
Abstract
The tetraspan membrane protein CD9 is normally expressed in the mature myelin sheath and is believed to suppress the metastatic potential of certain human tumors. In this study we identified CD9 in a variety of brain tumors by immunohistochemical (IHC) and immunoblotting analyses. We examined 96 tumor samples and three glioma cell lines in addition to a murine brain tumor model of transplanted glioma cells in CD9-deficient mice and control mice. CD9 was expressed not only in solid non-neuroepithelial tumors but also in infiltrative malignant neuroepithelial tumors. Among the neuroepithelial tumors, high-grade astrocytic tumors, including glioblastomas and anaplastic astrocytomas, showed higher immunoreactivity than low-grade cerebral astrocytomas. Thus, CD9 expression in astrocytic tumors correlated with their malignancy. In the murine brain tumor model, transplanted glioma cells were shown to grow and spread through myelinated areas irrespective of the presence or absence of CD9 expression in the recipient's brain. These results indicate that the CD9 expression of astrocytic tumors plays a significant role in the malignancy independent of CD9 expression in the surrounding tissue. This might be explained by the observation that the CD9 molecule is associated with a mitogenic factor, membrane-anchored heparin-binding epidermal growth factor, which is known to be upregulated in malignant gliomas.
CD9 is a cell surface glycoprotein of the tetra-membrane-spanning protein family. It is a constituent of cell surface complexes that include the integrins, other cell surface receptors, and other tetraspanins (Horejsi and Vlcek 1991; Rubinstein et al. 1994, 1996; Nakamura et al. 1995; Berditchevski et al. 1996; Hadjiargyrou et al. 1996; Radford et al. 1996; Slupsky et al. 1997). CD9 was originally characterized as a cell surface antigen on lymphohemopoietic cells (Boucheix and Benoit 1988). Platelets, pre-B-cells, mature oligo-dendrocytes, and Schwann cells are abundant in CD9 (Kersey et al. 1981; Jennings et al. 1990; Boucheix et al. 1991; Griffith et al. 1991; Tole and Patterson 1993; Kaprielian et al. 1995; Nakamura et al. 1996; Banerjee et al. 1997; Kagawa et al. 1997), and its expression is also detected in astrocytes and microglia in vitro (Deissler et al. 1996; Schmidt et al. 1996; Dohura et al. 2000), as well as in sympathetic neurons, dorsal root ganglion cells, and adrenal chromaffin cells in the early embryonic peripheral nervous system (Griffith et al. 1991; Tole and Patterson 1993). CD9 and other tetraspanins have been reported to participate in the activation, adhesion, and motility of cells and in normal and tumor cell growth (Maecker et al. 1997). For example, anti-CD9 antibody binding triggers platelet activation and aggregation (Griffith et al. 1991; Kroll et al. 1992). Recently, it was proved essential for sperm-egg fusion (Miyado et al. 2000; Naour et al. 2000). CD9 is localized mainly along the outermost membrane of compact myelin in the central nervous system (CNS) (Nakamura et al. 1996), and its expression developmentally appears at the end of the myelination process, increases with age, and persists in the adult brain (Kagawa et al. 1997). However, the function of CD9 in the CNS still remains unclear.
CD9 Immunoreactivity in brain tumors a
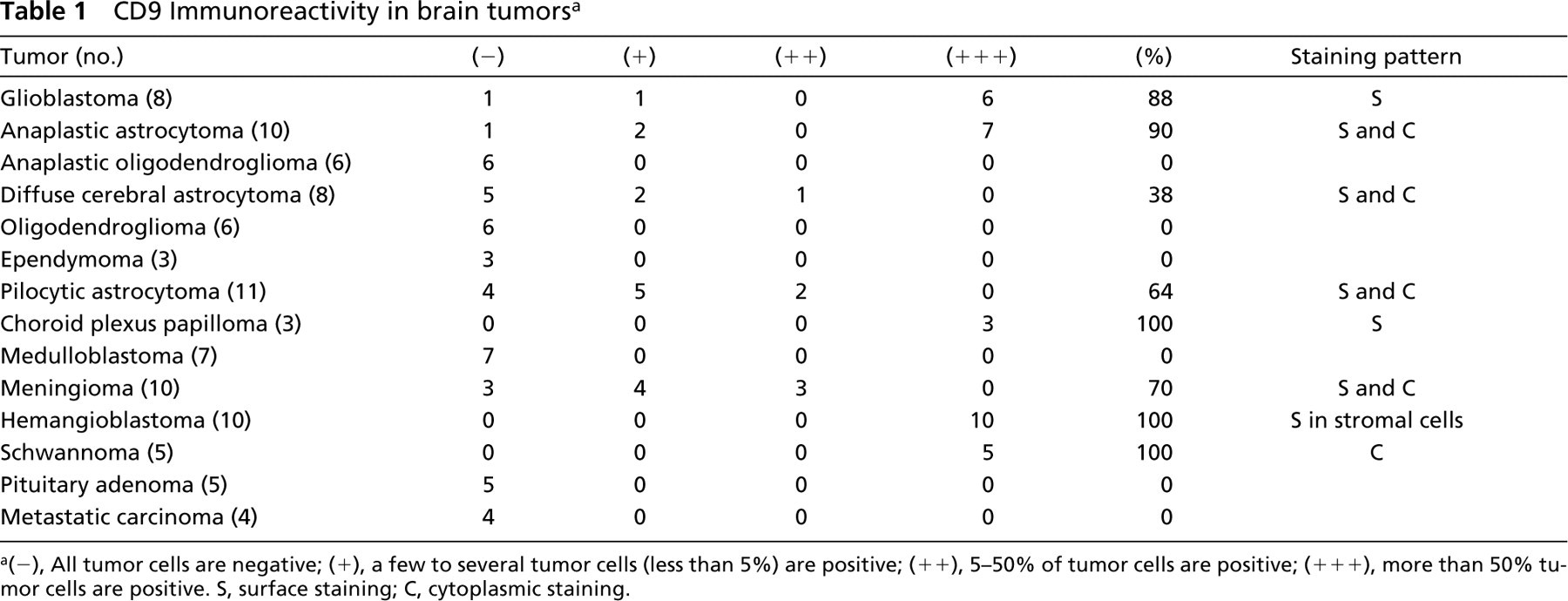
(−), All tumor cells are negative; (+), a few to several tumor cells (less than 5%) are positive; (++), 5-50% of tumor cells are positive; (+++), more than 50% tumor cells are positive. S, surface staining; C, cytoplasmic staining.
On the other hand, CD9 is considered to inhibit cell motility and is referred to as motility-related protein 1 (MRP-1) (Ikeyama et al. 1993). It has been demonstrated that the cells induced to express CD9 show suppressed cell motility in vitro (Ikeyama et al. 1993). An inverse correlation was found between CD9 expression and metastases in breast cancer (Miyake et al. 1995), and reduced CD9 gene expression resulted in poor prognoses in lung cancer (Higashiyama et al. 1995).
Here we studied the significance of the CD9 molecule in brain tumors by the immunohistochemical (IHC) analyses of brain tumor samples and a murine brain tumor model.
Materials and Methods
CD9 IHC in Normal Human Cerebrum and Spinal Cord
CD 9 immunoreactivity in a normal cerebrum and in a spinal nerve root and ganglion (autopsy samples) was examined.
Patients and Surgical Specimens
The tumor tissues listed in Table 1 were obtained at surgery from 96 patients (43 males and 53 females, aged 1 month to 79 years) at the Department of Neurosurgery, Kyushu University Hospital. A normal cerebrum (autopsy samples) was used as a control.
Antibodies and IHC
Data on antibodies are listed in Table 2. The indirect immunoperoxidase method was performed on paraffin sections of brain tumors. Surgical specimens were fixed in 10% neutral buffered formalin overnight and embedded in paraffin. The samples were then cut into 5-μm-thick sections. The sections were deparaffinized in xylene and hydrated in graded ethanols. Endogenous peroxidase activity was blocked with 0.3% H2O2 in absolute methanol for 30 min at room temperature (RT). After rinsing in tapwater, the sections were completely immersed in distilled water. Thereafter the sections were pretreated specifically as listed in Table 2. The sections were then washed in Tris buffer (TB; 50 mM Tris-HCl, pH 7.6), followed by overnight incubation with the primary antibodies at 4C. After washing in TB, the sections were incubated with horseradish peroxidase-conjugated secondary antibody (1:200 dilution; Vector Laboratories, Burlingame, CA). The colored reaction product was developed with 0.02% 3,3′-diaminobenzidine tetrahydrochloride (DAB), 0.003% H2O2, and 50 mM Tris-HCl, pH 7.6. The sections were counterstained lightly with hematoxylin. The tests were done with an appropriate positive control (normal cerebrum).
Data of antibodies

Cultured Cells
Human glioma cell lines KNS81 (Takeshita et al. 1990) and U-373MG (Pontén and Westermark 1987) and a mouse glioma cell line, G26-24 (Neskovic et al. 1981) were cultivated to 60-80% confluency in Dulbecco's modified Eagle's medium (GIBCO BRL; Gaithersburg, MD), containing 10% fetal bovine serum in a humidified atmosphere at 37C, in 5% CO2.
Immunocytochemistry of the Cultivated Human and Mouse Glioma Cells
After rinsing the cultivated human glioma cells KNS81 and U-373MG and the mouse glioma cells G26-24 in PBS, the cells were fixed with 10% buffered formalin for 30 min at RT. After rinsing in PBS, the human glioma cells were incubated with anti-human CD9 MAb (007) and mouse glioma cells with anti-mouse CD9 (KOBA4) overnight at 4C. Then the cells were incubated with horseradish peroxidase-conjugated secondary antibody (1:200 dilution). After rinsing in PBS, the colored reaction product was developed with DAB.
Immunoblotting Analysis
The specificities of two MAbs (clones 007 and 72F6) and a polyclonal antibody against CD9 in tumor tissue were assessed using soluble fractions extracted from human glioma cell lines KNS81 and U-373MG, a mouse glioma cell line G26-24, an autopsied human cerebrum, a fresh control, and CD9-deficient mice cerebrums and from frozen tumor samples of a hemangioblastoma, a choroid plexus papilloma, a meningioma, and a schwannoma. The samples were homogenized in 10 volumes of buffer containing 150 mM NaCl, 5 mM EDTA, 0.5% sodium deoxycholate, 0.5% Nonidet P-40, 5 mM Tris-HCl, pH 7.0. The protein concentrations were determined by the modified Lowry's procedure using bovine serum albumin as the protein standard. Laemmli's sample buffer was added to this mixture and the samples were boiled for 5 min. Each protein sample (10 μg/lane) was separated on a 15% SDS-polyacrylamide gel and transferred to a polyvinylidene difluoride membrane (Millipore; Bedford, MA). After blocking with 5% low-fat milk in TBST (25 mM Tris-HCl, pH 7.6, 0.15 M NaCl, 0.05% Tween-20, 0.05% NaN3), the membrane was incubated with anti-CD9 (either 007 or 72F6) MAb (1:100) or anti-mouse CD9 (KOBA4) polyclonal antibody (1:100) in TBST containing 5% low-fat milk at 4C overnight. The membrane was then washed in TBST and incubated with alkaline phosphatase-conjugated secondary antibody (1:7500 dilution; Promega, Madison, WI) in TBST containing 5% low-fat milk for 30 min. After washing the membrane in TBST, the signal was visualized with CDP-Star detection reagent (Amersham; Arlington Heights, IL) according to the manufacturer's instructions.
Murine Brain Tumor Model
The G26-24 glioma cell line was inoculated as a suspension into mouse brains. CD9−/− mice were developed as reported previously (Miyado et al. 2000). Six CD9+/+ mice, 11 CD9+/− mice, and 14 CD9−/− mice were used. The G26-24 glioma cells were kept at 4C before injection. Using the tip of a 24-gauge needle, a hole was drilled in the calvaria overlying the right parietal cortex, 2 mm caudal to the coronal suture and 3 mm lateral to the midline. Mice were injected with 8.0 × 104 cells in 3 μl volume at a depth of 3 mm using a Hamilton microsyringe. The glioma cells were delivered very slowly (30 sec) while the needle was being withdrawn 1 mm upward and then left in place for an additional 30-50 sec. The needle was withdrawn 1 mm upward, again, and left in place for 30-50 seconds before complete needle removal. The needle was cleansed of tumor cells with alcohol before the next injection.
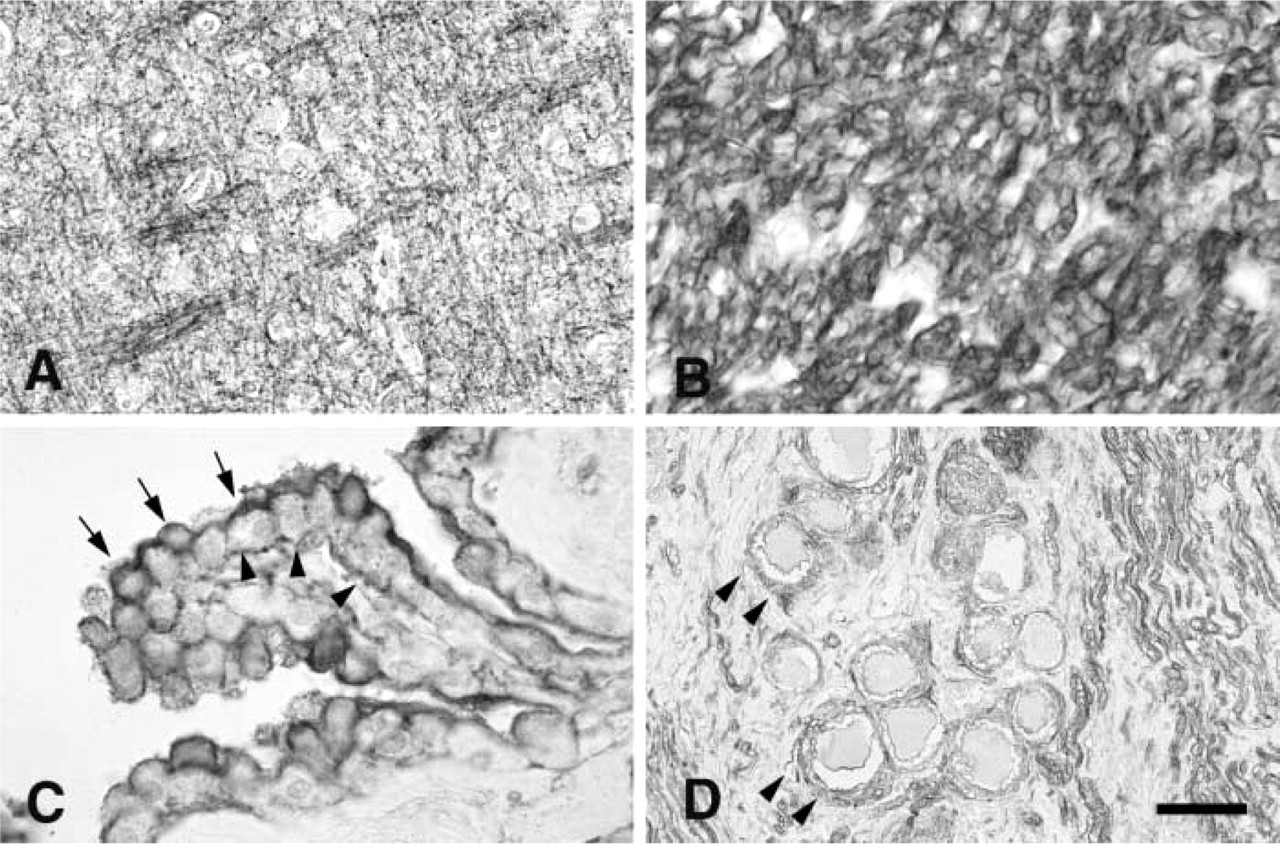
Immunostaining of normal human cerebrum probed with anti-CD9 antibody (MAb 007).
After the tumor implantation, the hole was disinfected with ethanol and 10% iodine. The incisions were sutured and the mice were placed on a heating pad for recovery before being returned to their cages. Tumors were allowed to grow 7, 14, or 21 days, and their development was studied on coronal sections at these survival times. The mice were divided into three groups. Group 1 mice (n = 9, CD9+/+: CD9+/−:CD9−/− = 2:3:4) were maintained for 7 days, those of group 2 (n = 13, CD9+/+:CD9+/−:CD9−/− = 3:4:8) for 14 days, and those of group 3 (n = 8, CD9+/+:CD9+/−:CD9−/− = 1:4:3) for 21 days after tumor implantation. Mice were then sacrificed by cervical dislocation and the brains were dissected for histological and IHC analyses. IHC staining was performed by the indirect immunoperoxidase method for CD9, glial fibrillary acidic protein (GFAP), myelin basic protein (MBP), and Ki-67 (Table 2). Ki-67 antibody was used to examine how the tumor cells grow and spread in the recipient's brain because it is difficult to distinguish the tumor cells from the normal myelin sheath by using CD9 antibody. The largest area of the tumor in coronal section stained with hematoxylin and eosin (HE) was measured using a micrometer. Then statistical analysis using the Mann-Whitney U-test was performed between control and CD9-deficient mice in every group.
Results
CD9 IHC in Normal Human Cerebrum and Spinal Cord
Increased CD9 (MAbs 72F6 and 007) immunoreactivity was obvious in the myelin sheath of the central and peripheral nervous system, choroid plexus, and satellite cells in the spinal ganglion (Figures 1A–1D). Positive immunoreactivity was observed on both apical and basal sides in normal choroid plexus (Figure 1C).
CD9 IHC in Brain Tumors
The results of the IHC of 96 brain tumors are summarized in Table 1.
Neuroepithelial Tumors. In astrocytic tumors, CD9 (MAbs 72F6 and 007) expression correlated with the tumor's histological grade. Glioblastomas and anaplastic astrocytomas showed frequent and strong CD9 expression (Figures 2A and 2B). Diffuse cerebral astrocytomas showed mainly surface staining for CD9 (Figure 2C). To distinguish CD9 expression of tumor cells from normal myelin staining, we examined involvement of the myelin sheath in the tumor specimens by using an antibody against MBP in consecutive sections (Figures 2D). Other infiltrative neuroepithelial tumors, including oligodendroglioma, anaplastic oligodendroglioma, and medulloblastoma, were completely immunonegative to CD9. Pilocytic astrocytomas showed mainly cytoplasmic immunoreactivity to CD9. Ependymomas showed no immunoreactivity to CD9. All cases of choroid plexus papilloma showed surface staining resembling normal choroid plexus. However, it was restricted to the apical side (Figure 3A).
Non-neuroepithelial Tumors. CD9 (MAbs 72F6 and 007) immunoreactivity was detected in certain solid benign tumors. Diffuse and cytoplasmic staining was observed in all schwannomas (Figure 3B). Immunostaining for CD9 in stromal cells in hemangioblastomas was detected in all cases (Figure 3C). Seven of 10 meningiomas showed both surface and cytoplasmic staining (Figure 3D). Pituitary adenomas showed no immunoreactivity. All four metastatic carcinomas were totally immunonegative to CD9.
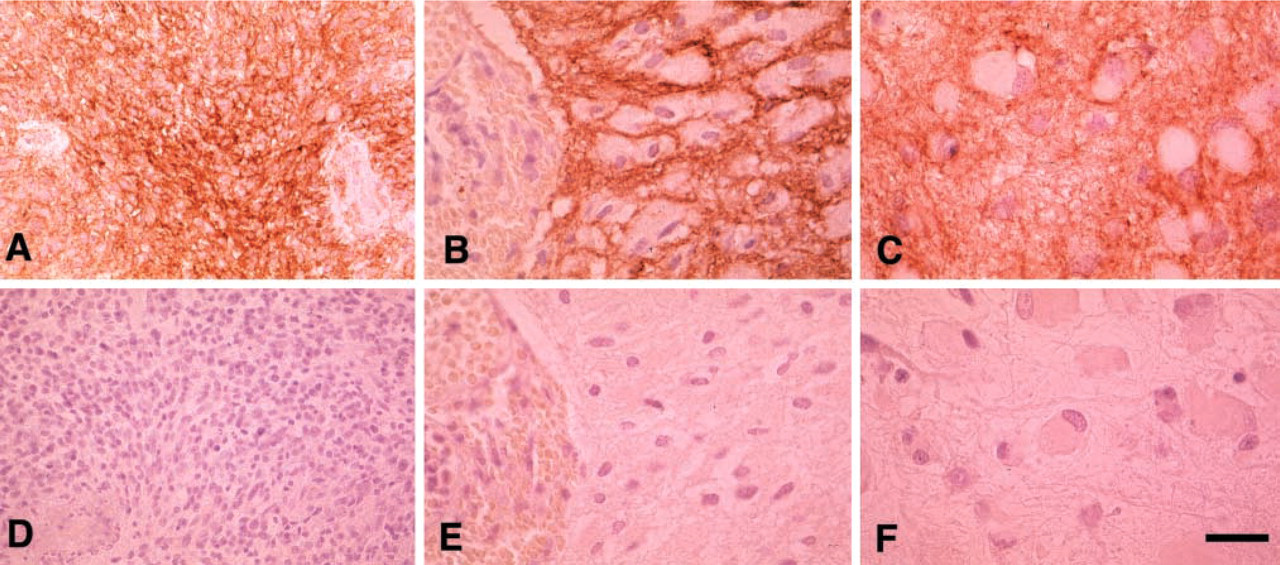
Immunostaining of astrocytic tumors probed with anti-CD9 antibody (MAb 72F6,
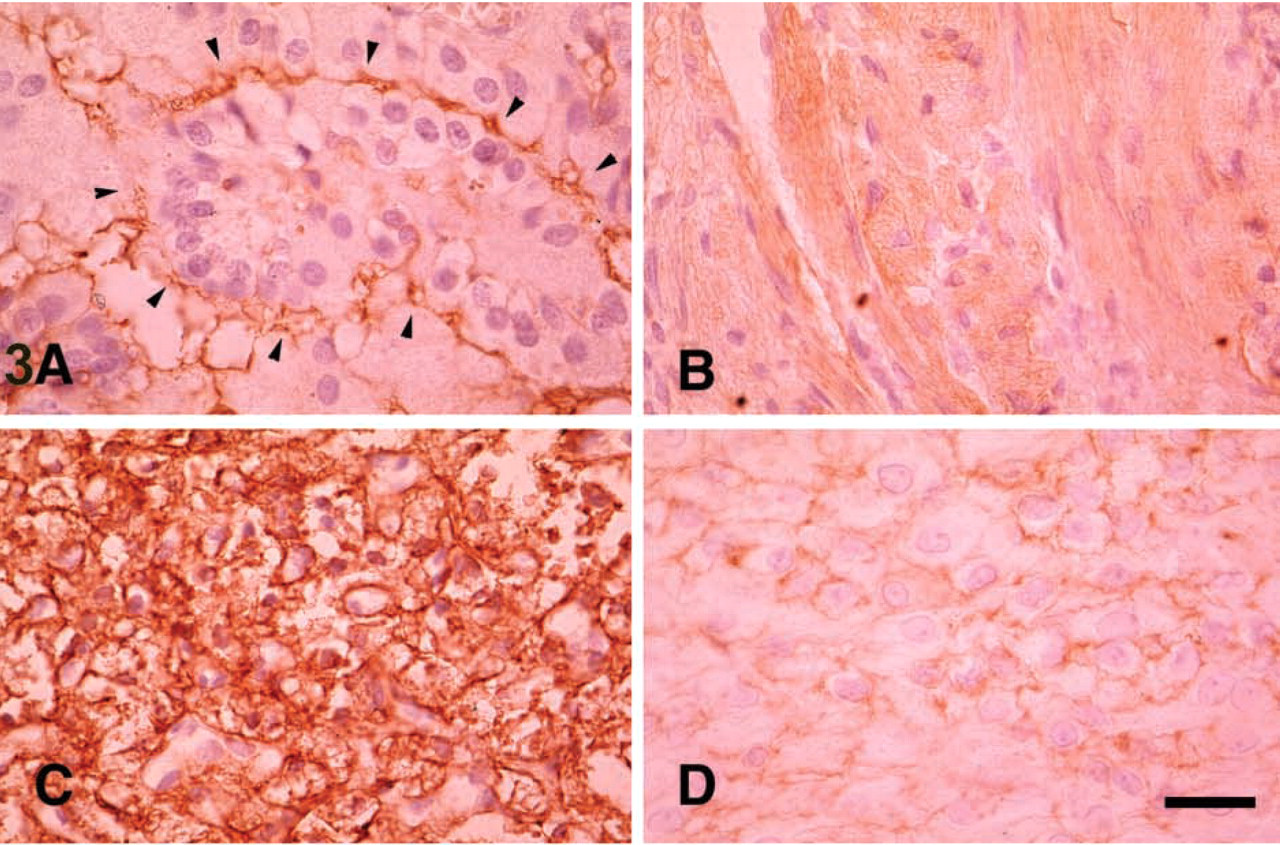
Immunostaining of solid benign tumors probed with anti-CD9 antibody (MAb 72F6).
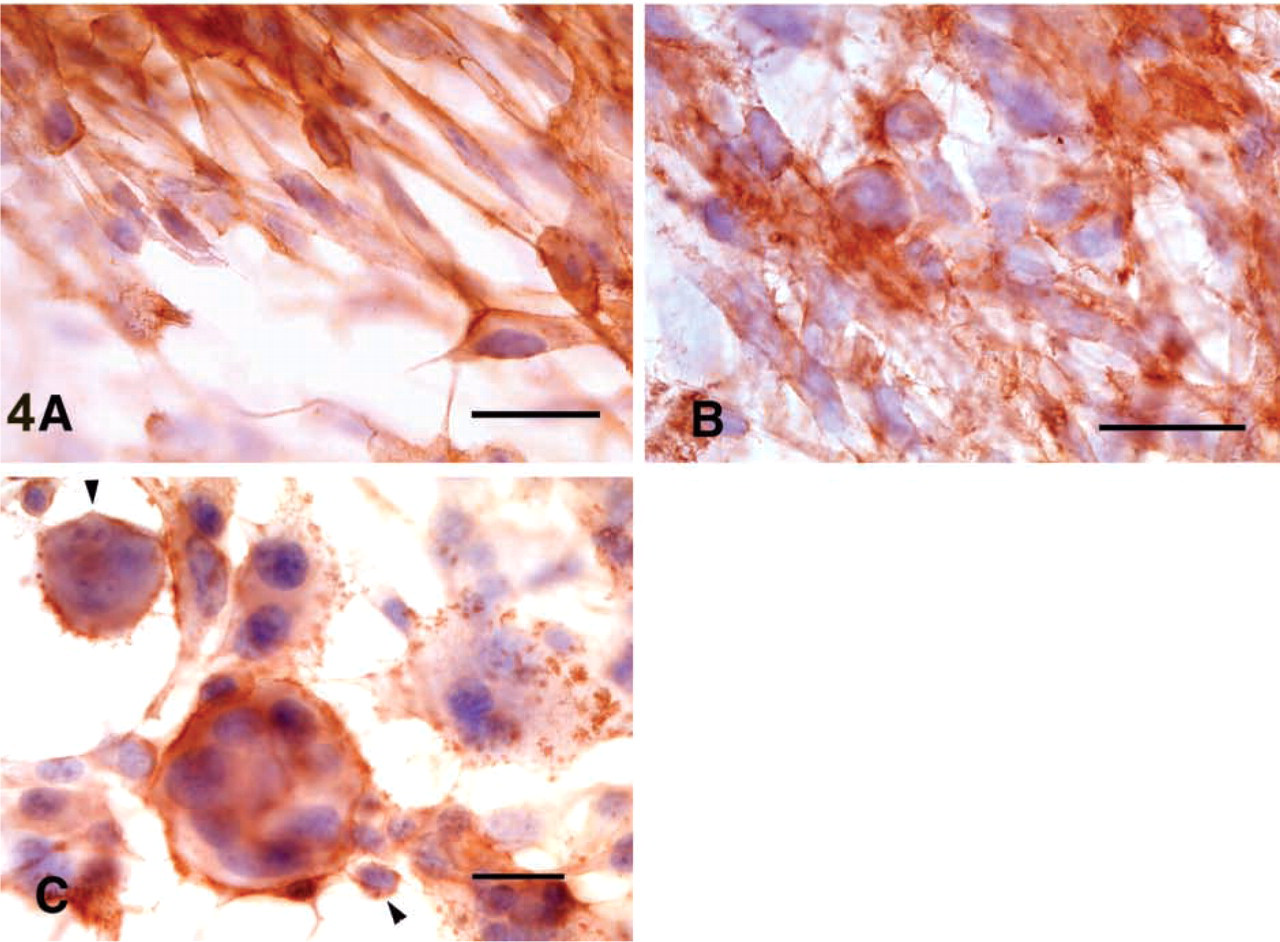
Glioma Cell Lines. We also investigated immunoreactivity to CD9 (MAbs 72F6 and 007) in human glioma cell lines KNS81 and U-373MG and in a mouse glioma cell line, G26-24. The human glioma cells showed surface immunostaining (Figure 4A and 4B). Mouse glioma cell line G26-24 showed short spicule-like surface staining in a multinucleated giant cell and in a mononuclear cell (Figure 4C). Immunoreactivity of the 72F6 antibody for cultured cells was much weaker than that of 007.
Western Blotting
The results of immunoblotting for benign solid tumors and culture cells are shown in Figure 5. The extract from adult human cerebral white matter showed a single band of approximately 29 kD when probed with anti-CD9 MAb 007 (Figure 5a, Lane 1). A hemangio-blastoma, a choroid plexus papilloma, a meningioma and a schwannoma (Figure 5a, Lanes 2-5) also showed single or doublet bands identical to that of the normal adult brain sample, thus confirming the specificity of the anti-CD9 MAb. Although the reason for the doublet is not clear, it may be due to a difference in glycosylation. On the other hand, the extracts from the cultured cells KNS81 and U-373MG (Figure 5b, Lanes 1 and 2) also showed doublet or triplet bands corresponding to CD9.
The extracts from fresh cerebral white matter of a CD9-deficient mouse (Figure 5c, Lane 1) showed no specific band, however, fresh cerebrum of a control mouse (Figure 5c, Lane 2) and cultured cells of G26-24 (Figure 5c, Lane 3) showed single bands corresponding to CD9.
Histological Evaluation and Tumor Size Measurement of Mouse Glioma G26-24 Model In Vivo
Using antibodies to MBP and Ki-67, we examined the histology of control mouse brains (CD9+/+ and CD9+/−) and CD9−/− mouse brains in each group after intracerebral inoculation of G26-24 cells. They showed no significant difference in their tumor growth pattern. Tumor cells developed as solid intraparenchymal tumors at the implantation site. G26-24 cells had infiltrated into the recipients' brains mainly through myelinated areas in every group, which was demonstrated by the presence of Ki-67-immunolabeled cells. This pattern was observed in all mice, whether they expressed CD9 or not (Figures 6A and 6B). Similar GFAP expression in astrocytes adjacent to the tumor was observed in every case examined. The average of the largest area of tumor in control and CD9−/− mice in every group also showed no significant difference (p>0.05).
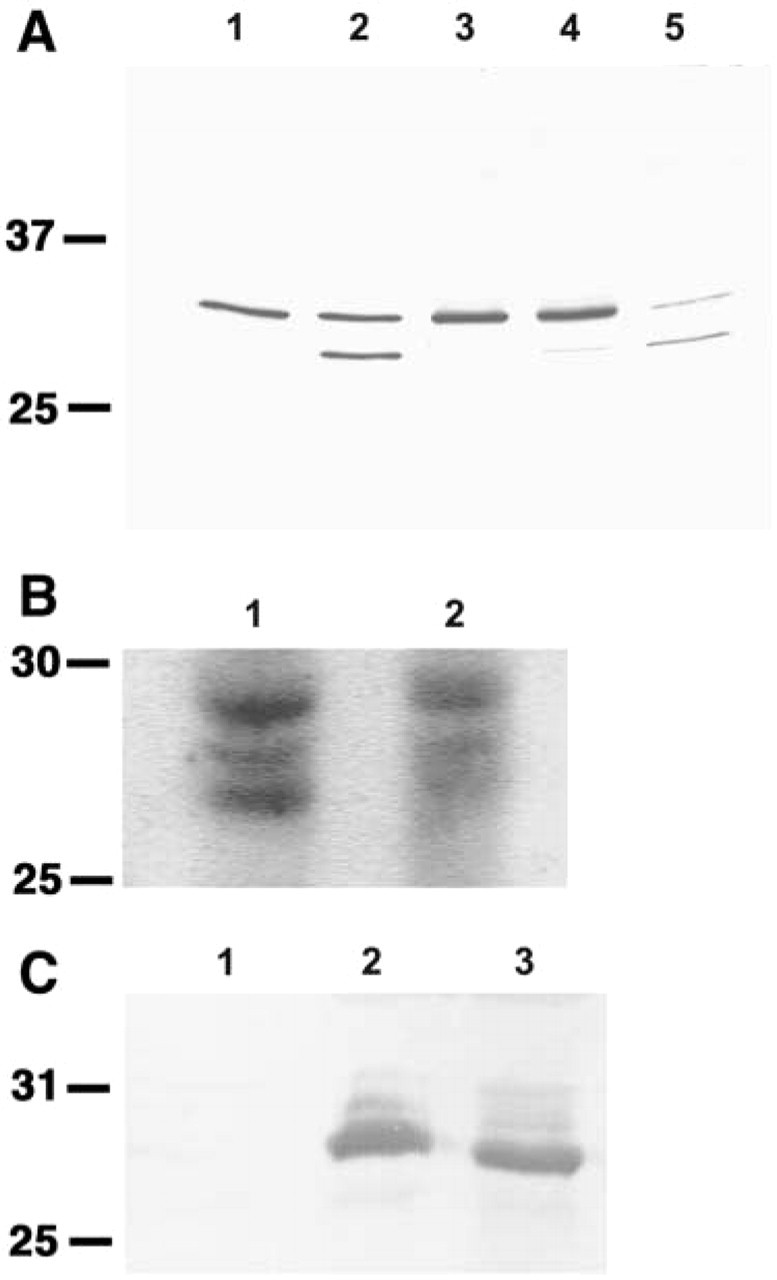
An immunoblot probed with anti-CD9 (MAb 007,
Discussion
Here we demonstrated that the CD9 molecule is expressed on the tumor cell surface and/or cytoplasm of a variety of brain tumors, including not only solid benign brain tumors but also some malignant brain tumors. We found that the CD9 expression correlated well with the malignancy of the astrocytic tumors. Finally, in the murine brain tumor model, we showed that the CD9-expressing glioma cells grow and invade the surrounding brain tissue in a similar way regardless of the presence or absence of the CD9 molecule in the surrounding tissue.
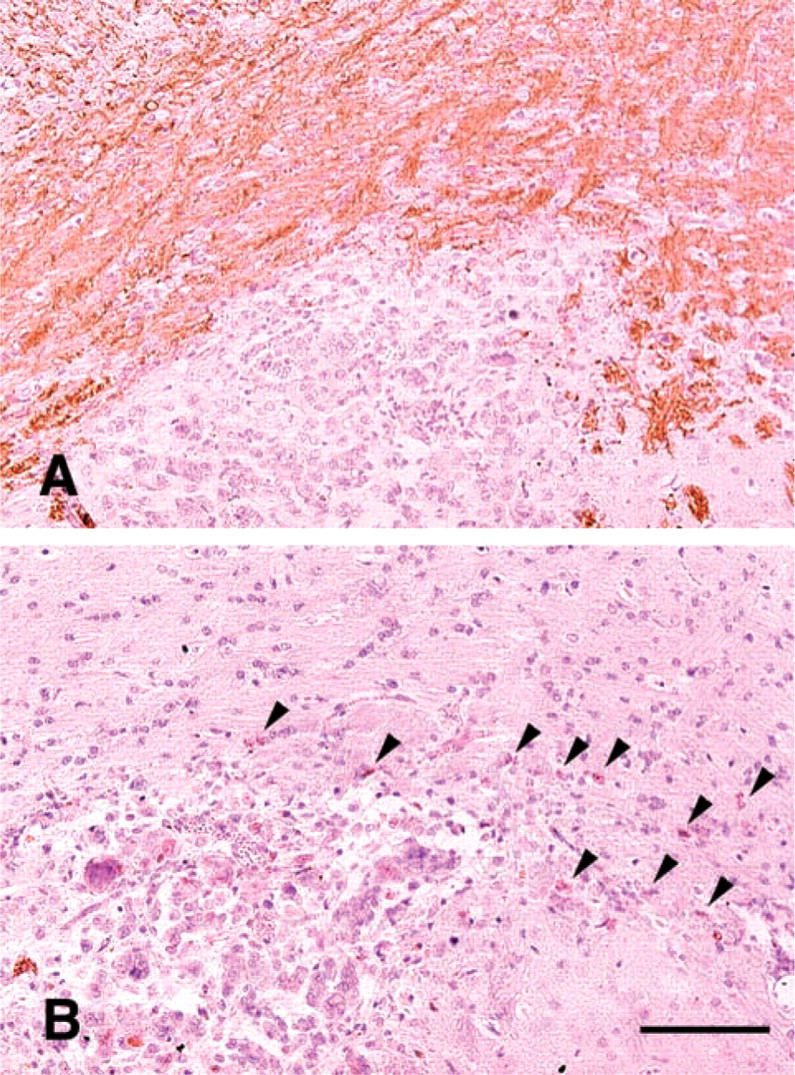
Immunostaining for MBP
The finding of the expression of CD9 molecules in invasive malignant astrocytic tumors, such as anaplastic astrocytoma and glioblastoma, appears to be inconsistent with the previous reports on non-brain tumors, including breast, lung, esophageal, colon, and pancreatic cancers (Higashiyama et al. 1995; Miyake et al. 1995; Cajot et al. 1997; Sho et al. 1998; Shimada et al. 1999). In these non-brain tumors, CD9 is known as MRP-1, and its expression is reported to correlate inversely with the metastatic potential of the tumors. Therefore, the positive correlation of the CD9 expression with the histological grade of the astrocytic tumors suggests that the CD9 molecule has other function(s) than that reported in those non-brain tumors.
One possible explanation of the correlation between the CD9 expression and the invasive malignancy of astrocytic tumors is that the CD9 expression in the malignant astrocytic tumors might reflect the cell growth associated with the heparin-binding epidermal growth factor-like growth factor (HB-EGF) (Nakamura et al. 1995). The CD9 molecule is reported to form a complex with a membrane-anchored mitogenic factor, HB-EGF (Nakamura et al. 1995), and HB-EGF is suggested to contribute to the growth of malignant gliomas. Mishima and colleagues (1998) reported that both HB-EGF expression and epidermal growth factor receptor expression are enhanced in malignant gliomas.
Another possible explanation is that the CD9 molecule in the malignant astrocytic tumors plays a role in the adhesion of glioma cells to the extracellular matrix in a mechanism involving integrins. Many cell surface receptors are involved in the cell adhesion, migration, and invasion of the glioma cells. Among them, the major role is played by integrins or the hyaluronate receptor CD44 in glioma cell-matrix adhesion (Merzak et al. 1994; Friedlander et al. 1996), and the CD9 molecule is reported to be strongly associated with the integrins (Boucheix et al. 1991; Deissler et al. 1996; Schmidt et al. 1996; Banerjee et al. 1997; Kagawa et al. 1997).
The expression of the CD9 molecule in such benign brain tumors as hemangioblastoma, choroid plexus papilloma, meningioma and schwannoma might be consistent with the idea of MRP-1 as an equivalent of CD9. Alternatively, in schwannoma and choroid plexus papilloma, it is also plausible that the neoplastic counterparts express the CD9 molecule, because the CD9 molecule is expressed in the myelin and choroid plexus of normal brain. However, this explanation does not apply to meningioma and hemangioblastoma, and the association of the CD9 molecule with the genesis of these tumors remains unclear.
On the other hand, non-astrocytic gliomas, including oligodendroglioma, anaplastic oligodendroglioma, and ependymoma, did not show any immunoreactivity to CD9. This indicates that the CD9 molecule is no longer expressed in the neoplastic counterparts in these brain cells, unlike schwannoma and choroid plexus papilloma. In addition, the CD9 molecule might not help the non-astrocytic tumor cell migration, but the astrocytic tumor cells may migrate in a similar way to that detected in reactive astrocytes in transmissible spongiform encephalopathies (TSE), which we recently described in mouse and human brains (Doh-ura et al. 2000). In brains infected with TSE, enhanced CD9 expression in astrocytes is associated with migration and proliferation of the astrocytes.
Finally, we demonstrated that CD9 expression in the brain tissue surrounding the glioma cells never influenced the growth and invasion of the tumor cells. This suggests that the cell surface molecules and/or growth factors of glioma cells might not necessarily interact with the CD9 molecule in the surrounding tissue, especially in the myelin sheath along which glioma cells migrate.
In conclusion, we have demonstrated the first direct evidence that the CD9 molecule is expressed not only in solid non-neuroepithelial brain tumors but also in infiltrative astrocytic tumors. Our data indicate that the CD9 molecule could have function(s) other than that of MRP-1 in some brain tumors. To understand fully the roles of the CD9 molecule in brain tumors, further studies of the functional relationship between the CD9 molecule and the secretion of proteases or growth factors, as well as of the mechanisms of signal transduction of the CD9 molecule on the cell membrane, remain.
Footnotes
Acknowledgments
We thank Ms. K. Hatanaka, Mr. T Kanemaru, and Ms. A Kawashima for their excellent technical assistance, and Ms. K. Ono for reviewing the manuscript.