steroid hormones (Yousef et al. 1999). It will be interesting to study steroid hormone-responsive organs for hK6 expression and to investigate its possible involvement in tumor development, progression, or both (Diamandis et al. 2000a; Yousef and Diamandis 2001). Some kallikreins might be good therapeutic targets and the enzymatic activity of these proteins might be beneficial or deleterious. The development of tissue kallikrein inhibitors or activators may provide a new generation of drugs against cancer and other disorders.