
Abstract
Tenascin-C is an extracellular matrix (ECM) glycoprotein expressed in human tissues during organogenesis and in fibrotic and neoplastic processes. We hypothesized that its expression would increase in human lung in neonatal disorders such as infant respiratory distress syndrome (RDS) and bronchopulmonary dysplasia (BPD). Tenascin-C expression was studied by immunohistochemistry (IHC) and mRNA in situ hybridization (ISH). The extent of tenascin-C immunoreactivity was scored as absent (0), low (+), moderate (++), strong (+++), or very strong (++++) separately in different types of pulmonary cells in controls (seven cases), RDS (19 cases), and BPD (12 cases). In controls, tenascin-C expression was low (+) underneath alveolar and bronchiolar epithelium, moderate (++) in intima of veins, and strong (+++) around chondrocytes. In RDS, tenascin-C expression was moderate (++) or strong (+++) underneath both bronchiolar and often detached alveolar epithelium underlying hyaline membranes in the walls of dilated alveoli. In particular, the patients with RDS who survived for 1 day or more had strong expression of tenascin-C within alveolar walls. In patients with BPD, tenascin-C was very strongly (++++) expressed in the remodeled fibrotic alveolar walls underneath regenerative epithelium. Increased expression of tenascin-C mRNA was seen below the alveolar and bronchiolar epithelia in RDS and BPD. The cells in these locations showed α-smooth muscle actin immunoreactivity, suggesting a myofibroblast phenotype. In conclusion, tenascin-C is highly expressed in the walls of alveoli and bronchioli in RDS and BPD, suggesting an association between the expression of this protein and the presence of these disorders.
In 1967, Northway and co-workers described bronchopulmonary dysplasia (BPD) in pre-term infants who developed a chronic lung disease (CLD) after having received assisted ventilation for hyaline membrane disease, i.e., respiratory distress syndrome (RDS) (Northway et al. 1967). Despite many later investigations, the pathogenesis of BPD is still not clearly defined. BPD may be a result of pulmonary immaturity leading first to RDS and then to abnormal repair (Delemos and Coalson 1992). The roles of oxygen and barotrauma have also been studied (Anderson et al. 1973; Rhodes et al. 1975). In recent years, the number of infants with BPD has been increasing because of improved survival among infants with very low birth weights (Hack et al. 1991).
Interactions between epithelial and mesenchymal cells play an important role both in lung development and in fibrotic disorders of the lung. It is now evident that cytokines control these processes. Transforming growth factor-β (TGF-β) is a cytokine that controls the expression of several extracellular matrix (ECM) glycoproteins. It is increased at the site of lung injury in patients with pulmonary fibrosis and BPD (Broekelmann et al. 1991; Toti et al. 1997). Fibronectin is an ECM glycoprotein shown to be more abundant in the lungs of adult pulmonary fibrosis and BPD than in normal lungs (Torikata et al. 1985; Sinkin et al. 1998). Tenascin-C is another ECM glycoprotein that is synthesized during embryonic development, being absent or restricted in most adult tissues and re-expressed in tumors, wound healing, and inflammation (Erick-son 1993; Chiquet-Ehrismann 1995). Its expression is increased in several types of fibrotic adult lung disorders (Kaarteenaho-Wiik et al. 1996), and in a recent study tenascin-C was shown to be one of the most up-regulated genes in mouse pulmonary fibrosis (Kaminski et al. 2000). To our knowledge, no studies on the expression of tenascin-C in RDS and BPD have been published thus far.
The aim of the present study was to investigate the distribution of tenascin-C in RDS and BPD and to compare the expression of tenascin-C in these disorders to control cases of the same gestational ages. Most clinical definitions consider 28 days of age as the beginning of BPD. However, early pathological changes of BPD can be recognized within a few days after birth (Northway 1992). In this study we used a definition based on morphological criteria (Rosan 1975). The immunoreactivity for tenascin-C was scored from absent to very strong in and around different types of pulmonary cells in pre-term infants with RDS and BPD and in control lung. We hypothesized that tenascin-C expression would increase in these disorders comparable to that in fibrotic lung disorders of adults.
Materials and Methods
Patients and Handling of Specimens
Histopathologically typical cases of RDS and BPD were retrieved from the files of the Department of Pathology, Oulu University Hospital. The study protocol was approved by the Ethical Committee of the Medical Faculty of the University of Oulu. The patients were 38 autopsied babies who had died at Oulu University Hospital between 1987 and 1999. Autopsies had been performed within 3 days. A total of 19 patients developed RDS requiring mechanical ventilation after birth, and 12 patients developed BPD after having RDS. The controls (seven cases) included in the study were newborn infants (23–28 weeks of gestational age) who had died within 1 hr after delivery for different reasons without lung disorders. The study group consisted of 19 cases with RDS, i.e., hyaline membrane disease, and 12 cases of BPD with typical morphological findings. BPD was classified on morphological grounds into four stages according to Rosan: stage I, acute (2–4 days); stage II, regenerative (4–8 days); stage III, transitional (8–6 days); and stage IV, chronic (after 16 days) (Rosan 1975). BPD cases were as follows: stage II (four cases), stage III (two cases), and stage IV (six cases). Infants with pneumonia or cardiac abnormalities were excluded. The clinical information was obtained from the patient records. The clinical information on the controls is shown in Table 1 and that on the study groups in Table 2.
Lung tissues from either right or left lung removed at autopsy were fixed in 10% formalin, then dehydrated and embedded in paraffin. Sections of 4 μm were stained with hematoxylin-eosin. The entire material was re-evaluated and one representative tissue block from each case was selected for immunohistochemical (IHC) studies in the entire study material. In 12 cases (four cases with RDS, four cases with BPD, and four control cases) one tissue block was selected for tenascin-C mRNA in situ hybridization (ISH). In these cases the autopsy had been performed within 1 day after death. Some mRNAs have been shown to be stable within a 24-hr period under postmortem conditions enabling application of ISH techniques to postmortem material (Walker and McNicol 1992). To verify the ISH reaction, a positive control previously demonstrated to express signals for tenascin-C mRNA in human lung, such as interstitial pneumonia, was used in the experiments (Pääkkö et al. 2000).
Control casesa
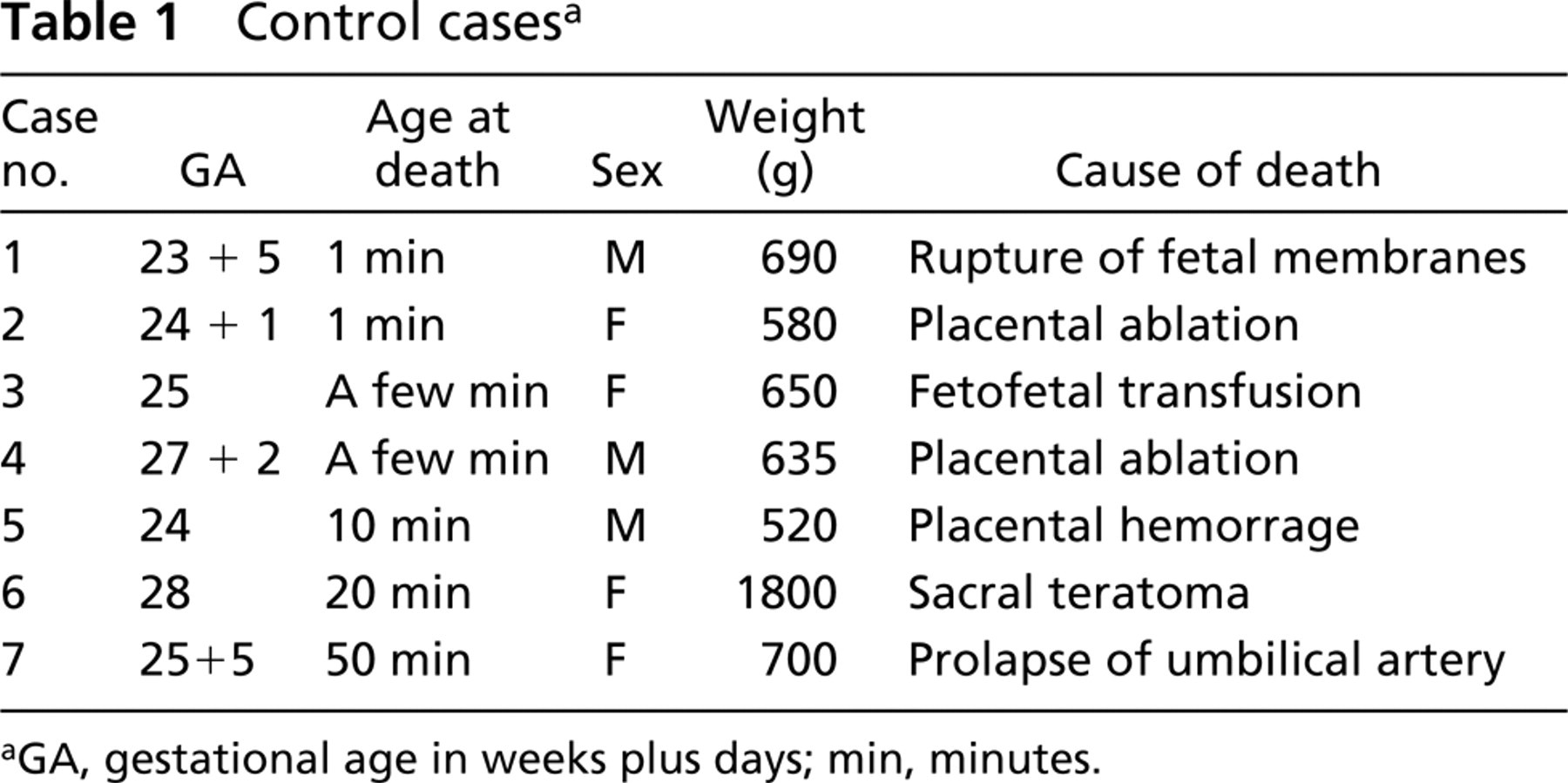
GA, gestational age in weeks plus days; min, minutes.
Newborns with respiratory distress syndrome (RDS) and bronchopulmonary dysplasia (BPD)a
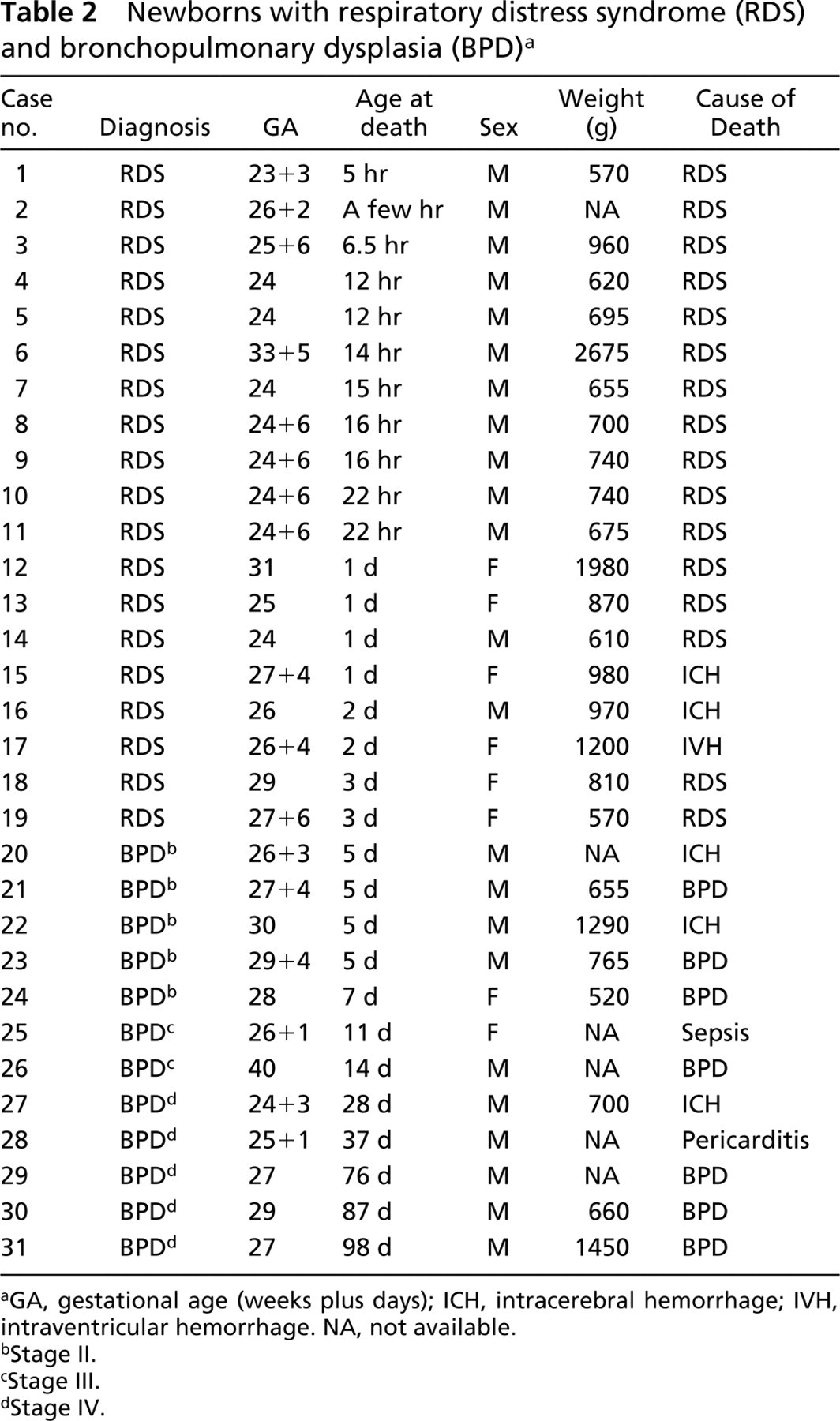
GA, gestational age (weeks plus days); ICH, intracerebral hemorrhage; IVH, intraventricular hemorrhage. NA, not available.
Stage II.
Stage III.
Stage IV.
Anti-tenascin-C Antibody and IHC Staining
A monoclonal antibody (MAb) 143DB7, known to react with the two major isoforms of tenascin-C, was used. The Mab 143DB7 was developed to detect tenascin-C in formaldehyde-fixed tissue and has been characterized in detail elsewhere (Soini et al. 1993). It is available from Locus-Genex (Helsinki, Finland).
Sections 4 μm thick were deparaffinized in xylene and re-hydrated in graded ethanol. Endogenous peroxidase was consumed by incubating the sections in 0.1% hydrogen peroxide in absolute methanol for 20 min. Before immunostaining, the sections were treated with 0.4% pepsin (Merck; Darmstadt, Germany) at 37C for 30 min. For the immuno-staining, the avidin-biotin-peroxidase complex method was used as previously described (Kaarteenaho-Wiik et al. 1996).
MAb 143DB7 at a dilution of 1:1000 of the hybridoma supernatant was used as the primary antibody. The sections were incubated with the primary antibody at 4C overnight, followed by a biotinylated rabbit anti-mouse secondary antibody (at a dilution of 1:300 for 30 min) and the avidin-biotin-peroxidase complex (both from Dakopatts; Glostrup, Denmark). The color was developed with diaminobenzidine. Sections were counterstained with a light hematoxylin stain and mounted with Eukitt (Kindler; Freiburg, Germany). The negative control consisted of substituting the primary antibody with PBS (pH 7.2) or serum isotype control (Zymed Laboratories; San Francisco, CA).
To identify the phenotype of the tenascin-C-expressing cells, the sections were stained with a commercially available antibody against α-smooth muscle actin (Clone 1A4 from Sigma BioSciences; St Louis, MO). The extent of α-smooth muscle actin immunoreactivity was scored as absent (0), low (+), moderate (++), strong (+++), or very strong (++++) in different types of lung cells.
Preparation of Tissue Sections for ISH
Four-μm-thick sections from paraffin-embedded lung biopsies were collected on clean Superfrost Plus glass slides (Erie; Portsmouth, NH), paraffin was removed by xylene, and tissues rehydrated through a graded ethanol series. After three immersions in PBS, pH 7.2, the sections were treated in 0.2 M HCl for 20 min, twice in PBS for 3 min each, followed by proteinase K (100 μg/ml in PBS) treatment for 15 min at 37C. Then tissue sections were transferred into 0.025 M glycine for 30 sec. Tissues were postfixed in 4% paraformaldehyde (Fluka; Buchs, Switzerland) in PBS, pH 7.2 (all solutions made with 0.1% diethyl pyrocarbonate-treated water). After postfixation, tissue sections were transferred into 0.025 M glycine in PBS for 3 min and then acetylated in 0.25% acetic anhydride in 0.1 M triethanolamine (pH 8.0) for 10 min (Rosenfeld et al. 1991). After that, tissue sections were rinsed in 2 × SSC, dehydrated through a graded ethanol series, and air-dried.
Preparation of RNA Probes
A cDNA fragment (bases 814–1316) of the full-length human tenascin-C cDNA (Siri et al. 1991) was synthesized by PCR from the HT-11 subclone kindly provided by Dr. Luciano Zardi (Instituto Nazionale Per la Ricerca Sul Cancro; Genova, Italy) by using the following primers: 5′ CCC TGC AGT GAG GAG CAC GGC ACA 3′ and 5′ TGC CCA TTG ACA CAG CGG CCA TGG 3′.
The 503-bp PCR product containing a specific sequence for tenascin-C was subcloned into a TA vector (TA Cloning Kit; Invitrogen, Carlsbad, CA). Sense and antisense RNA probes were generated from a linearized template by using a riboprobe transcription kit (Promega; Madison, WI) and the probes were labeled with [35S]-UTP (Amersham; Little Chalfont, UK). The radioactively labeled RNA probes were purified by centrifugation through Bio-Gel P-30 columns (Bio-Spin 30; Bio-Rad, Richmond, CA). Each in vitro transcription reaction yielded RNA probes of high specific activity (typically 4.5–6 × 108 dpm/1 μg DNA template and 50–70% incorporation).
In Situ Hybridization
The hybridization mixture contained the [35S]-labeled RNA probe (1.2 × 105 dpm/μl), 50% deionized formamide (Gibco BRL; Rockville, MD), 5 mM dithiothreitol, 500 μg/ml yeast tRNA (Gibco), 2 mg/ml bovine serum albumin (Gibco), and 4 × SSC. The samples were hybridized overnight at 50C while covered with Parafilm (Pääkkö et al. 1992) and then washed three times in 2 × SSC/50% formamide at 52C and 2 × SSC at room temperature (RT), followed by incubation in an RNase solution (100 μg/ml RNase A; Boehringer Mannheim, Mannheim, Germany) in 2 × SSC at 37C for 30 min. The tissue sections were subsequently washed in 2 × SSC/50% formamide at 52C for 5 min and three changes of 2 × SSC at RT, dehydrated sequentially in 70, 80, and 95% ethanol for 1 min each with agitation, and air-dried. Autoradiography was performed by dipping them in Kodak NTB-2 nuclear track emulsion diluted 1:1 with sterile distilled water at 42C. After a weeks' exposure in dark at 4C, the slides were developed in Kodak D-19 developer for 5 min, rinsed in 1% acetic acid in distilled water for 30 sec, fixed in Kodak Agefix for 5 min, rinsed in distilled water, and stained with hematoxylin-eosin.
To evaluate the specificity of the 35S-labeled antisense tenascin-C probes to the tissue sections of lung specimens, control experiments were performed using 35S-labeled sense tenascin-C probes separately for each sample.
The hybridized tissue sections of lung samples were examined by light microscopy and the number of grains over the cells evaluated in general and especially at the locations where tenascin-C immunoreactivity was seen. Cells or cell groups hybridized with the 35S-labeled antisense tenascin probe were considered positive if they contained more grains than corresponding cells and tissue areas that had been hybridized with the 35S-labeled sense tenascin-C probe.
Scoring of Tenascin-C Immunoreactivity and Statistical Analysis
The extent of tenascin-C immunoreactivity was scored as absent (0), low (+), moderate (++), strong (+++), or very strong (++++) separately in and around different types of pulmonary cells in controls and study groups. In each case, the entire section of lung tissue (roughly 1–2 cm2) was evaluated. Two investigators (RKW and PP) performed the analysis of tenascin-C immunoreactivity independently. The inter-observer repeatability was excellent (
The significance of the associations was determined using Fisher's exact probability test designed for small sample groups.
Results
Controls
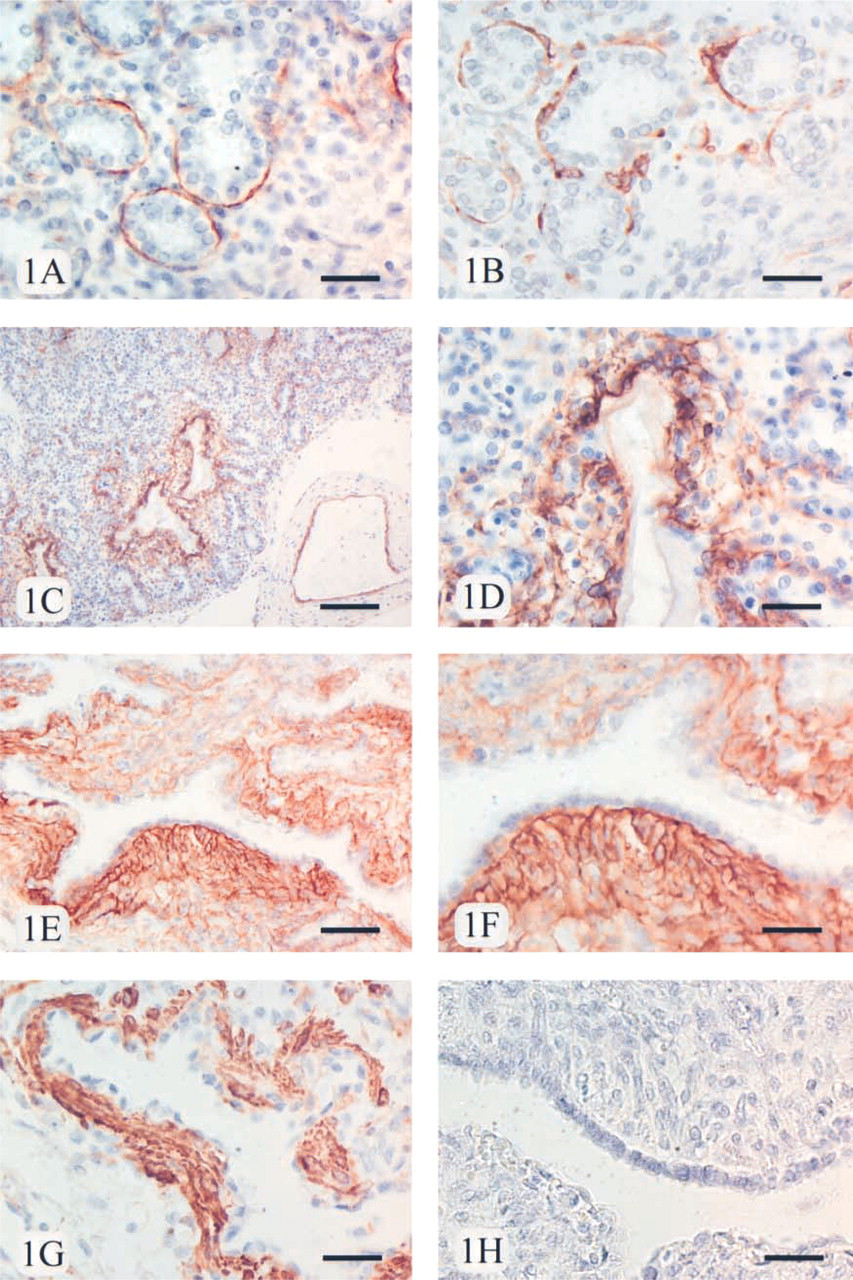
Tenascin-C immunoreactivity, scored as low (+) in all seven cases, was observed as thin and linear fibers underneath the basement membranes of the cuboidal alveolar epithelium consisting evidently of Type II prepneumocytes (Figure 1A. Low immunoreactivity (+) for tenascin-C was also detected underneath the bronchiolar epithelium and among smooth muscle cells in the walls of bronchi in 7/7 cases. Low (+) immunoreactivity was shown in the interstitium in all cases. In veins, the immunoreactivity for tenascin-C was moderate (++) in the intima, i.e., beneath the endothelial cells, in 7/7 cases. In contrast, no immunoreactivity for tenascin-C (0) was observed in the endothelial cells and intima of arteries. Media of veins was negative in all cases. Occasionally, in some small arteries a faint immunoreactivity for tenascin-C was shown in media. Strong immunoreactivity for tenascin-C (+++) was detected in and around chondrocytes in the walls of the bronchi in every control case. No immunoreactivity (0) was observed in mesothelial cells or submesothelial connective tissue of the pleura (Table 3).
RDS
In alveolar walls, the immunoreactivity for tenascin-C was usually strong (+++ in 14/19 cases) underneath the detached alveolar epithelium underlying the hyaline membranes, which were mainly negative for tenascin-C. The expression within alveolar walls was scored as moderate in five cases that had survived less than 1 day (++; case numbers 1, 3, 6, 9, and 10 in Table 2). The tenascin-C-immunopositive fibers were both linear and reticular (Figures 1C and 1D). Low (+, 12/19 cases) or moderate (++, 7/19 cases) tenascin-C immunoreactivity was seen in the cells of interstitium. Also underneath the bronchiolar epithelium, the expression of tenascin-C was moderate (++, 5/19 cases) or strong (+++, 14/19 cases), while the expression among the smooth muscle cells of the bronchi was moderate (++) in all cases. Strong immunoreactivity for tenascin-C (+++) was detected in and around chondrocytes in 19/19 cases. In the intima and the endothelial cells of the veins, the expression of tenascin-C varied from low to moderate (+, 3/19 cases; ++, 16/19 cases, respectively), whereas the endothelial cells of the arteries were all negative (0). No immunoreactivity (0) was observed in the mesothelial cells or submesothelial connective tissue of the pleura (see Table 3).
BPD
A very strong and wide IHC expression for tenascin-C (++++) was observed in the fibrotic interstitium of the remodeled alveolar walls underneath the regenerative epithelium in every case (Figures 1E and 1F), including the patients who had survived only 5 days (see Table 2). Also under the bronchiolar epithelium, the immunoreactivity for tenascin-C varied from strong (+++, 2/12 cases) to very strong (++++, 10/12 cases), while the expression among the smooth muscle cells of the bronchi was moderate (++, in 6/12 cases) or strong (+++, in 6/12 cases). As in controls and the RDS group, tenascin-C expression in and around chondrocytes was strong (+++) in every case. In the endothelial cells and intima of the veins, the tenascin-C immunoreactivity was low (+, 11/12 cases) or moderate (++, 1/12 cases), whereas in the endothelial cells of the arteries it was absent (0). No immunoreactivity for tenascin-C was observed in the pleura or mesothelium. For a summary of the results see Table 3.
mRNA ISH Findings
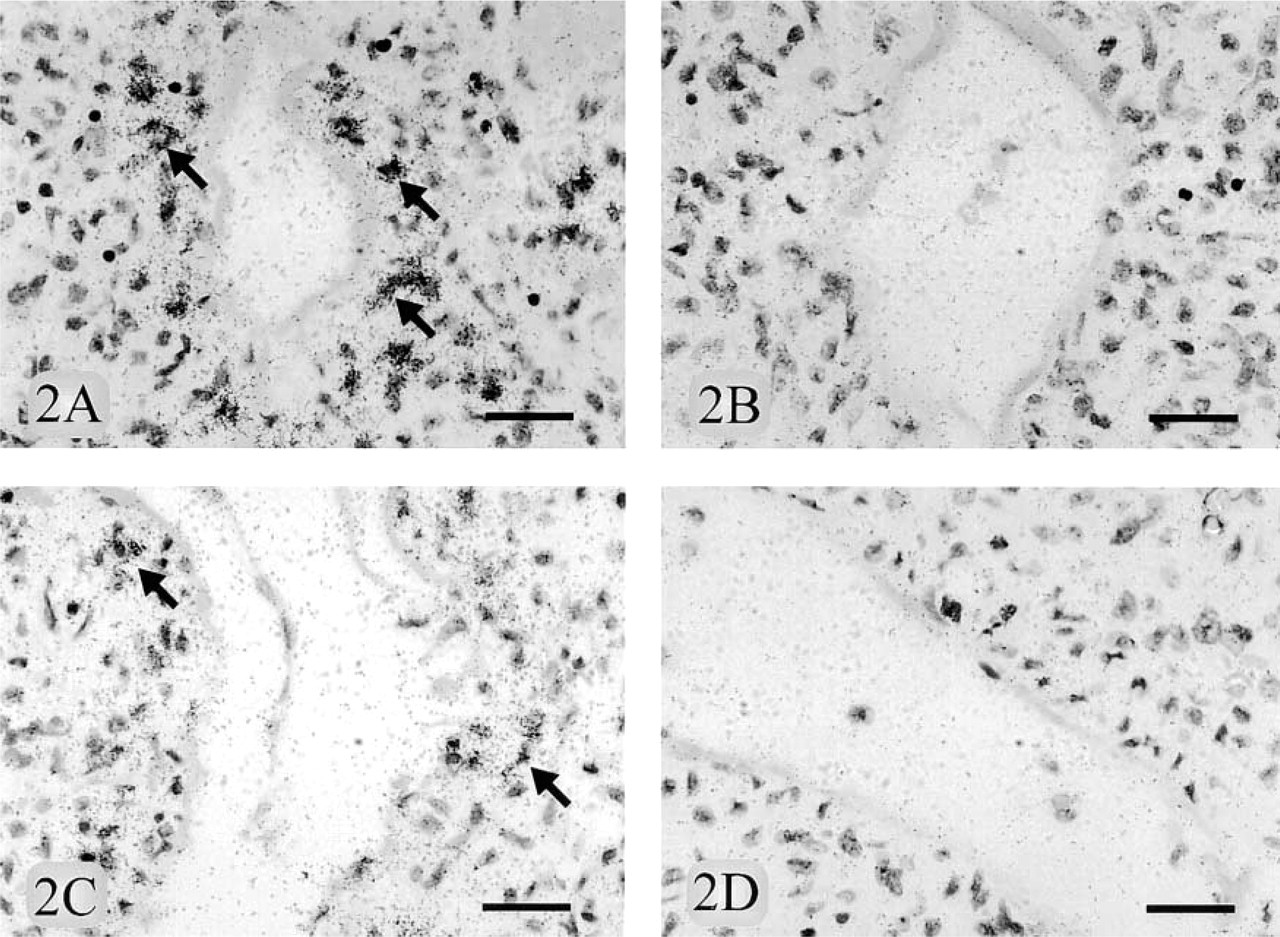
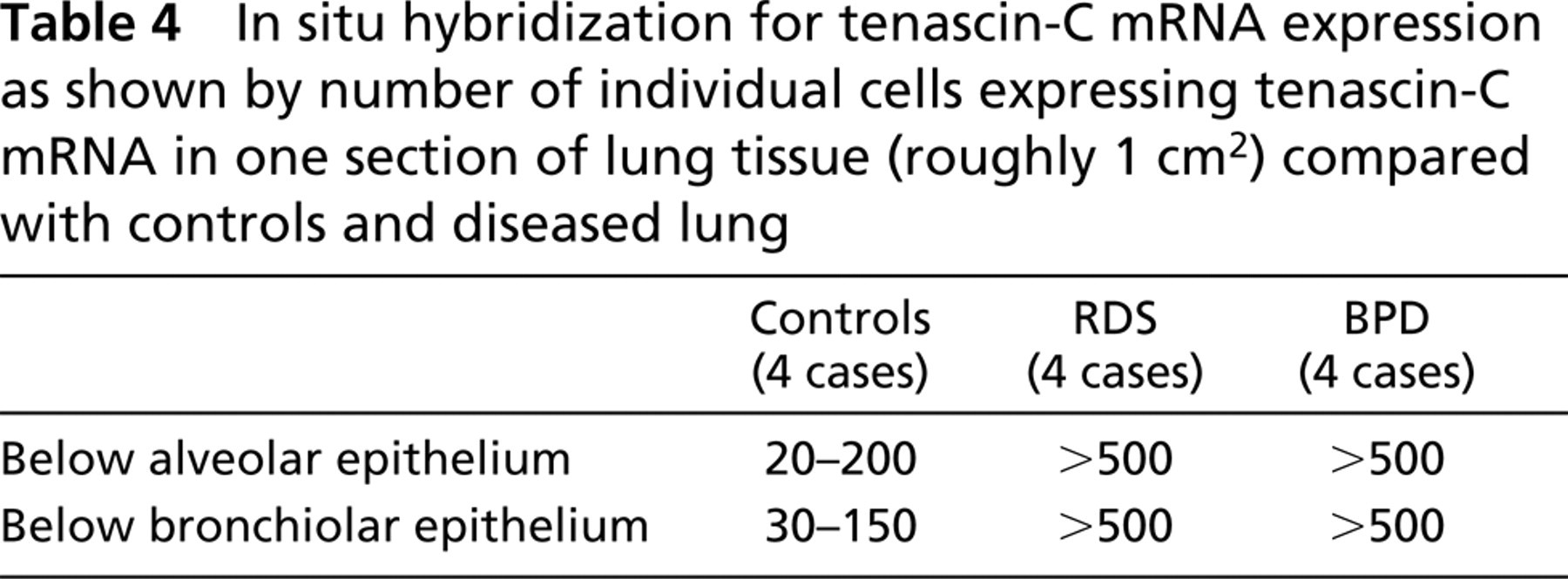
The number of mRNA grains was diffusely increased in all 12 cases in tissue hybridized with 35S-labeled antisense tenascin-C RNA probe compared to those hybridized with 35S-labeled sense RNA probe. In controls, the number of cells expressing tenascin-C mRNA varied from 20 to 200 underneath the alveolar epithelium and from 30 to 150 underneath the bronchiolar epithelium in one section of lung tissue (roughly 1 cm2). The corresponding numbers of tenascin-C mRNA-expressing cells below the alveolar and the bronchiolar epithelium were over 500 in both RDS and BPD (Figures 2A-2D) (see Table 4). IHC staining demonstrated that the cells in these locations were positive for α-smooth muscle actin, suggesting a myofibroblast phenotype. The number of tenascin mRNA grains was increased in the endothelial cells of veins and chondrocytes, but the number of positive cells in these locations did not vary in controls and diseased lung. In general, the expression of tenascin-C mRNA by ISH studies corresponded well with the findings of the IHC studies.
(
Scores of tenascin-C immunoreactivity in and around different types of pulmonary cells in control cases and patients with respiratory distress syndrome (RDS) and bronchopulmonary dysplasia (BPD)a
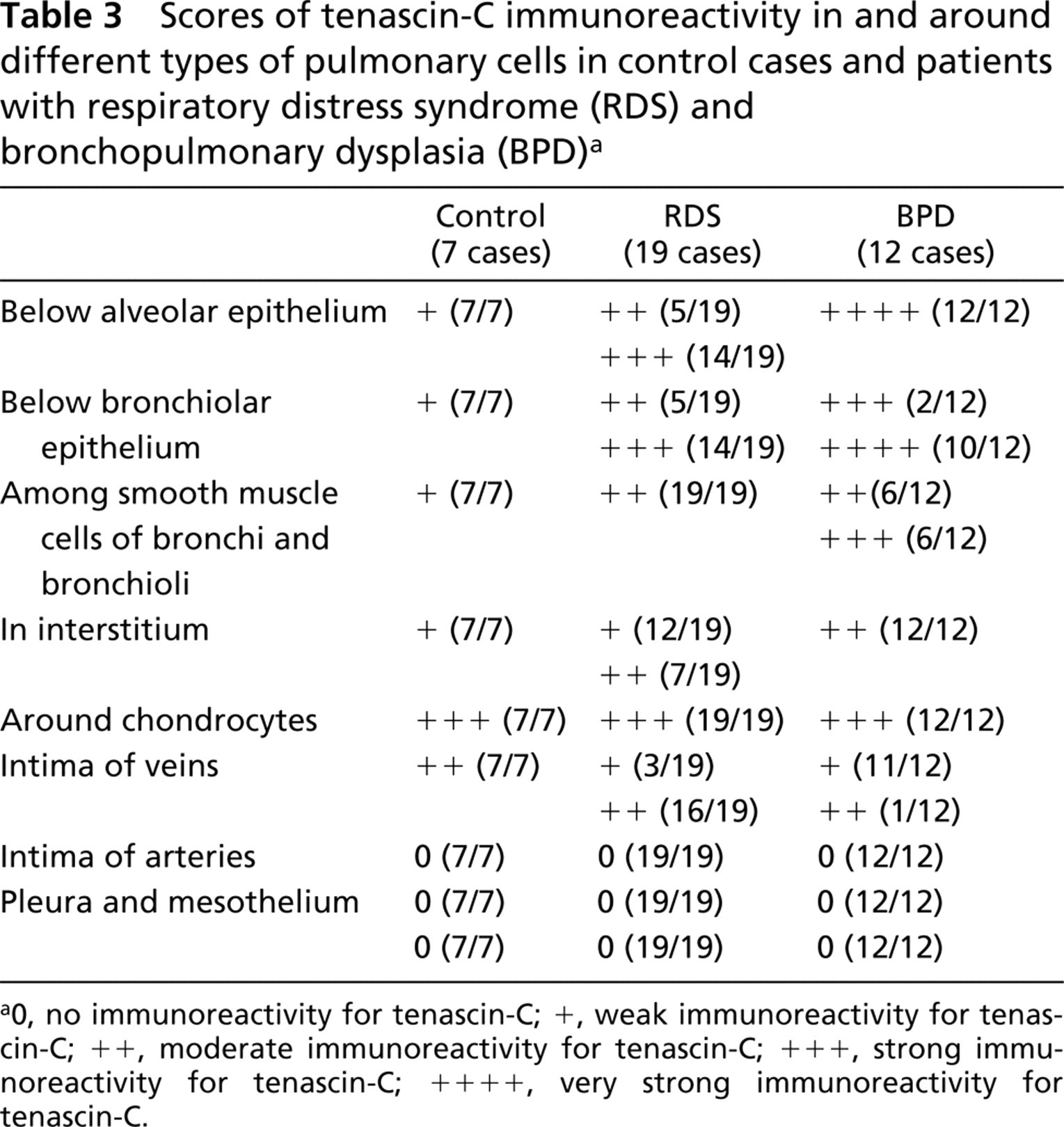
0, no immunoreactivity for tenascin-C; +, weak immunoreactivity for tenascin-C; ++, moderate immunoreactivity for tenascin-C; +++, strong immunoreactivity for tenascin-C; ++++, very strong immunoreactivity for tenascin-C.
IHC Findings for α-Smooth Muscle Actin
In controls, IHC staining for α-smooth muscle actin showed intracellular positivity in the single row of cells below both the alveolar (Figure 1B and the bronchiolar epithelium scored as low (+). The positive cells within the alveolar walls were localized beneath the basement membranes of the alveolar epithelium, and these cells corresponded to obviously myofibroblast-type cells. α-Smooth actin positivity is a typical phenotype for lung myofibroblasts (Kuhn and McDonald 1991; Toti et al. 1997). The positive immunoreactivity for α-smooth muscle actin was observed also in the media of arteries, scored as very strong (++++), and in the cells underneath the endothelia of veins (++). Very strong expression for α-smooth muscle actin in the media of arteries is caused by the abundant smooth muscle cells. A few interstitial cells were faintly positive for α-smooth muscle actin, whereas the submesothelial connective tissue cells of the pleura were negative.
The IHC reactivity for α-smooth muscle actin was stronger in the spindle-shaped cells underlying the alveolar epithelium in patients with RDS than in controls, and also in cells below the bronchiolar epithelium (scored as moderate ++ or strong +++) and in the interstitium. In patients with BPD, the cells expressing α-smooth muscle actin were highly increased in number underneath both the alveolar (Figure 1G and the bronchiolar epithelium (scored as strong +++ or very strong ++++) (Table 5). Bundles of α-smooth muscle actin-positive cells, which were obviously myofibroblasts, accumulated especially in the areas of newly formed interstitial fibrosis (Figure 1G. Some positively stained cells were also detected in pleural submesothelial connective tissue. The findings in the arteries and veins were similar in control and disease groups, i.e., patients with RDS and BPD.
Statistical Analysis
In comparing the control group (n = 7) to the group of the patients with RDS and BPD (n=31), the latter group showed significantly more strong or very strong tenascin-C immunoreactivity below alveolar epithelium (p<0.00001 by Fishers' exact probability test; Table 6). In comparing the cases with RDS and BPD that had survived less than 1 day (n=11) to the cases that survived 1 day or longer (n=20), the latter group showed significantly more often strong or very strong IHC expression for tenascin-C underneath alveolar epithelium than the previous group (p=0.002 by Fishers' exact probability test; Table 7).
Discussion
The role of various ECM glycoproteins, such as fibronectin and collagen, has been widely studied in both RDS and BPD (Gerdes et al. 1988; Cherukupalli et al. 1996; Sinkin et al. 1998). Fibronectin and another ECM glycoprotein, tenascin-C, share many structural and functional similarities, such as adhesive and reparative functions. Tenascin-C has also been shown to have anti-adhesive properties (Chiquet-Ehrismann 1991). In this investigation the IHC distribution and mRNA expression of tenascin-C were studied for the first time in patients with RDS and BPD. Moreover, we assessed semiquantitatively the IHC expression of tenascin-C in different types of pulmonary cells in these disorders and compared the findings with a control group of the same gestational age without a respiratory disease.
(
In control cases with a gestational age of 23–28 weeks, tenascin-C was localized in alveolar walls, whereas in adult normal alveolar interstitium its expression is absent (Kaarteenaho-Wiik et al. 1996). In the recent study of Toti and co-workers (1997), myofibroblasts appeared to be normally present along terminal airways in the developing lung. In our recent study we observed that myofibroblasts mainly produce tenascin-C in usual interstitial pneumonia (UIP), which is the most common type of adult idiopathic interstitial pneumonia, with a poor prognosis (Pääkkö et al. 2000). It is possible that myofibroblasts are responsible for the production of tenascin-C in developing human lung, because BPD has similar morphological features in common with adult pulmonary fibrotic disorders (Stocker 1986). Furthermore, in this study we observed that the number of α-smooth muscle-positive cells, which were apparently myofibroblasts, greatly increases in the newly formed fibrosis of BPD, corresponding to the same localization as the increased expression of tenascin-C protein and mRNA. Moreover, myofibroblasts have been shown to increase in number and to form bundles of cells encircling terminal air spaces some days after lung injury in neonates (Toti et al. 1997). These findings are in concordance with the results of the present study and suggest that myofibroblasts are mainly tenascin-C-synthesizing cells in RDS and BPD. In early chicken lung bud, however, tenascin-C mRNA was found to be synthesized by the epithelium, inconsistent with the results of the present study (Koch et al. 1991).
In situ hybridization for tenascin-C mRNA expression as shown by number of individual cells expressing tenascin-C mRNA in one section of lung tissue (roughly 1 cm2) compared with controls and diseased lung
Scores of α-smooth muscle actin immunoreactivity in different types of pulmonary cells in control cases and patients with RDS and BPDa
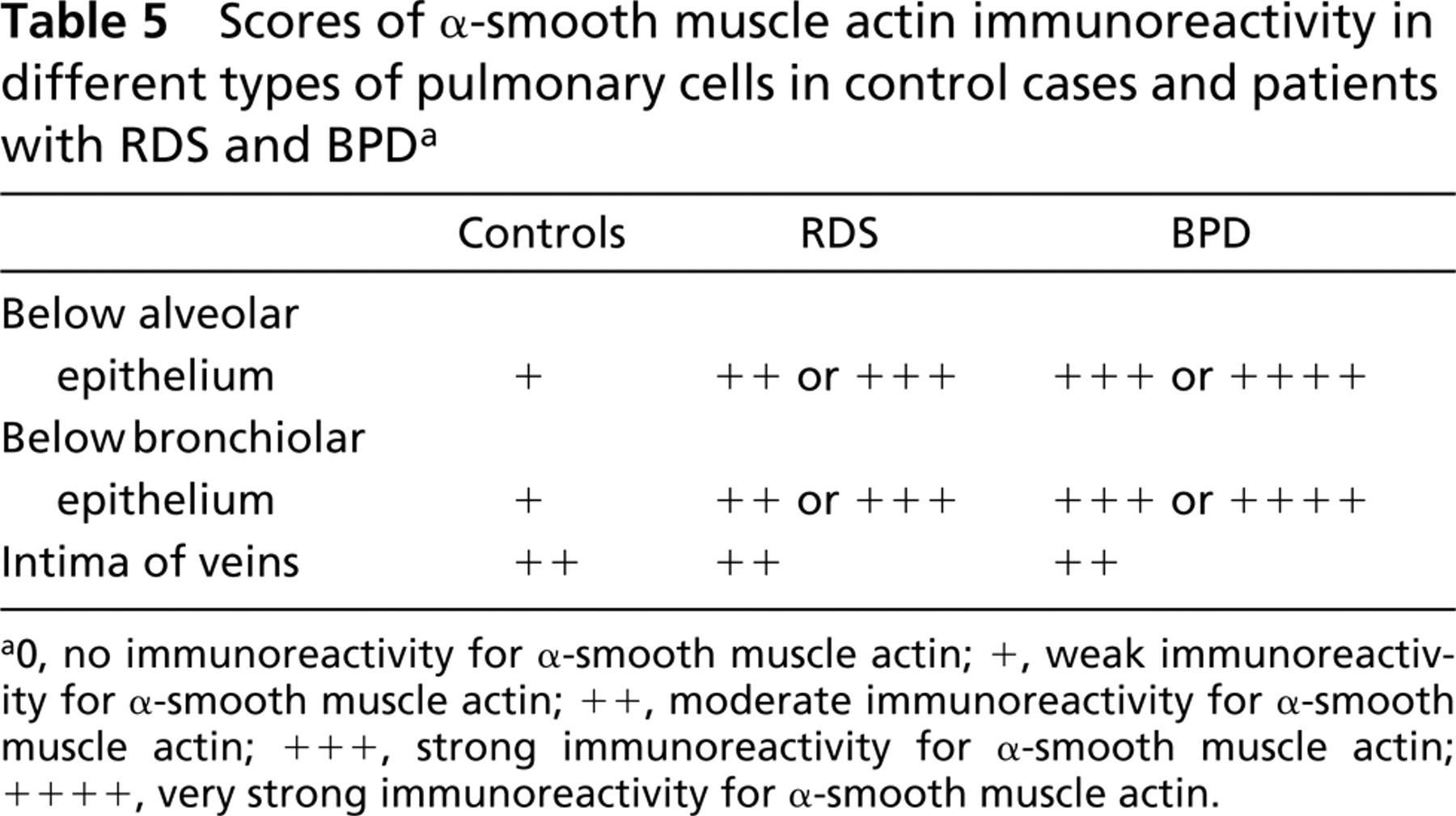
0, no immunoreactivity for α-smooth muscle actin; +, weak immunoreactivity for α-smooth muscle actin; ++, moderate immunoreactivity for α-smooth muscle actin; +++, strong immunoreactivity for α-smooth muscle actin; ++++, very strong immunoreactivity for α-smooth muscle actin.
Comparison of tenascin-C immunoreactivity below alveolar epithelium in controls with the patients with RDS and BPD
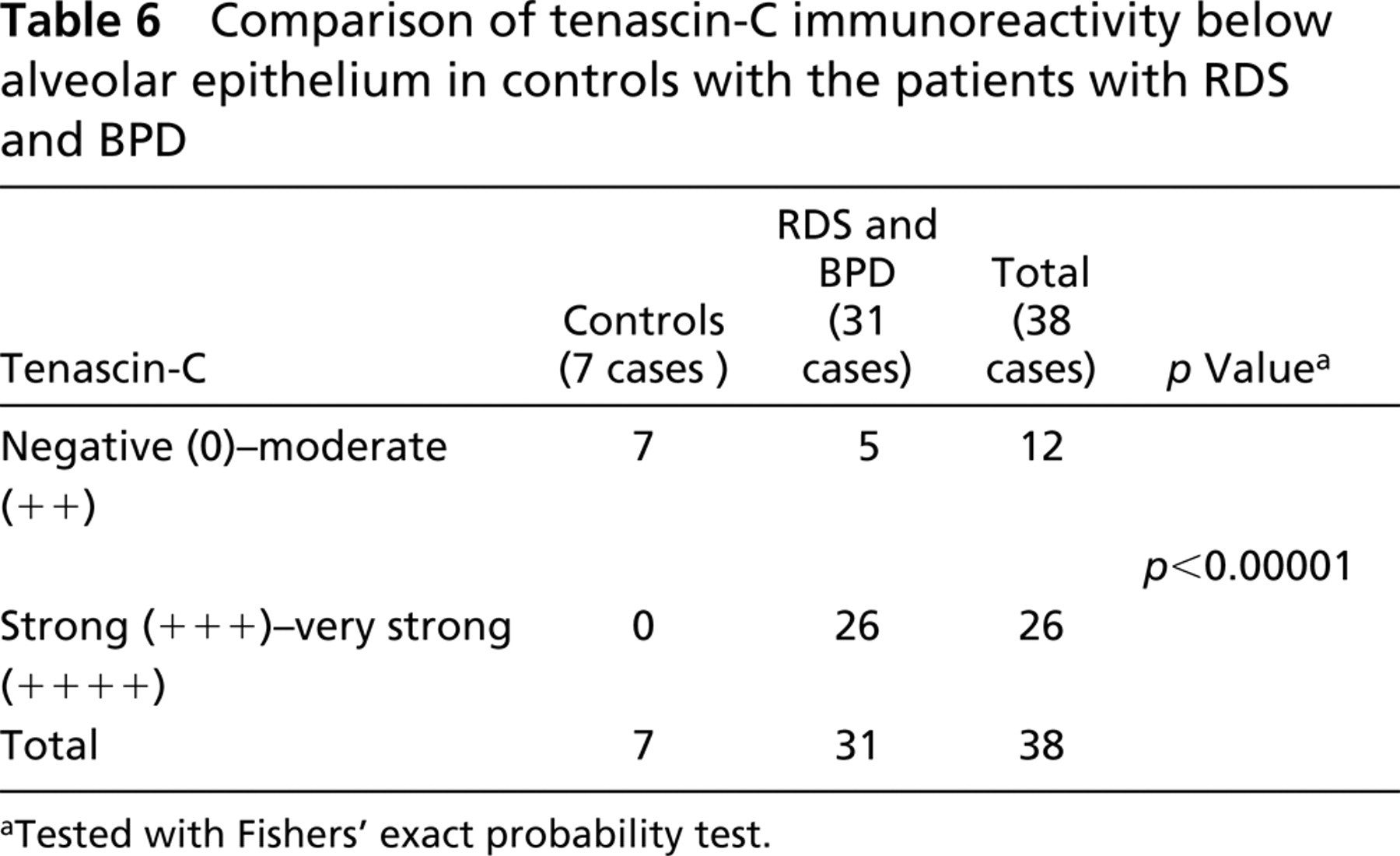
Tested with Fishers' exact probability test.
The IHC expression for tenascin-C within alveolar and bronchiolar walls was stronger in patients with RDS and BPD than in controls. In comparing the cases with RDS and BPD, the scores of tenascin-C were higher in BPD. This is understandable because the fibrotic process is more advanced in BPD than in RDS. By IHC, however, the scores of RDS and BPD showed no difference, which may indicate that the increased expression of tenascin-C mRNA begins rapidly after the injury. In adult pulmonary fibrotic disorders, tenascin-C has been shown to increase, especially in active, newly formed fibrosis (Kaarteenaho-Wiik et al. 1996). We observed a very strong expression of tenascin-C in every studied case of BPD. This suggests that the process of fibrogenesis is active in different stages of the disease, even after 2–3 months. Strong expression of tenascin-C was observed in every patient with RDS who survived for at least 1 day, whereas the five cases with moderate expression survived for less than 1 day. However, some RDS cases that survived only a few hours showed strong IHC immunopositivity for tenascin-C. This suggests that the synthesis of tenascin-C begins rapidly after injury in RDS. Our finding is supported by a recent study showing a maximal IHC tenascin-C expression at 24 hr in tracheal wound healing (Kanno and Fukuda 1994). In an IHC study by Sinkin and others hyaline membranes were shown to express fibronectin (Sinkin et al. 1998). We did not note marked immunoreactivity for tenascin-C in hyaline membranes in this study. We observed, however, that tenascin-C is mainly localized underneath the hyaline membranes overlying detached alveolar epithelium. This finding is in concordance with our previous studies on UIP in adults, in which tenascin-C was expressed especially at the sites of epithelial cell damage (Pääkkö et al. 2000). The upregulation of tenascin-C in the processes of epithelial damage and also in regeneration may be associated with cell growth, which would be in concordance with the result of the recent study showing that tenascin-C increases neurite outgrowth in culture (Meiners et al. 2001). The difference between the immunoreactivity for tenascin-C and fibronectin within the hyaline membranes may be due to the fact that fibronectin is produced in two forms, i.e., as a soluble form and a cellular form (Vartio et al. 1987), which is not known to occur for tenascin-C. Furthermore, tenascin-C, but not fibronectin, has been shown to induce apoptosis in mammary epithelial cells, at least in vitro (Boudreau et al. 1996).
Comparison of tenascin-C immunoreactivity in patients with RDS and BPD who had survived less than 1 day or longer

Tested with Fishers' exact probability test.
Tenascin-C has been shown to be elevated in epithelial lining fluid (ELF) and in serum of adult patients with pulmonary fibrotic disorders (Kaarteenaho-Wiik et al. 1998). Fibronectin is increased in bronchoalveolar lavage both in adults with fibrotic pulmonary disorders, and in infants with BPD (Rennard and Crystal 1981; Gerdes et al. 1988). Probably tenascin-C would be increased in ELF and, clinically more importantly, also in serum in patients with BPD. Interestingly, in this study very strong expression for tenascin-C was shown in lung tissue of BPD cases as early as 5 days of age. Perhaps serum tenascin-C could then function as a marker of developing fibrosis in children with RDS leading to attempts to decrease the fibrogenesis with proper medical treatment. BPD is often clinically diagnosed at the age of 28 days, although the fibrosis is known to begin as early as the age of 2 days. Interestingly, in a recent study by Zhao (1999), tenascin-C was observed to be expressed in the mesenchyme of the embryonic rat lung and downregulated by dexamethasone in early organogenesis.
In conclusion, we observed that tenascin-C is highly expressed within alveolar walls underneath detached alveolar epithelium underlying hyaline membranes in patients with RDS who survive for 1 day or more. It was very strongly expressed in remodeled fibrotic alveolar walls underneath regenerative alveolar epithelium in every patient with BPD, of whom some had survived only for 5 days. This suggests that there is an association between the expression of tenascin-C and the presence of these disorders and, moreover, that its expression increases rapidly after injury.
Footnotes
Acknowledgements
Supported by the Finnish Anti-Tuberculosis Association Foundation and the Paulo Foundation.
The technical assistance of Ms Mirja Vahera, Ms Erja Tomperi, Ms Annikki Huhtela, Ms Heli Auno, Ms Riitta Vuento, Mr Hannu Wäänänen, and Mr Tapio Leinonen is gratefully acknowledged.