
Abstract
Type II and III fibrillar collagens were localized by immunogold electron microscopy in resin sections of human femoral articular cartilage taken from the upper radial zone in specimens from patients with osteoarthritis. Tissue samples stabilized by high-pressure cryofixation were processed by freeze-substitution, either in acetone containing osmium or in methanol without chemical fixatives, before embedding in epoxy or Lowicryl resin, respectively. Ultrastructural preservation was superior with osmium-acetone, although it was not possible to localize collagens by this method. In contrast, in tissue prepared by low-temperature methods without chemical fixation, collagens were successfully localized with mono- or polyclonal antibodies to the helical (Types II and III) and amino-propeptide (Type III procollagen) domains of the molecule. Dual localization using secondary antibodies labeled with 5- or 10-nm gold particles demonstrated the presence of Types II and III collagen associated within single periodic banded fibrils. Collagen fibrils in articular cartilage are understood to be heteropolymers mainly of Types II, IX, and XI collagen. Our observations provide further evidence for the complexity of these assemblies, with the potential for interactions between at least 11 distinct collagen types as well as several non-collagenous components of the extracellular matrix.
Keywords
I
Electron microscopy employing labeled antibodies has played a major role in studies of the distribution of different collagen types and in demonstrating conclusively by multiple immunolocalization their association within single fibrils comprising connective tissue matrices. However, collagen epitopes for specific antibodies appear intolerant to many of the methods in routine use for microscopy, particularly chemical fixation with aldehydes. Consequently, many previous investigations of collagen interactions have resorted to immunoelectron microscopy of chemically disrupted fibrils or fibrils extracted from disrupted matrix (Birk et al. 1988; Mendler et al. 1989). In fact, for the demonstration of collagen epitopes apparently hidden within the major interstitial collagen fibril, such as Type V within Type I fibrils, fibril disruption was necessary for localization of the minor collagen (Birk et al. 1988). In contrast, to investigate the distribution of collagen heterogeneity with respect to selected anatomic sites, it is essential to maintain tissue integrity.
To overcome the difficulties with collagen sensitivity to chemical fixation and to avoid the extractive effects now known to be associated with conventional processing techniques involving aqueous fixation and dehydration (Hunziker 1993), we previously employed low-temperature tissue preparation to immunolocalize collagen Type III in resin sections of intact human articular cartilage (Young et al. 1995). This method, involving cryofixation at high pressure, freeze-substitution as a dehydration step, and low-temperature resin embedding, was modified from that used previously to demonstrate considerably improved ultrastructural preservation in cartilage tissue (Hunziker et al. 1984, 1996; Hunziker and Schenk 1984; Keene and McDonald 1993). In the absence of aldehyde fixative exposure, collagen Type III was detected in association with the large interstitial fibrils of the cartilage upper mid-zone. This result confirmed earlier biochemical and histochemical evidence for Type III collagen in osteoarthritic and normal cartilage (Adam and Deyl 1983; Wotton and Duance 1994) and suggested a unique association between Type III and the major collage-nous constituent of the tissue, Type II, already known to form heterotypic aggregates. Here we present subsequent studies employing a dual immunogold localization technique on cryopreserved tissue that conclusively demonstrates the presence of these two collagen types in single fibrils of human articular cartilage extracellular matrix.
Materials and Methods
Articular cartilage was obtained from the femoral heads of patients undergoing hip replacement surgery for late-stage osteoarthritis. Samples were selected from areas of unfibrillated intact tissue and were processed for electron microscopy.
Cryofixation-High-pressure Freezing (HPF)
Full-thickness slices of cartilage were taken with a scalpel and stored for up to 16 hr, either in Eagle's MEM + 5% fetal calf serum at 37C or wrapped tightly in plastic film within a sterile vial at 4C to reduce changes in tissue hydration before cryofixation. Cylindrical full-thickness plugs of cartilage 1.7 mm in diameter were first prepared using a microtrephine. Slices approximately 0.1-0.2 mm thick and corresponding to the upper radial zone of the cartilage were then cut from the plugs with a scalpel and immersed in 1-hexadecene. These tissue discs, bathed in 1-hexadecene and sandwiched between two aluminum planchettes, were rapidly cryofixed at 2100 bar pressure in a Baltec 010 HPM machine. Frozen samples were maintained in liquid nitrogen until subsequent processing by freeze-substitution at −90C in a Reichert AFS system (Leica; Milton Keynes, UK), using one of two different methods.
Freeze-substitution with Chemical Fixation Followed by Conventional Epoxy Resin Embedding
Samples were freeze-substituted at −90C for 48 hr in acetone (99.9%, HPLC grade: Sigma-Aldrich, Gillingham, UK), containing 2% w/v osmium tetroxide (Agar Scientific; Stansted, UK) over a 4Å molecular sieve. After programmed warming to 20C at 10C/hr, the tissue blocks were infiltrated with 1:3, 1:1, 3:1 mixtures of Araldite resin:acetone, several changes of 100% resin, and then polymerized in fresh resin at 60C.
Freeze-substitution Without Fixatives Followed by Resin Embedding at Low Temperature
After freeze-substitution at −90C in pure methanol for 24 hr (99.8% anhydrous: Aldrich), the temperature was raised at 10C/hr to −50C. Samples were then infiltrated with Lowicryl HM20 resin (Agar Scientific), through 1:3, 1:1, and 3:1 resin:methanol mixtures for 1 hr each, several changes in 100% resin, and polymerized by UV light. Polymerization was carried out at −50C for 2 days and at room temperature for an additional 2 days.
Antibody Labeling
Ultrathin sections were cut from resin blocks on a Reichert Ultramicrotome equipped with a diamond knife and collected on collodion-coated nickel grids. Before immunostaining, sections were etched by exposure to enzyme solutions: bovine testicular hyaluronidase (Sigma Chemicals, Poole, UK; H-3506, 700 U/ml) and Pronase (Sigma Chemicals; Protease XIV, P5147, 2 U/ml), both for 1 hr at 37C.
The sections were then incubated on droplets of primary antibodies diluted in PBS/1% bovine serum albumin (BSA): (a) a mouse monoclonal antibody to Type II collagen (CII/ CI). This antibody was developed by R. Holmdahl and K. Rubin and is available from the Developmental Studies Hybridoma Bank maintained by the University of Iowa; (b) a polyclonal antibody raised in goat to the pepsin-extracted triple-helical domain of human Type III collagen; (c) a rabbit polyclonal antibody to pN (propeptide) collagen Type III. These antibodies are the same as those used previously (Aigner et al. 1993; Young et al. 1995) and have been extensively characterized by ELISA and Western blotting to exclude crossreactivity to other collagen types and fragments (Nowack et al. 1976; Wotton and Duance 1994). In control experiments, sections were incubated with PBS/1% BSA without primary antibody. In addition, to test for nonspecific binding of rabbit and goat antibodies to human collagens, some sections were incubated with antibodies to cytoskeletal proteins, rabbit anti-tropomyosin and goat anti-vimentin, at dilutions comparable to those of anti-collagen antibodies.
After 2-3-hr incubations and washing with several changes of PBS/1% BSA, the sections were transferred to secondary antibodies labeled with colloidal gold particles, either goat anti-mouse, rabbit anti-goat, or goat anti-rabbit IgG, as appropriate (Amersham International, Poole, UK, or Biocell, Cardiff, UK). Secondary antibodies were preabsorbed to exclude any cross-species reactivity and were used at a dilution of 1:20 in PBS/1% BSA.
In double-labeling experiments to localize collagen Types II and III simultaneously, the first primary and appropriate secondary antibody, followed by the second primary and secondary antibodies, were applied in sequence, with copious washings between incubations. In these experiments, secondary antibodies were employed conjugated to gold particles of different sizes with the 5-nm smaller probe applied before the 10-nm conjugate.
After immunostaining and washing steps, sections were fixed briefly by floating grids on droplets of 2% glutaraldehyde in PBS, followed by final washes in boiled Millipore-filtered distilled water. The sections were then contrasted in aqueous uranyl acetate and lead citrate for 10 and 5 min, respectively, before examination in a Philips 400 or 208 transmission electron microscope at an accelerating voltage of 80 Kv.
Results
HPF-Acetone-Osmium Freeze-substitution-Araldite Embedding
Collagen fibrils in the upper radial zone of articular cartilage exhibited a wide range of diameters and orientations (Figure 1), occasionally undergoing abrupt bends or twists along their longitudinal axes. Finer filamentous elements were also identified scattered among the major interstitial fibrils. In transverse section, fibrils often had an electron-lucent center, a more intensely contrasted perimeter, and were surrounded by an electron-lucent halo concentric to the fibril border. Fibril centers sometimes appeared to be less well embedded than their peripheries, although this was not evidenced in fibrils sectioned longitudinally. Some fibrils with irregular transverse section profile nevertheless were surrounded by a halo with regular smooth contours (Figure 1). This lucent perifibrillar feature was also evident in fibrils viewed in longitudinal section. The associated interfibrillar matrix in these sections displayed a uniform coarse granularity, which terminated abruptly at its interface with the perifibrillar halo.
Attempts to immunolocalize individual collagen types with specific antibodies on sections of HPF tissue freeze-substituted in acetone-osmium were unsuccessful. However, immunogold detection of the proteoglycans keratan and chondroitin sulfate in the interfibrillar cartilage matrix of these same preparations was readily achieved (not shown).
HPF-Methanol Freeze-substitution Without Fixatives-Low-temperature Embedding
As expected, electron contrast was reduced in Lowicryl resin sections of cartilage prepared without exposure to osmium tetroxide, even after prolonged section staining with uranyl acetate and lead citrate. The collagenous cartilage matrix was well preserved, although little nonfibrillar matrix structure could be observed and some tissue shrinkage was evident. Ultrastructural features, such as the electron-lucent perifibrillar halos and regions of reduced contrast at the centers of transverse-sectioned collagen fibrils, were absent from tissue embedded at low temperature.
In all cases in which sections were exposed to buffer solutions without anti-collagen antibodies or in which antibodies to vimentin or tropomyosin were used, immunogold labeling either was completely absent or was located in very small numbers after prolonged searching (Figure 2). However, unlike in the osmium-treated tissue, localization of collagens was possible (Figures 3-8).
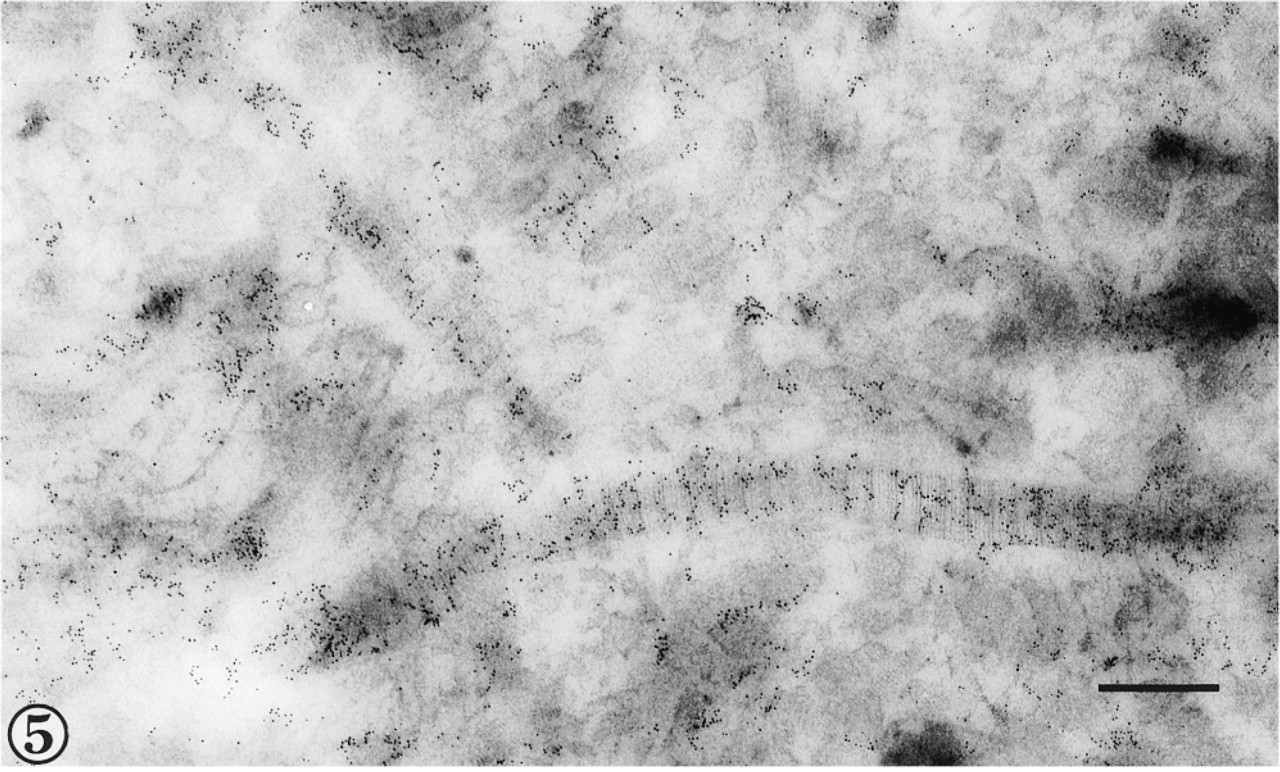
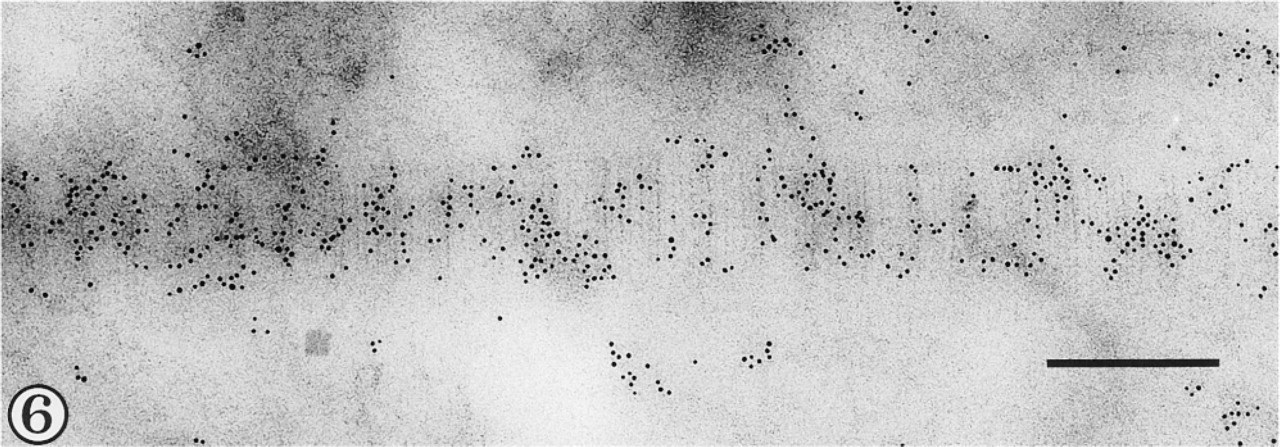
Collagen Type III could be detected both with an antibody against the N-propeptide and with one reacting with the triple-helical domain of the molecule (Figures 3 and 4). Immunogold labeling was more abundant with the first antibody, which appeared densely scattered over regions containing finer fibrils but which also labeled associated fibrils of larger diameter (Figure 3). The N-propeptide antibody labeled some sites apparently unattached to fibrillar material, particularly where aggregations of fine fibrils were located. The antibody against the helical domain of collagen Type III did not appear to label fine fibrils as densely as the larger collagen fibrils. Both antibodies labeled fibrils sectioned transversely to a lesser extent than those displayed in longitudinal section. Collagen Type II immunolocalization was also evident on fibrils of widely different diameters. Immunogold labeling was best observed in larger-diameter fibrils viewed in longitudinal section (Figures 5 and 6).
Sections exposed sequentially to antibodies to Type II and Type III collagens, followed, after each, by the appropriate gold-conjugated secondary antibody, clearly showed the presence of both collagen types within individual collagen fibrils (Figures 7 and 8). Co-localization of Type II and Type III collagens was most evident on large-diameter fibrils sectioned longitudinally and was not detected on the finer fibrils in the matrix. Experiments applying the two anti-collagen primary antibodies in different sequence yielded similar staining patterns. In addition, Type II:Type III collagen co-localization was demonstrated with either of the anti-Type III collagen antibodies, with similar results.
Discussion
The collagenous fibrillar matrix of mammalian articular cartilage is composed primarily of Type II collagen, with at least ten additional collagens, including Types VI, IX, X, and XI, present as minor components (Duance et al. 1999). The functional role of these minor collagens, the nature of their organization within the Type II meshwork, and their influence on development, aging, and disease processes are important aspects about which relatively little is yet known. Earlier views of different collagen types as tissue-specific entities are no longer tenable, and with increasing knowledge of the domain structure of the different collagen species, their potential for interaction both with each other and with other components of the extracellular matrix is now becoming evident.
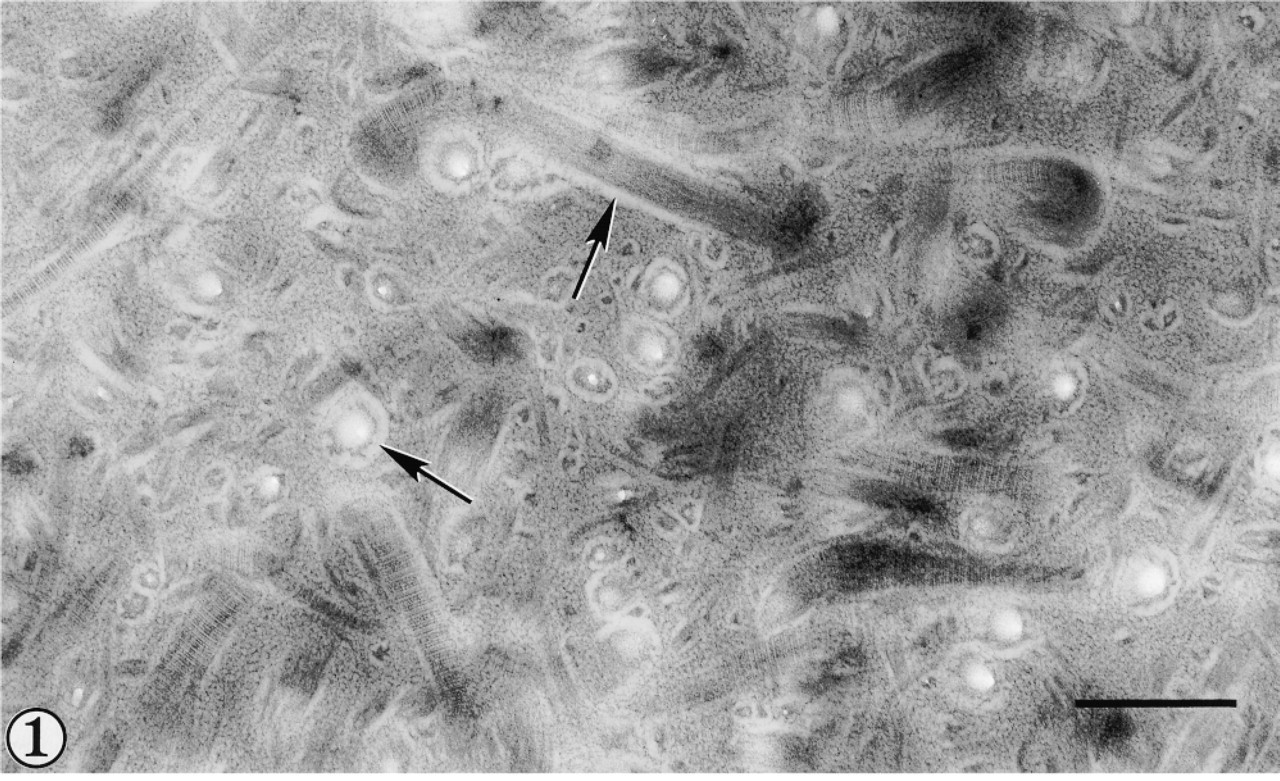
Interterritorial matrix of human articular cartilage from the upper radial zone prepared by high-pressure freezing, freeze-substitution in acetone-osmium, and embedding in epoxy resin. Collagen fibrils in longitudinal and transverse section exhibit a perifibrillar electron-lucent halo (arrows). Bar = 500 nm.
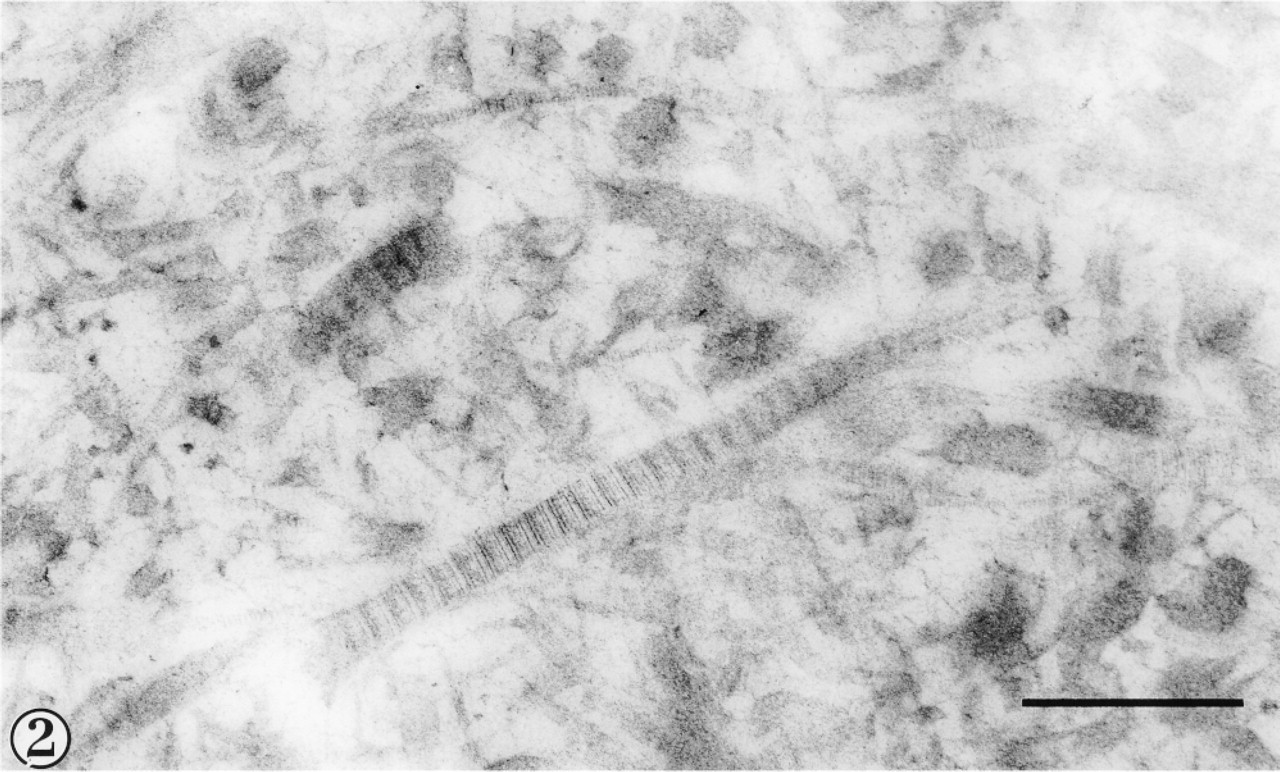
Human cartilage after high-pressure freezing, freeze-substitution in methanol, and Lowicryl resin embedding at low temperature. Control section after exposure to anti-vimentin primary antibody followed by colloidal gold-conjugated secondary antibody. Few gold particles are evident. Bar = 500 nm.
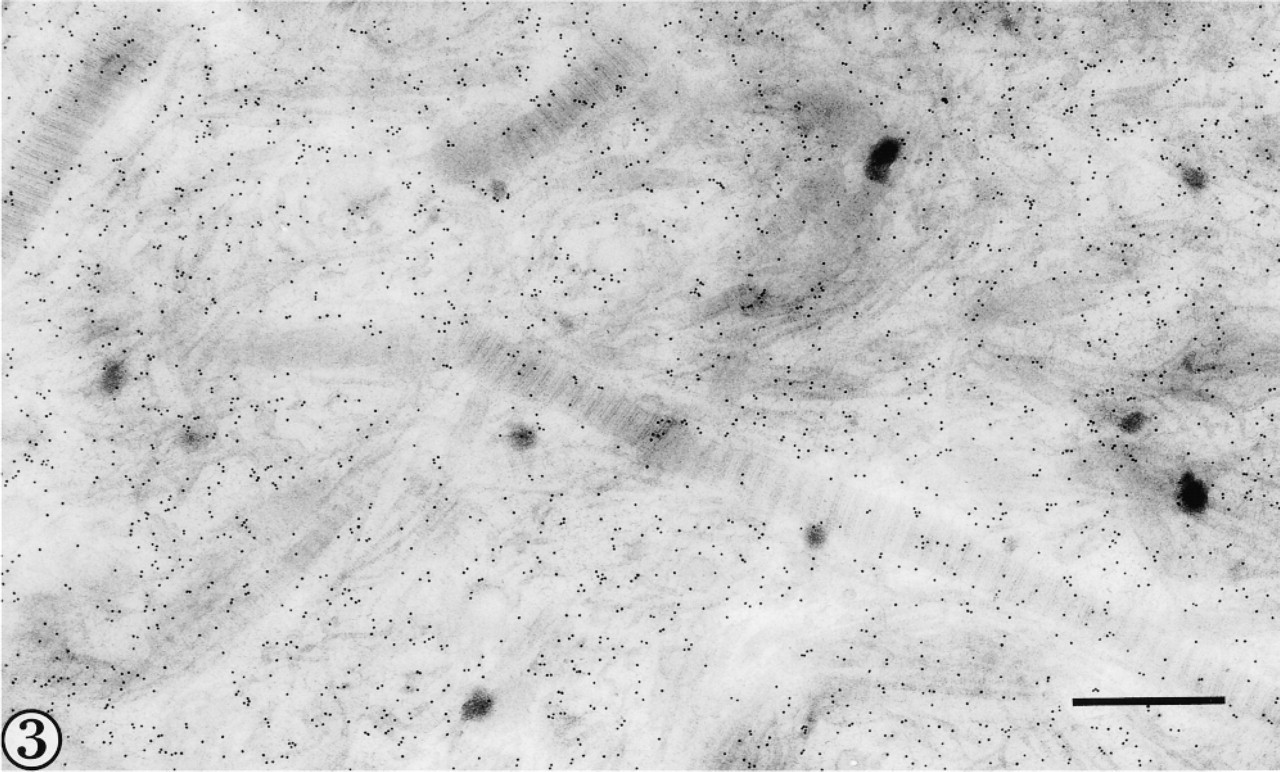
Type III collagen is localized in fine and large-diameter fibrils using an antibody to N-propeptide of collagen Type III. Bar = 500 nm.
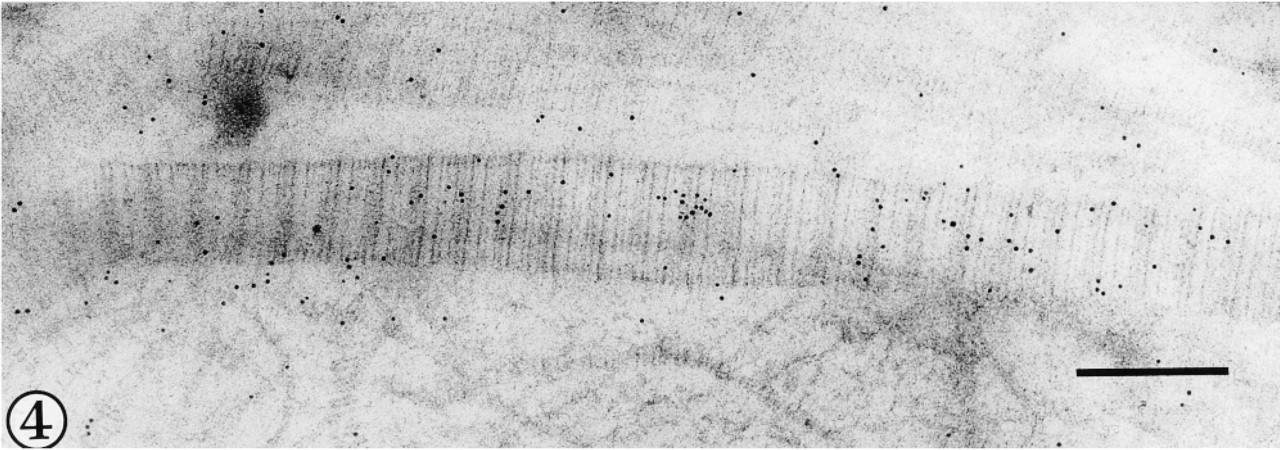
Gold particles indicate positive labeling for Type III collagen in a large diameter fibril after exposure to antibody to the helical domain of the molecule. Bar = 200 nm.
This study is in accord with earlier reports of Type III collagen present as a minor component of articular cartilage (Wotton and Duance 1994; Young et al. 1995). Type III collagen has usually been found associated with Type I collagen (Fleischmajer et al. 1990). However, several examples of coordinated synthesis of Types II and III collagen have been reported, including cartilage canals of human thyroid cartilage (Claassen et al. 1996), during development in rabbit knee articular cartilage (Bland and Ashhurst 1996), and in pathological conditions of the temporomandibular joint (Salo and Raustia 1995). None of these studies addressed the possibility of Type II and Type III collagen assembly into single matrix structures, although the potential for interaction between these collagens was highlighted by biochemical isolation of Type II-Type III collagen crosslinks from normal adult human articular cartilage (Wu et al. 1996). Here we have demonstrated by high-resolution immunodetection that Type III does indeed associate with Type II, and thus potentially also with collagen Types IX and XI, in complex heterotypic fibrils in the cartilage extracellular matrix.
Type II collagen is localized in large-diameter fibrils viewed in longitudinal and oblique section. Bar = 300 nm.
Gold particles illustrate dense labeling for Type II collagen epitope, but without obvious periodicity, in a fibril in longitudinal section. Bar = 200 nm.
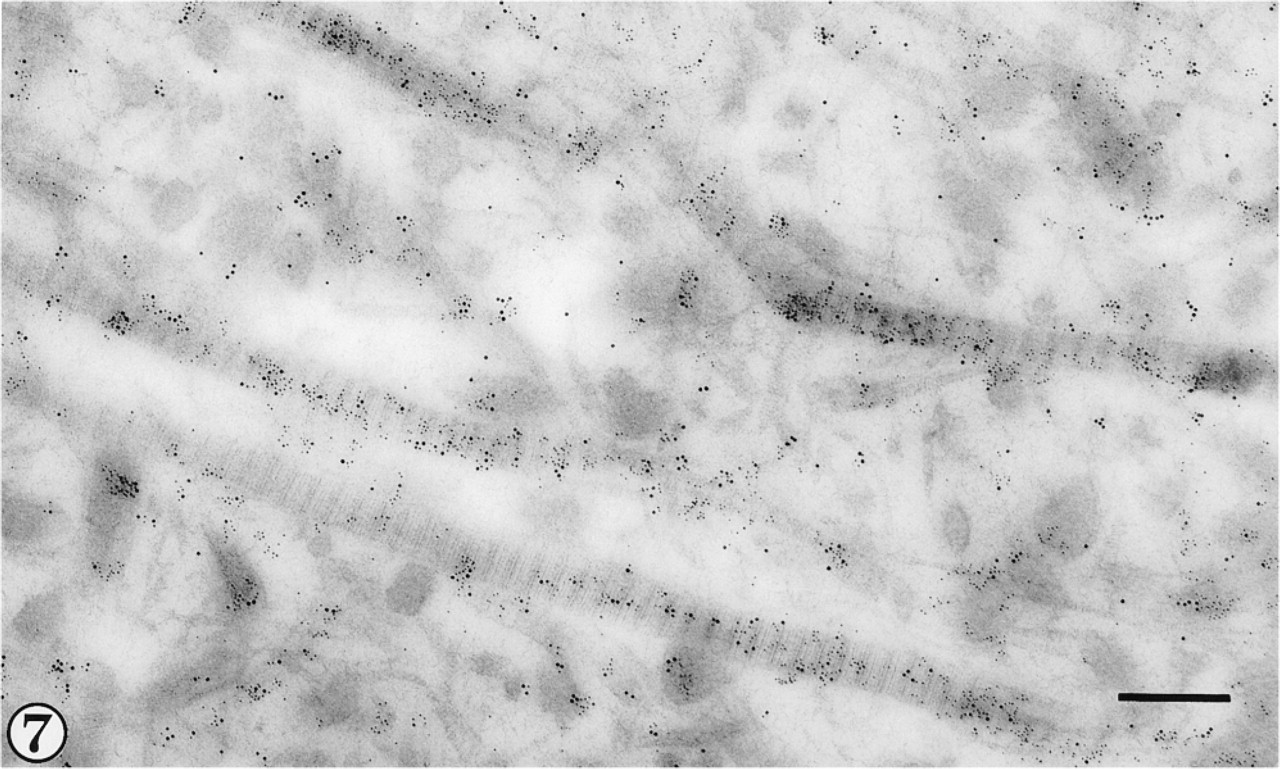
After exposure to anti-Type II and anti-(helical domain) Type III primary antibodies, Types II and III collagen are identified, respectively, by 5- and 10-nm gold particles in large-diameter fibrils of the upper radial zone. Bar = 300 nm.
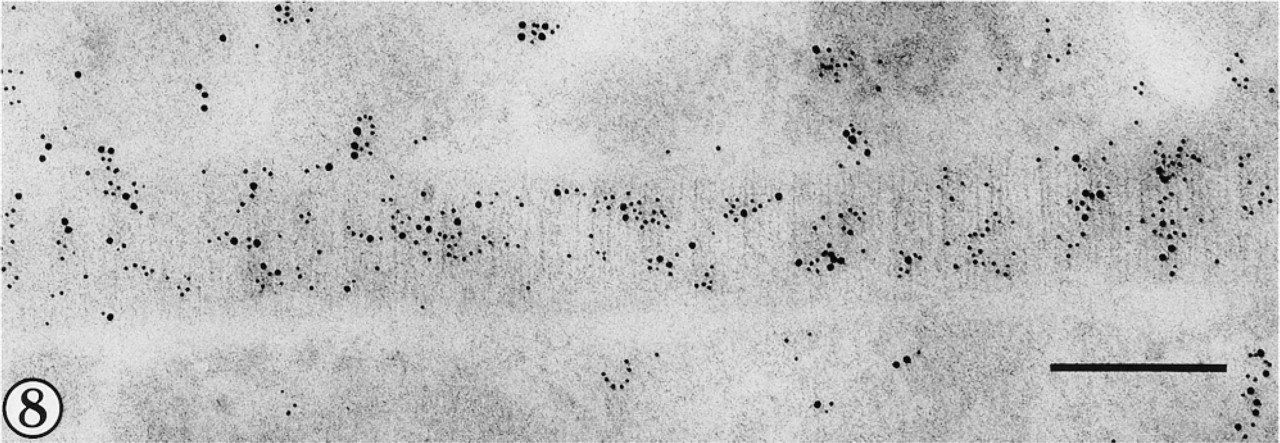
Collagen Type II is indicated by 5-nm and Type III by 10-nm gold particles in a fibril viewed in longitudinal section. Bar = 200 nm.
Collagen Type III, like collagens I and II, conforms to the classical interstitial molecular structure with 300-nm rod-like tropocollagen molecules produced from procollagen by enzyme-induced cleavage of terminal N- and C-propeptides before their assembly into fibrils in the extracellular matrix. Collagen Type III retains the N-propeptide for some time after fibrillogenesis (Fessler et al. 1981; Fleischmajer et al. 1981).
Type III N-propeptide would be expected to be more abundant in newly formed fibrils of smaller diameter, although some is cleaved to remain free in the interfibrillar matrix for a short time before clearance. This is consistent with our observations of more widespread immunolabeling with the collagen Type III N-propeptide antibody and the lower level of staining in fibrils of larger diameter compared to labeling with antibody against the helical domain of the molecule. Although the mechanisms of organization and assembly of collagen molecules into crossbanded fibrils remain the subject of intensive research (see Kadler et al. 1996), this phenomenon remains incompletely understood even for homotypic fibrils. Even less is known about the sequence of assembly during fibrillogenesis of hybrid collagen polymers. The N- to C-terminal polarity of constituent molecules within the collagen fibril is believed to have a profound influence on fibril growth (Kadler et al. 1996). There may be important differences in molecular organization between Type II and Type I collagen fibrils that influence the surface binding of Type III collagen molecules or molecular aggregates. However, the potential for Type III interaction with these structures may be further influenced by the distribution of other moieties at the fibril surface, such as proteoglycans and additional minor collagens (Hagg et al. 1998). Immunogold localization studies suggesting that Type XII and Type XIV collagens also occupy sites at the Type II fibril surface (Watt et al. 1992) provide additional evidence for the considerable complexity of molecular interactions that are presumed to be involved in maintaining tissue integrity at the fibril perimeter.
A major question remains concerning the significance of Type III collagen in the sequence of fibril assembly in cartilage and its relationship to the pathogenesis of osteoarthritis. At present, it is unknown whether cartilage fibrils incorporate Type III during synthesis of new fibrils or whether this collagen is assembled onto the surface of existing Type II/Type IX/ Type XI polymers as part of age- or disease-related degradation or repair. Our previous study showed Type III collagen to be present on fibrils in the matrix of normal as well as osteoarthritic human articular cartilage (Young et al. 1995). Increased Type III collagen expression associated with Type II biosynthesis was previously detected in osteoarthritic tissue by Aigner et al. (1993). However, we have not yet confirmed whether fibrils in normal cartilage are also heterotypic for Types II and III collagen, as shown here in diseased cartilage. Further work is required to elucidate the effects of increasing proportions of Type III collagen on fibril stability and thus upon the functional integrity of cartilage extracellular matrix.
Cryofixation, by high-pressure freezing, and freeze-substitution followed by ambient or low-temperature embedding, has now been used by many investigators as a means to prepare cartilage for ultrastructural examination (Hunziker and Schenk 1984; Hunziker et al. 1984, 1996, 1997; Hunziker 1993; Keene and McDonald 1993; Engfeldt et al. 1994; Young et al. 1995; Studer et al. 1995, 1996). These methods have the potential to preserve the native conformation of proteins and other tissue constituents. The problems of extraction and distortion of the hydrated matrix of cartilage connective tissue, which represent serious disadvantages in conventional techniques of aqueous fixation and dehydration at ambient temperature, may thus be overcome.
There have been relatively few reports of immunolocalization of collagens in high-pressure frozen tissues. Although immunolocalization of invertebrate tissue antigens, including cuticular collagen in Arenicola, was improved by the inclusion of osmium in acetone substitution media (McDonald and Morphew 1993; Nicolas et al. 1997), other studies have suggested that freeze-substitution in the presence of chemical fixatives interferes with epitope availability for collagen-specific antibody binding (Hunziker 1993). Freeze-substitution of high-pressure cryofixed tissue, omitting chemical fixatives, followed by low-temperature embedding in Lowicryl resins, is currently the method of choice for immunolocalization of sensitive tissue antigens (Monaghan and Robertson 1990; Usada et al. 1990; Nicolas and Bassot 1993; Kiss and Staehelin 1995; Monaghan et al. 1998).
High-pressure freezing is capable of vitrifying cartilage specimens of up to 150-μm thickness (Studer et al. 1995; Hunziker et al. 1996). By thus avoiding the destructive effects of ice crystal deposition, the native conformation of tissue constituents are believed to be preserved. Under conditions of optimal freezing, the interfibrillar matrix of vitrified cartilage appears finely granular or densely amorphous without any reticular structure (Keene and McDonald 1993; Studer et al. 1995; Hunziker et al. 1997). Morphologically, although no filamentous reticular structure or large segregation artifacts typical of crystalline ice deposition were evident in the present study, the interfibrillar matrix appeared densely granular. Therefore, it appears unlikely that true vitrification was achieved. Even under optimal experimental conditions, vitrification appears to be greatly influenced by the water and proteoglycan composition of cartilage matrix. This was most favorable in the lower radial zone of bovine articular cartilage (Studer et al. 1995), which could be completely vitrified. Specimens for the present study were sampled from the upper radial zone of human cartilage, which may deviate significantly from that considered optimal for vitrification in bovine tissue. In addition, the composition of osteoarthritic cartilage matrix may exhibit increased localized heterogeneity in relation to degradative and reparative processes associated with the disease.
Although it appears that the efficacy of high-pressure freezing for vitrification of bulk tissue samples may well have been overestimated previously, its potential for immunodetection remains underexploited. It therefore continues to offer an unrivaled opportunity for investigations of molecular interactions in connective tissues and of the changes that occur in disease.
Footnotes
Acknowledgment
Supported by a grant from the Arthritis Research Campaign, UK.