
Abstract
The control of extracellular matrix (ECM) production is important for the development, maintenance, and repair of cartilage tissues. Matrix molecule synthesis is generally regulated by the rate of gene transcription determined by DNA transcription factors. We have shown that transcription factors Sox9, AP-2, and [delta]EF1 are able to alter the rate of CD-RAP transcription in vitro: Sox9 upregulates, AP-2 exhibits biphasic effects, and [delta]EF1 represses expression of the CD-RAP gene. To correlate these in vitro activities in vivo, transcription factors were co-immunolocalized with ECM proteins in three different cartilage tissues in which the rates of biosynthesis are quite different: articular, meniscal, and growth plate. Immunoreactivities of type II collagen and CD-RAP were higher in growth plate than in either the articular or meniscal cartilages and correlated positively with Sox9 protein. Sox9 staining decreased with hypertrophy and was low in articular and meniscal cartilages. In contrast, AP-2 and [delta]EF1 were low in proliferating chondrocytes but high in lower growth plate, articular, and meniscal cartilages. This increase was also accompanied by intense nuclear staining. These immunohistochemical results are the first to localize both [delta]EF1 and AP-2 to adult articular, meniscal, and growth plate cartilages and provide in vivo correlation of previous molecular biological studies.
T
The mechanism of transcriptional regulation of chondrogenesis is not understood in detail. Significantly, however, transcription factor Sox9, a close relative of the sex-determining region Y protein (SRY), was shown to have an activating role in chondrogenesis by enhancing type II collagen gene expression (Bell et al. 1997; Lefebvre et al. 1997; Ng et al. 1997) and to be necessary for cartilage formation (Bi et al. 1999). Mutations in the human Sox9 gene are known to cause campomelic dysplasia, resulting in severe abnormalities of skeletal tissues (Foster et al. 1994; Wagner et al. 1994). Co-expression of Sox9 and type II collagen genes has been demonstrated for both embryogenic chondrogenesis and in the growth plate of newborn mice (Wright et al. 1995; Ng et al. 1997; Zhao et al. 1997).
Our studies have focused on three transcription factors, Sox9, AP-2 and [delta]EF1 (also called ZEB1), which are important for regulating the chondrocyte genes, type II collagen, and cartilage-derived-retinoic acid-sensitive protein (CD-RAP). We have shown that, like type II collagen, Sox9 enhances CD-RAP expression via binding to a corresponding consensus region in its promoter (Xie et al. 1999). We have also shown that the transcriptional activities of CD-RAP and type II collagen are regulated by the transcription factor AP-2 in a biphasic manner. Low doses of AP-2 increased the transcriptional activity of CD-RAP and type II collagen promoters, whereas high doses of AP-2 repressed these promoters (Xie et al. 1998; Purple et al. 2001). Most recently, the bHLH transcription factor [delta]EF1 has been shown to downregulate both the type II collagen (Murray et al. 2000) and CD-RAP (Li and Sandell 2002) promoter activities.
Previous reports on transgenic and knockout mice for each of these transacting factors clearly indicate that they are important for regulation of cartilage differentiation during development because mutations in each demonstrate pronounced skeletal defects (Schorle et al. 1996; Zhang et al. 1996; Nottoli et al. 1998; Takagi et al. 1998; Bi et al. 1999). However, it is not known whether these factors play a role in regulating the mature cartilage phenotype. Our in vitro studies on the regulation of CD-RAP and type II collagen suggest that AP-2 and [delta]EF1 play an essential role in suppressing cartilage matrix proteins. However, it is critical to demonstrate that this effect can be correlated in vivo. For most transacting factors, activity is associated with their availability in the nucleus, including the transcriptional repressor [delta]EF1 (Franklin et al. 1994). We have previously shown that the level of steady-state CD-RAP and type II collagen mRNAs, as determined by Northern blotting analysis, closely parallels the intensity of protein immunostaining within the ECM (Sakano et al. 1999). In this study we sought to determine whether the presence of the transcription factors Sox9, AP-2, and [delta]EF1 in chondrocyte nuclei correlate with the cartilage-specific molecules type II collagen and CD-RAP in the ECM. We chose to localize these factors in the growth plate, articular, and meniscal cartilages of the sexually mature mouse because these tissues represent two very different models of in vivo regulation: skeletal growth and maintenance of cartilage tissue.
Materials and Methods
Tissue Preparation
Five 8-week C57BL/6J male mice were used in this study (Washington University School of Medicine animal protocol number 20000244). The mice were purchased from the Jackson Laboratory (Bar Harbor, ME). Knee joint and proximal tibiae were dissected from each mouse and fixed in 4% paraformaldehyde in 0.1 M PBS (pH 7.4) for 16 hr. The samples were decalcified with Formical-2000 (Decal Chemical; Congers, NY) for 24 hr. After dehydration the tissues were embedded in paraffin, sectioned at 5 μm, and mounted on Superfrost/Plus Microscope slides (Fisher Scientific; Pittsburgh, PA).
Antibodies
The human AP-2α and rat [delta]EF1 (ZEB1 R-17) antibodies were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). The antibody recognizing bovine type II collagen was kindly provided by Dr. Michael Cremer (Cremer and Kang 1988) and the antibody recognizing human Sox9 by Dr. Benoit de Crombrugghe (Lefebvre et al. 1997). The human type × collagen polyclonal antibody was a kind gift from Dr. Thomas Schmid (Limoni et al. 1995). The antibody raised against human CD-RAP/MIA was generated by Boehringer–Mannheim (Penzburg, Germany) (Bosserhoff et al. 1997).
Immunohistological Analysis
Immunochemical stainings for CD-RAP/MIA, type II collagen, type × collagen, Sox9, and AP-2α were carried out using a DAKO LSAB Kit (DAKO; Carpinteria, CA). The sections were rehydrated in a serial series of ethanol and immersed in 3% hydrogen peroxide for 15 min to quench endogenous peroxidase activity. The tissue sections were digested with 1% hyaluronidase for 45 min at 37C. The primary antibody was applied and incubated for 45 min at room temperature (RT). Antibodies against human CD-RAP/MIA, bovine type II collagen, human type × collagen, human Sox9, and human AP-2α were used at dilutions of 1:500, 1:100, 1:500, 1:30, and 1:100, respectively. After linking with the second antibody for 30 min at RT, the slides were incubated in streptoavidin solution for 30 min at RT. The antigen site was visualized by 3-amino-9-ethylcarbazole, a chromogen resulting in a reddish-brown precipitate. Immunostaining for CD-RAP included a 20-min incubation in 2% β-mercaptoethanol in PBS for unmasking the epitope before primary antibody application. Immunostaining for [delta]EF1 was performed using the Elite PK-6101 Rabbit IgG ABC Kit (Vector; Burlingame, CA). Tissue sections were processed as above except that the hyaluronidase digestion was replaced by a 15-min incubation in Triton X-100 (1%) before overnight incubation with primary antibody (1:100). Sections were then counterstained with hematoxylin before mounting. The proliferating cell nuclear antigen (PCNA) was detected using a ZYMED PCNA Staining Kit (ZYMED Laboratories; San Francisco, CA) to identify proliferating chondrocytes (Hall et al. 1990). The histological sections were also examined by staining with toluidine blue and with hematoxylin and eosin.
Results
Growth Plate
Hematoxylin–eosin staining of the proximal tibiae from an 8-week postnatal mouse revealed a typical growth plate containing cartilage cells characteristic of various stages of development: the small chondrocytes of the less mature proliferation zone (p), active mature chondrocytes (m), and enlarged chondrocytes of the upper (uh) and lower (lh) hypertrophic zones (Figure 1A). PCNA, a marker of cell division, was detected in both the proliferation and maturation zones, verifying the high division rate of the cells in these areas (Figure 1B). PCNA was not detected in uh or lh chondrocytes. Type II collagen was detected uniformly throughout the growth plate from the zones of proliferation to hypertrophy (Figure 1D). The distribution of type II collagen paralleled that of toluidine blue, a general indicator of negatively charged proteoglycans. The distribution of CD-RAP throughout the growth plate was similar to that of type II collagen. However, it was particularly intense in the pericellular and interterritorial matrices of chondrocytes in the proliferating and resting zones (Figure 1E). The immunostaining for type × collagen was strictly limited to the enlarged chondrocytes typical of hypertrophic cartilage (Schmid and Linsenmayer 1985) (Figure 1F).
Biochemical studies have shown a positive linear relationship between the presence of Sox9 and CD-RAP promoter activity (Xie et al. 1999). Immunolocalization of Sox9 was consistent with positive CD-RAP expression; Sox9 was detected in the nuclei of proliferating chondrocytes and appeared to increase with maturity (Figure 1G). However, no signal for Sox9 was detected in the nuclei of lh chondrocytes (arrowheads), suggesting a loss of Sox9 activity in these cells. In contrast, AP-2 was detected throughout the growth plate and appeared to increase in intensity in uh and lh chondrocytes (Figure 1H). Although there was some cytoplasmic staining in mature and proliferating chondrocytes, there appeared to be no nuclear staining for AP-2 in these cells. Some cells in bone marrow also showed immunoreactivity for AP-2 (not shown). Like high levels of AP-2, [delta]EF1 has also been demonstrated to negatively regulate the CD-RAP promoter (Li and Sandell 2002) and type II collagen (Murray et al. 2000). Immunolocalization of [delta]EF1 is consistent with these data (Figure 1I, brown staining). Both the amount of protein and the nuclear localization appeared to increase with hypertrophy of chondrocytes in which CD-RAP protein appeared to diminish. [Delta]EF1 was also localized in the osteoblasts, bone marrow, and surrounding muscle tissue (data not shown).
Articular and Meniscal Cartilage
The signals for all ECM proteins found in the articular and meniscal cartilages were significantly lower than those found in the growth plate of the same sections (Figures 2A–2D). This likely reflects the age of the matrix and its relative inaccessibility to antibodies, in contrast to newly synthesized matrix that has less crosslinking and is more easily detected. The articular (a) and meniscal (m) cartilages of the joint of the proximal tibiae in an 8-week mouse stained with toluidine blue is shown in Figure 2E. Even though the levels of type II collagen appeared to be less than that in the growth plate (Figures 2B and 2G), the distribution was coincident with toluidine blue (Figures 2A and 2E). We have previously shown that CD-RAP protein can be detected intracellularly in the articular surface and meniscus of an 8-week mouse (Xie et al. 2000). Using dithiothreitol to aid in matrix detection, here we show that CD-RAP deposition in both the articular and meniscal cartilages is very low (Figure 2I). Type X collagen was localized around the enlarged chondrocytes adjacent to the subchondral bone (Figure 2K).
A faint signal for Sox9 was identified in the nuclei of chondrocytes of the articular and meniscal cartilages (Figure 2F). Like type II collagen and CD-RAP, the levels of Sox9 in these chondrocytes were significantly lower than the growth plate. In contrast, high levels of the transacting factors AP-2 and [delta]EF1 were detected in the nuclei of chondrocytes in both these same areas (Figures 2H and 2J), similar to the intensity observed in hypertrophic chondrocytes (Figures 1H and 1I).
Discussion
The localization of cartilage-specific proteins (type II and × collagens and CD-RAP), DNA transcription factors (Sox9, AP-2, and [delta]EF1) and a cell proliferation marker (PCNA) were examined in single sections containing adult murine growth plate, articular, and meniscal cartilages. Consistent with previous mRNA data, the detection of type II collagen in the growth plate was higher than either the articular or meniscal cartilages and correlated well with levels of Sox9 protein (Sakano et al. 1999). This same correlation was observed between CD-RAP and Sox9. Very low levels of CD-RAP and Sox9 were detected in the articular and meniscal cartilages but relatively high levels of both proteins were found in the growth plate. In developing mouse embryos Sox9 mRNA is colocalized with type II collagen and is necessary for chondrogenic differentiation (Ng et al. 1997; Bi et al. 1999). In the postnatal mouse, both type II collagen and Sox9 message levels are coordinately downregulated in the mouse articular cartilage upon maturity (Salminen et al. 2001). In vitro, both type II collagen and CD-RAP expression increase in a dose-dependent manner with Sox9 (Lefebvre et al. 1997; Xie et al. 1999). The results from this study in mature mice are consistent with these in vivo and in vitro data. Levels of Sox9 also appeared to correlate with PCNA, being highest in the proliferative chondrocytes of growth plate and relatively low at the articular surface. This positive correlation of Sox9 expression and proliferation was also observed in our recent studies on fracture healing (Sakano et al. 1999), where Sox9 expression preceded expression of type II collagen and CD-RAP. Although the presence of Sox9 has been implied to imbue function (Ng et al. 1997; Bi et al. 1999), de Crombrugghe and colleagues have shown that phosphorylation of Sox9 significantly increases its activity (Huang et al. 2000). We were not able to address whether the Sox9 detected in this study was indeed active.
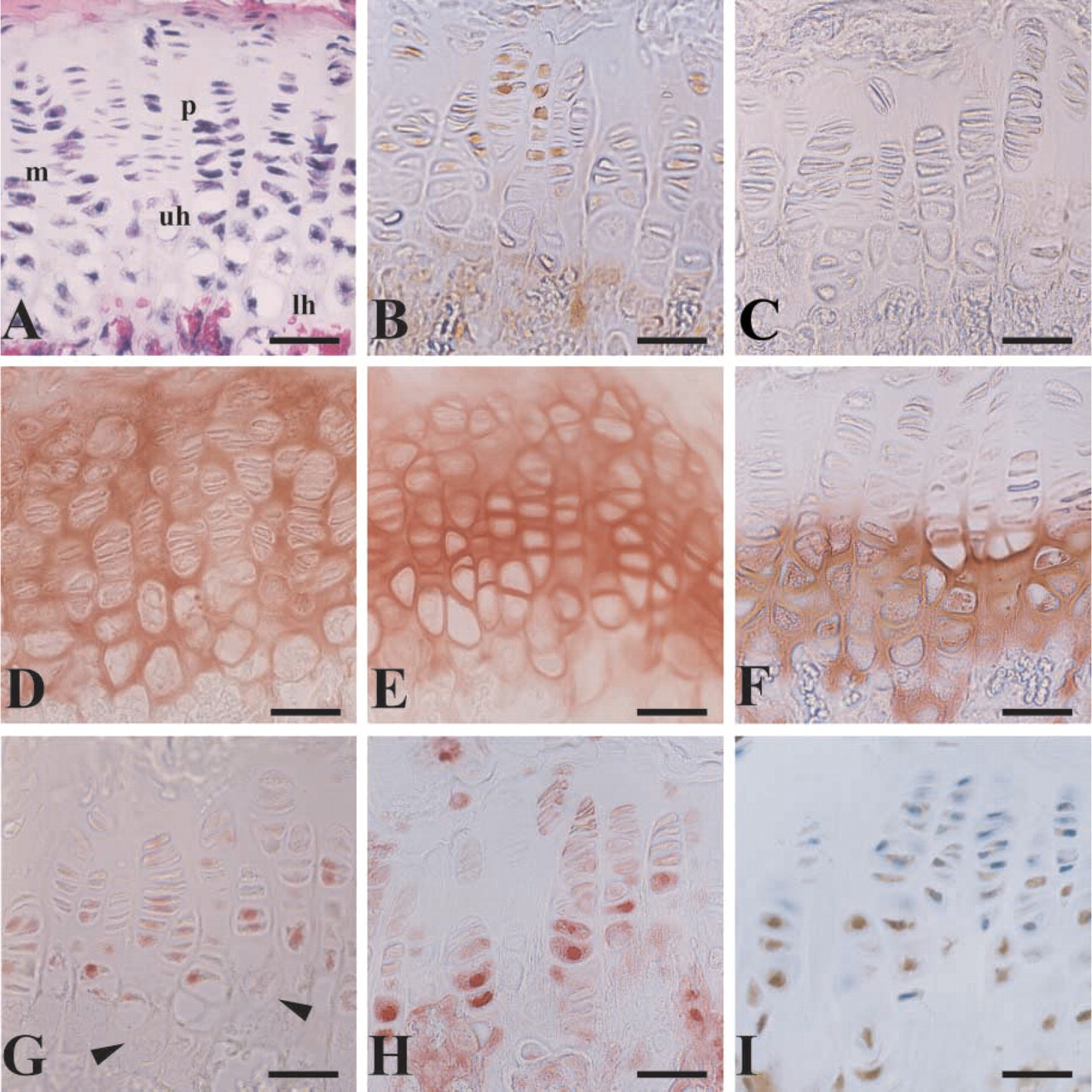
Photomicrographs showing a growth plate in an 8-week mature male mouse. Knee joint and proximal tibiae were fixed, paraffinembedded, and processed for histology with (
An inverse correlation was observed between the localization of the transcription factors AP-2 and [delta]EF1 in nuclei, and CD-RAP and type II collagen in the matrix. Our in vitro studies on CD-RAP gene regulation indicate that both AP-2 (Xie et al. 1998) and [delta]EF1 (Li and Sandell 2002) can suppress promoter activity. We have also found that high levels of AP-2 can suppress type II collagen promoter activity (Purple et al. 2001). Murray et al. (2000) have demonstrated that [delta]EF1 is inversely correlated with type II collagen mRNA and can suppress its promoter activity in chondrocytes. Both [delta]EF1 (Takagi et al. 1998) and AP-2 (Schorle et al. 1996; Zhang et al. 1996) null mice exhibit profound defects in skeletal development and therefore these factors are likely to be important for regulating bone and cartilage differentiation. [Delta]EF1 has also been shown to inhibit hematopoiesis (Takagi et al. 1998; Postigo and Dean 1999) and myogenic (Postigo and Dean 1997) differentiation. Therefore, the lower amounts of type II collagen and CD-RAP detected in the articular and meniscal cartilages may be directly related to the high levels of these transcription factors detected in these same tissues.
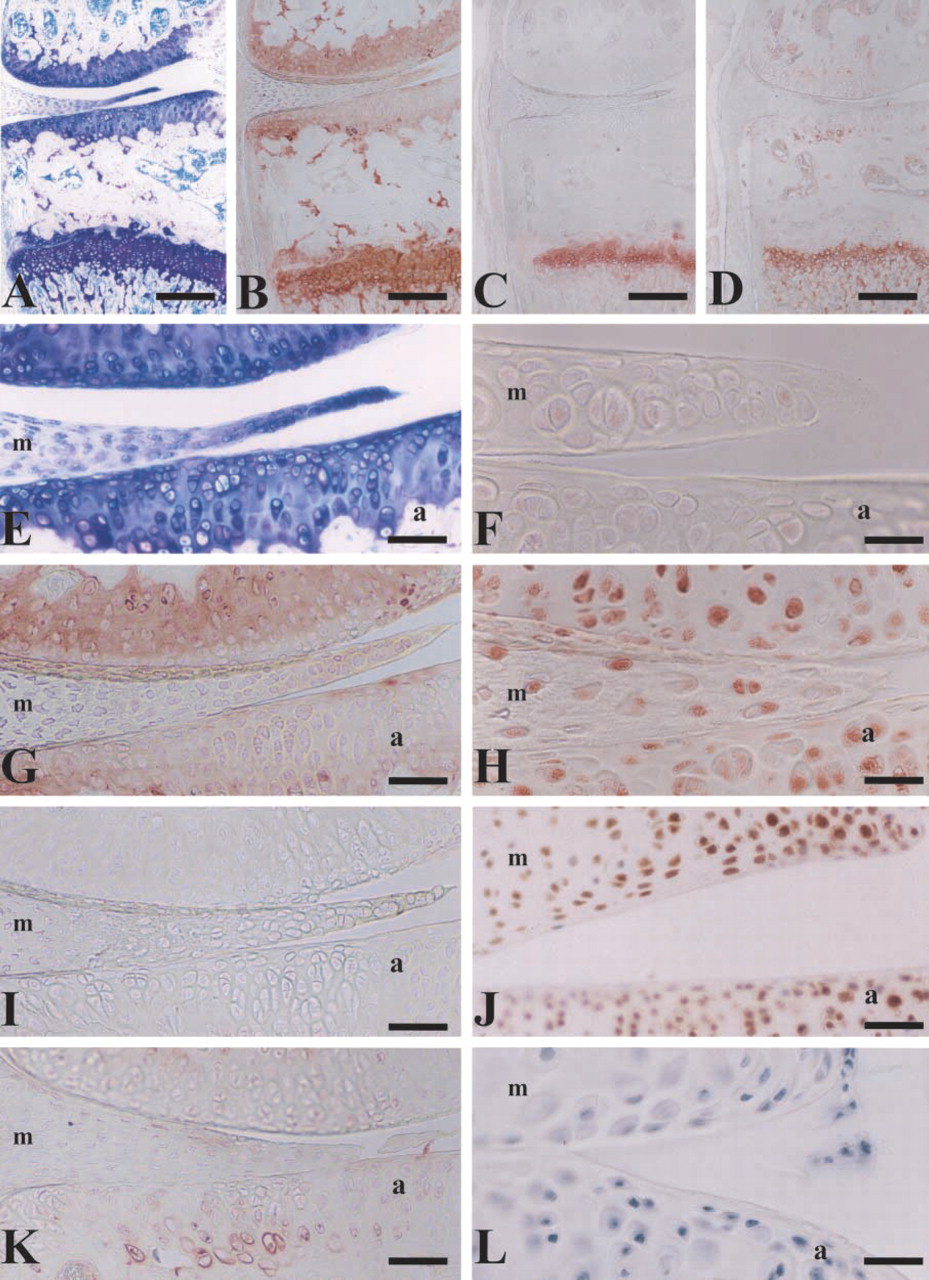
Photomicrographs showing the epiphysis of an 8-week mature male mouse. Knee joint and proximal tibae were fixed, paraffinembedded, and processed for histology with (
The fact that CD-RAP was discovered as a gene downregulated by retinoic acid (Dietz and Sandell 1996) and that AP-2 is upregulated by retinoic acid (Luscher et al. 1989; Oulad–Abdelghani et al. 1996) further supports the hypothesis that AP-2 may play a role as a suppressor of CD-RAP and may act as an effector of retinoic acid during cartilage differentiation. Indeed, retinoic acid may play a role in the transition from the mature to the hypertrophic chondrocyte phenotype (Koyama et al. 1999) because it is increased in hypertrophic chondrocytes. Removal of the AP-2 site in the CD-RAP promoter attenuates the suppression of CD-RAP promoter activity by retinoic acid (Xie et al. 1998). Recently, retinoic acid has also been shown to downregulate Sox9 mRNA and protein levels in chondrocytes in vitro (Sekiya et al. 2001). Taken together with the in vivo localization in this study, it appears that [delta]EF1 and AP-2 may play essential roles in downregulating cartilage matrix genes that are essential for development of cartilage but which are not abundantly synthesized in the adult articular surface or in the terminally differentiated hypertrophic state.
In this study we also examined the distribution of type × collagen in the knee joint. The presence of type × collagen correlated positively with AP-2 and [delta]EF1 in the hypertrophic chondrocytes of the growth plate. However, both of these factors were also intensely expressed throughout the articular surface and meniscus, whereas type × collagen was virtually undetectable in these same areas. This discrepancy between the presence of AP-2 and [delta]EF1 and type × collagen suggests that the expression of type × collagen is not likely to be directly regulated by either factor and is regulated differentially from type II collagen and CD-RAP. This is consistent with previous studies in chick chondrocytes that show differential regulation between types II and × collagens in response to parathyroid hormone-related protein (PTHrP) (O'Keefe et al. 1997). The transcription factor Runx2/Cbfa1 is a likely candidate because it increases type × collagen promoter activity (Leboy et al. 2001) in vitro and is essential for the terminal differentiation of chondrocytes to hypertrophy in several different transgenic mouse models (Inada et al. 1999; Takeda et al. 2001; Ueta et al. 2001).
In summary, the presence of the transacting factors Sox9, AP-2, and [delta]EF1 in the nuclei of chondrocytes is directly correlated with the transcriptional effects observed in vitro (Lefebvre and de Crombrugghe 1998; Xie et al. 1998,1999; Purple et al. 2001; Li and Sandell 2002). That is, increased levels of Sox9 are correlated with increased levels of CD-RAP and type II collagen production, whereas AP-2 and [delta]EF1 are negatively correlated with both CD-RAP and type II collagen expression. The three transcription factors bind to independent sites on DNA and likely work in concert to control the appropriate level of chondrocyte matrix gene expression necessary for both differentiation and maintenance of cartilage tissues. These results support the hypothesis that the chondrocyte phenotype is regulated both positively and negatively, a concept that may prove clinically relevant to metabolic control of chondrocyte gene expression during development and repair processes.
Footnotes
Acknowledgements
Supported by NIH grants R01AR36994 and R01AR45550.