
Abstract
We studied the expression and regulation of TIMP-3, a recently cloned member of the tissue inhibitor of the metalloproteinase family, during human fetal development and in various human tissues, with emphasis on epithelial structures. Expression of TIMP-3 mRNA was detected by in situ hybridization in developing bone, kidney, and various mesenchymal structures. At 16 weeks of gestation, ectoderm-derived cells of hair germs expressed TIMP-3 mRNA, and beginning from the twentieth week consistent expression was detected in epithelial outer root sheath cells of growing hair follicles. In normal adult human skin, expression of TIMP-3 mRNA was limited to hair follicles, starting at the early anagen (growing) phase and vanishing at the catagen (regressing) phase. TIMP-3 mRNA was not detected in benign hair follicle-derived tumors but was present in tumor cells of infiltrative basal cell carcinomas and in surrounding stromal cells in squamous cell carcinomas. Human primary keratinocytes in culture expressed TIMP-3 mRNAs, the levels of which were upregulated by transforming growth factor-β (TGF-β), whereas interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α) had no effect. Our results suggest a role for TIMP-3 in connective tissue remodeling during fetal development, hair growth cycle, and cancer progression.
T
TIMP-3 mRNA was first detected in human breast tumors (Uria et al. 1994), in metastatic melanoma cell lines (Silbiger et al. 1994), and in various normal adult tissues such as placenta, kidney, heart, prostate, small intestine, and lung, and in fetal tissues including heart, lung, and kidney by Northern blot hybridization (Apte et al. 1994b; Wick et al. 1994; Wilde et al. 1994). With in situ hybridization, expression of murine TIMP-3 mRNA has been reported during mouse embryo implantation (Harvey et al. 1995; Reponen et al. 1995; Alexander et al. 1996; Leco et al. 1996) and in developing kidney, cartilage, and various epithelial structures, including epidermis and intestinal mucosa (Apte et al. 1994a). Little is known, however, about the role of TIMP-3 in human tissues in vivo, because expression of TIMP-3 mRNA has only been localized to cells of early placental structures (Byrne et al. 1995; Higuchi et al. 1995; Hurskainen et al. 1996), fetal retinal epithelium (Ruiz et al. 1996), and fibroblastic cells within breast cancer stroma (Byrne et al. 1995). Furthermore, TIMP-3 protein has been found to be an extracellular matrix component of Bruch's membrane of the human eye (Fariss et al. 1997).
TIMP-3 is induced in response to mitogenic stimulation and is regulated during normal cell cycle progression (Wick et al. 1994). A potential role for TIMP-3 in carcinogenesis has been proposed. Chicken TIMP-3 promotes oncogenic transformation in cultured cells (Yang and Hawkes 1992), and TIMP-3 overexpression inhibits human colon carcinoma growth in vivo (Bian et al. 1996). A direct implication for a human disease is the presence of point mutations in the TIMP-3 gene in patients with Sorsby's fundic dystrophy, an autosomal dominant disorder leading to visual loss (Weber et al. 1994).
To explore the physiological role of TIMP-3 in human tissues, we have determined its spatial and temporal expression during fetal development. We have also examined its expression in adult skin and various organs of the body, with special emphasis on epithelial components based on previous data on mouse tissues (Apte et al. 1994a). We report here that in early human fetus TIMP-3 is expressed in cartilage, bone, kidney, and in mesenchymal cells within the connective tissue. At 16 weeks of gestation, ectoderm-derived cells of hair germs express TIMP-3 mRNA, and beginning from the twentieth week consistent expression is detected in epithelial outer root sheath cells of growing hair follicles. In fully developed follicles, expression of TIMP-3 mRNA is cyclic, starting at the early anagen (growing) phase and vanishing at the catagen (regressing) phase. TIMP-3 mRNA is not detected in benign hair follicle-derived tumors but is present in tumor cells of infiltrative basal cell carcinomas and in surrounding stromal cells in squamous cell carcinomas. In vitro TIMP-3 gene expression in primary human epidermal keratinocytes is induced by TGF-β, but not by IL-1β or TNF-α. Our results substantiate the role of TIMP-3 in development, hair growth cycle, and tumor growth.
Materials and Methods
Tissues
Formalin-fixed, paraffin-embedded specimens were obtained from the Departments of Dermatology and Pathology, University of Helsinki, Finland. All fetal material originated from medical abortions and was obtained from the Department of Pathology, University of Oulu, Finland. Fetal age was estimated by menstrual age and histological examination. This study was approved by the ethics committee of the Department of Dermatology, Helsinki, Finland. The following subgroups of histological sections were examined.
Fetal tissues: complete fetus at gestational age of 7, 8–9, 10, and 12 weeks and biopsies of the scalp and trunk skin at gestational age of 16, 20, 21, and 23 weeks.
Adult skin specimens: basal cell carcinoma n = 8 (infiltrative n = 5, keratotic n = 3), squamous cell carcinoma n = 5, trichofolliculoma n = 4, trichoepithelioma n = 4, blistering skin diseases n = 11 (dermatitis herpetiformis n = 5, pemphigus n = 2, pemphigoid n = 2, epidermolysis bullosa n = 2), and normal skin from various parts of the body n = 9.
Various organs with epithelial components displaying normal histology: kidney n = 3, liver n = 3, pancreas n = 3, parotic gland n = 3, prostatic gland n = 3, mammary gland n = 3, testis n = 3, bronchus n = 3, normal gastric mucosa n = 3, normal duodenal mucosa n = 3, and normal colon mucosa n = 3.
Carcinoma ductale mammae n = 4 and carcinoma adenomatosum coli n = 6.
Probes
A 518-
In Situ Hybridization
In vitro transcribed anti-sense and sense RNA probes were labeled with α[35S]-UTP. Sections were hybridized with probes (2.5–4 X 104 cpm/μl of hybridization buffer) and were washed under stringent conditions, including treatment with RNase A, as described (Saarialho–Kere et al. 1993). After autoradiography for 10–35 days, the photographic emulsion was developed and the slides were stained with hematoxylin and eosin. Samples of breast carcinomas were used as positive controls (Uria et al. 1994). Each sample was hybridized in at least two experiments, and a sense probe was used as a negative control. The slides were independently analyzed by two investigators.
Keratinocyte Cultures
Primary cultures of normal human epidermal keratinocytes were established from skin specimens from a woman undergoing mammoplasty for nonmalignant disease, as described previously (Boyce and Ham 1985). The cells were maintained in Keratinocyte Growth Medium (Clonetics; San Diego, CA) supplemented with epidermal growth factor (0.2 ng/ml) and bovine pituitary extract (30 μg/ml) (both from Life Technologies; Paisley, UK). Cells were incubated for 24 hr with human recombinant IL-1β (5 U/ml), TNF-α (20 ng/ ml) (both from Boehringer Mannheim; Mannheim, Germany) and bovine TGF-β2 (5 ng/ml) (kindly provided by Dr. David R. Olsen, Celtrix Co., Santa Clara, CA).
HaCaT cells, transformed human epidermal keratinocytes (obtained from Dr. Norbert Fusenig, DKFZ, Heidelberg, Germany) (Boukamp et al. 1988), were cultivated in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal calf serum. At confluence, cells were washed twice with PBS and incubated under serum-free conditions for 18–20 hr, followed by incubation with 5–50 ng/ml EGF (epidermal growth factor), 1–100 nM PMA (phorbol myristate acetate) (both from Sigma Chemical; St Louis, MO), or 0.1–10 ng/ml TGF-β1 (R & D Systems; Minneapolis, MN) for 6 hr.
RNA Analysis
Total cellular RNA was isolated from primary keratinocyte cultures using the guanidine thiocyanate–cesium chloride method (Chirgwin et al. 1979), and from HaCaT cells using the guanidine thiocyanate–phenol–chloroform extraction (Parks et al. 1988). Fifteen μg of RNA was fractionated on formaldehyde-agarose gel and transferred to nylon membranes. Northern blot hybridizations were performed as described previously (Thomas 1980) with cDNAs labeled with [α-32P]-dCTP using random priming and [32P]-cDNA-mRNA hybrids were visualized by autoradiography. The mRNA levels were quantitated by densitometric scanning of the X-ray films on gray scale with background subtraction using MCID software (Imaging Research; St Catharines, Ontario, Canada), and corrected for the levels of rRNA visualized by ethidium bromide staining.
Results
TIMP-3 mRNA Is Expressed in Fetal Kidney, Cartilage, Bone, and Mesenchyme
The youngest fetal sample studied was an embryo 7 weeks of age whose main organ systems had developed. Epidermis, seen as a single layer of cells, showed no signal for TIMP-3 mRNA, but expression was detected in neuroepithelium (data not shown), in developing kidney, and in the adjacent gonadal ridge (Figures 1A, 1B, and 1D), and in cells within mesenchymal tissues (data not shown). Developing bone, heart, liver, and lungs, as well as bronchial epithelium, remained negative. At 8–9 weeks of gestation, chondrocytes of hand and foot plate cartilages (Figures 2A and 2B) and vertebral bodies (data not shown) expressed TIMP-3 mRNA. In other organs the distribution of TIMP-3 mRNA remained the same. At 10 weeks, signal was again detected in chondrocytes of ribs and limbs, and surrounding mesenchymal cells were positive (data not shown). At 12 weeks of gestation, expression of TIMP-3 mRNA was seen in hypertrophic chondrocytes of developing ribs and in surrounding mesenchymal cells that become osteoblasts (Figures 2C and 2D). In the same sample, bones that were already undergoing ossification had TIMP-3-expressing osteoblasts within the newly formed bone matrix (data not shown). Unlike in mouse embryos (Apte et al. 1994a), TIMP-3 mRNA was not detected in developing epidermis or in the epithelium of gastrointestinal tract or bronchial trees (data not shown).
At 7 and 8–9 weeks of gestation, no expression of TIMP-1 mRNA was detected. However, at 12 weeks mesenchymal cells surrounding hypertrophic cartilage (Figures 2E and 2F) were positive. In addition, osteoblasts within the bone matrix showed intense signal, as described earlier (Nomura et al. 1989). Chondrocytes remained negative.
Outer Root Sheath Cells of Hair Follicles Express TIMP-3 mRNA at the Anagen Phase
Biopsies of armpit and scalp skin at 16 weeks of gestation showed hair germs and early hair buds penetrating the underlying dermis. Epidermis was devoid of signal, but epithelial cells of the hair germs showed expression of TIMP-3 mRNA (Figure 3A, Inset a). By 20 weeks, follicles of the scalp showed established morphology, with epithelial cells forming the outer root sheath and mesenchymal cells forming the dermal papilla. Cells of the outer root sheath were consistently positive for TIMP-3 mRNA (Figure 3B, Inset b). In trunk skin, signal was again detected in hair germs and buds, and some follicles already in a more advanced stage of development showed signal in the outer root sheath cells (data not shown). In skin biopsies at 21 and 23 weeks of gestation, outer root sheath cells of each follicle were positive for TIMP-3 mRNA in the biopsies of both scalp and trunk (data not shown).
In adult hair follicles, TIMP-3 gene was activated during the early anagen (growing) phase and the expression persisted until the follicle entered the catagen phase and started to regress. TIMP-3 mRNA localized mainly to basal cell layer of the outer root sheath in the lower portion of the follicle (Figure 3C, Inset c). In some anagen follicles, a few pale-staining medulla cells within the hair matrix were also positive (Figure 3C). No expression was seen in telogen (resting) follicles (data not shown).
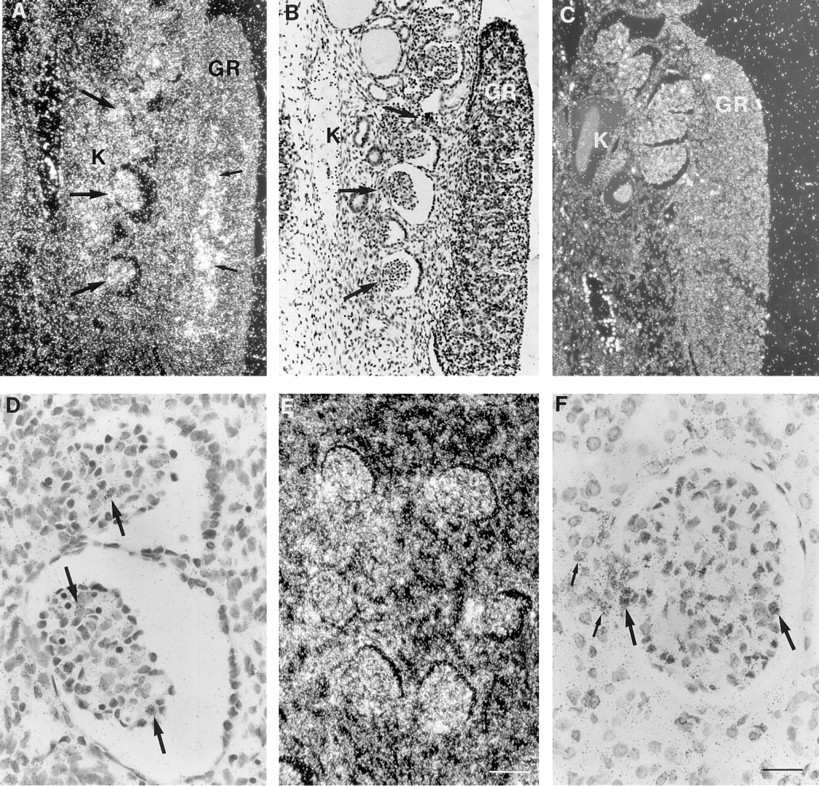
Expression of TIMP-3 mRNA in fetal and adult kidney. Samples were hybridized with TIMP-3 cRNA antisense (
TIMP-3 was not upregulated during re-epithelialization of blistering skin diseases, and expression was not detected in normal epidermis or in sebaceous and sweat glands. However, expression was often seen in fibroblast-like stromal cells surrounding sweat glands and some blood vessels (data not shown). TIMP-1 mRNA was commonly detected in sebaceous glands and in perivascular cells (data not shown) but only occasionally in some outer root sheath cells of anagen hair follicles (Figure 3D). Expression was not detected in fetal hair follicles.
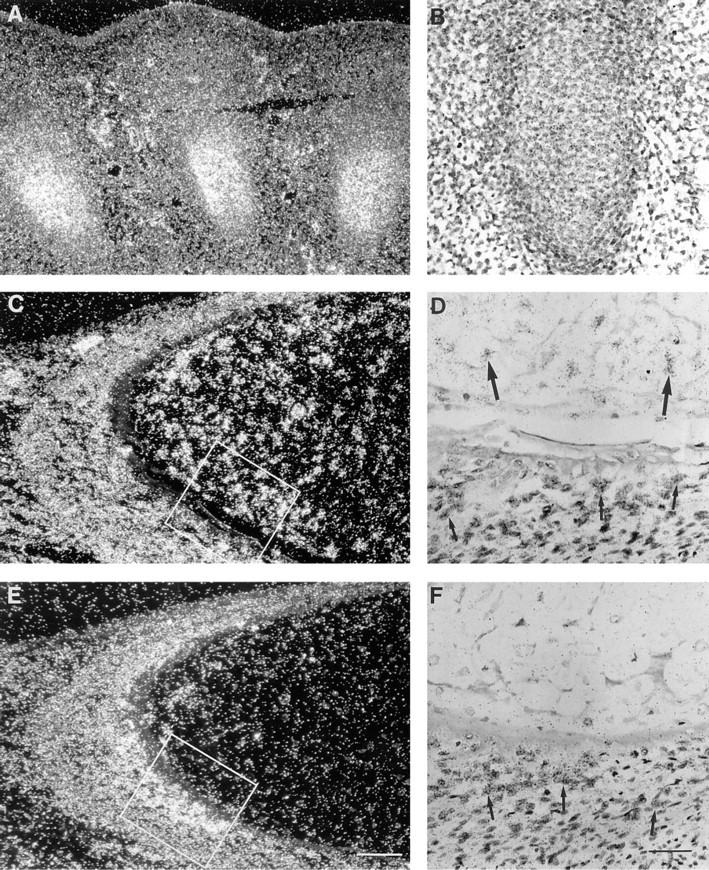
Fetal hypertrophic chondrocytes express TIMP-3, but not TIMP-1 mRNA. Samples were hybridized with TIMP-3 (
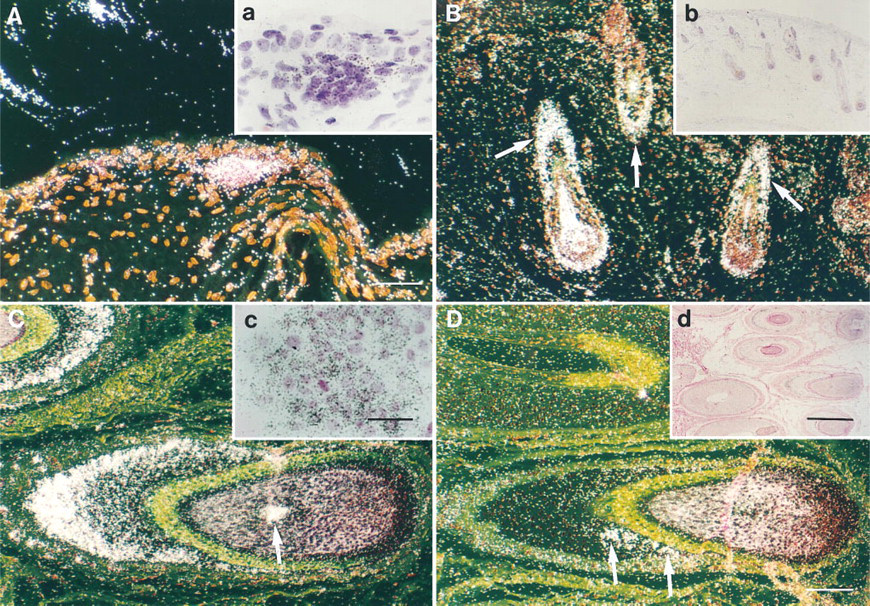
TIMP-3 is expressed during the hair growth cycle. In situ hybridizations with TIMP-3 (
TIMP-3 Expression Is Induced by TGF-β in Primary Human Keratinocytes
Because TIMP-3 was expressed in hair follicles, we studied its expression in keratinocytes treated with various growth factors known to influence hair growth cycle. Primary human epidermal keratinocytes were treated with TGF-β2, IL-1β, and TNF-α, and levels of TIMP-3 mRNAs were assayed by Northern blot hybridizations. As shown in Figure 4, epidermal keratinocytes expressed clearly detectable levels of three distinct TIMP-3 mRNAs (2.4, 2.8, and 4.8
TIMP-3 Is Expressed in Cancer Tissues
To determine whether TIMP-3 also plays a role in the behavior of benign and malignant skin tumors, samples of hair follicle-derived tumors, trichofolliculomas and trichoepitheliomas, and epidermal skin cancers were examined. Interestingly, no signal for TIMP-3 mRNA was detected in the benign tumors (data not shown). Basal cell carcinomas with keratotic (hairlike) differentiation were also negative, whereas in four of five infiltrative basal cell carcinomas TIMP-3 mRNA was detected in tumor cells at the margins of this aggressively growing tumor. In these samples, TIMP-3 mRNA was also detected in areas showing nodular tumor growth (Figure 5A, Inset a) but not in stromal cells. In squamous cell carcinomas, all of which were well- or rather well-differentiated, some stromal cells located diffusively adjacent to the tumor expressed TIMP-3 mRNA (Figures 5C and 5D, Inset c), while malignant cells were negative. There was no correlation between the number of cells expressing TIMP-3 mRNA and the histopathology of the tumor. As in the samples representing normal skin, TIMP-3 mRNA was present in fibroblast-like cells surrounding sweat glands deeper in the dermis.
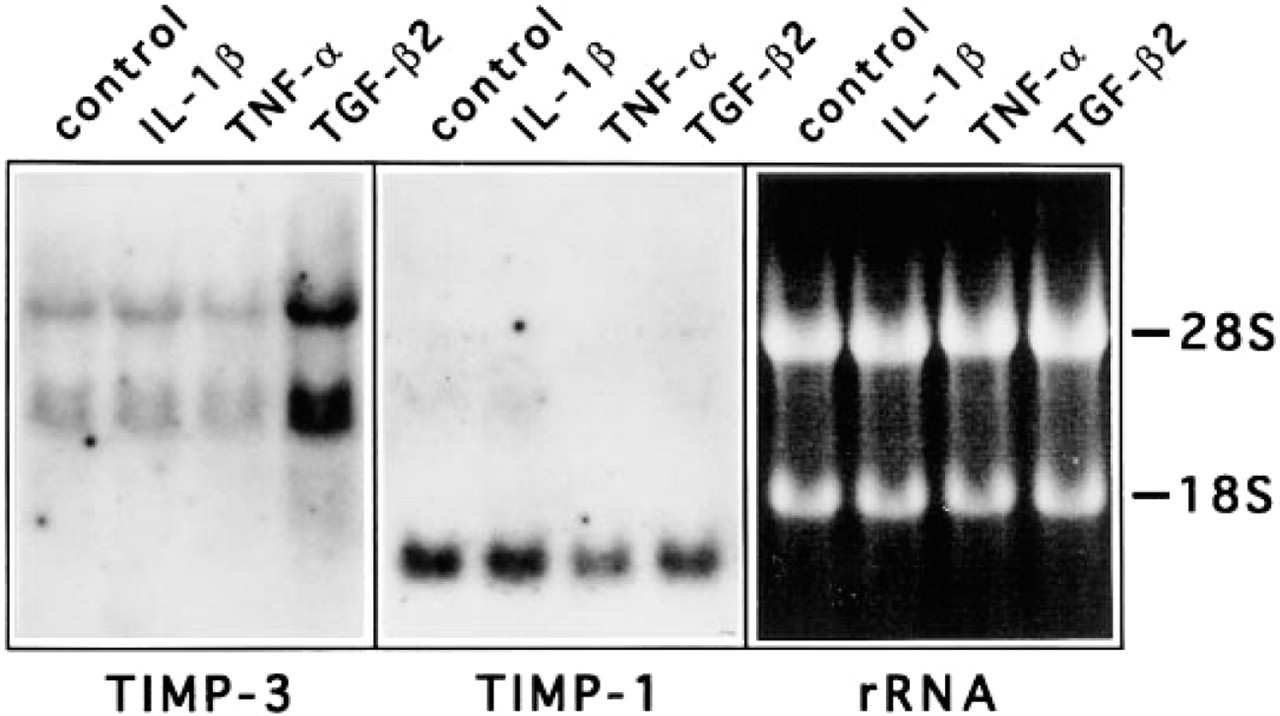
Expression of TIMP-3 mRNAs in epidermal keratinocytes is enhanced by TGF-β. Human primary epidermal keratinocytes were maintained as described in Materials and Methods and treated for 24 h with IL-1β (5 U/ml), TNF-α (20 ng/ml), and TGF-β2 (5 ng/ml). TIMP-1 and -3 mRNA levels were examined by Northern blot hybridizations of 15 μg of total RNA, and 18S and 28S rRNAs were visualized by ethidium bromide staining. IL-1β and TNF-α had no marked effect on TIMP-3 mRNA expression, whereas TGF-β2 induced the expression by 4.3-fold. None of these cytokines markedly altered TIMP-1 mRNA levels.
Normal gastrointestinal mucosa and mammary gland showed no signal for TIMP-3 mRNA. However, in samples of colon carcinoma intense expression was detected in both macrophages and spindle-like fibroblasts adjacent to the tumor tissue (data not shown). In agreement with the results of Byrne et al. (1995), myoepithelial cells as well as some fibroblasts surrounding tumor nodules of intraductal mammary carcinoma consistently expressed TIMP-3, whereas tumor cells remained negative (Figures 5E and 5F). In both colon and mammary carcinomas expression of TIMP-1 co-localized with that of TIMP-3 (data not shown).
Constitutive expression of TIMP-3 mRNA was also studied in some other organs with epithelial structures. In normal adult kidney, tubule epithelial and glomerular cells expressed TIMP-3 mRNA (Figures 1E and 1F). However, the glandular epithelia of liver, pancreas, parotid gland, testis, and prostate were negative. In addition, bronchial epithelium and adult bronchial cartilage were devoid of signal. The abundance of mRNA in Northern blot hybridizations of these organs (Apte et al. 1994b; Wick et al. 1994; Wilde et al. 1994) can be explained by the presence of various amounts of TIMP-3-expressing stromal cells, especially if any inflammation is present.
Discussion
The controlled physiological degradation of the ECM requires a fine balance between MMPs and their inhibitors, whereas in processes such as arthritis and tumor invasion MMP activity predominates, leading to excessive tissue degradation. There are accumulating data on the role of TIMPs and synthetic MMP inhibitors in suppressing tumor growth and invasion. Additional functions independent of metalloproteinase inhibitory activity have also been reported: TIMP-1 and -2 are anti-angiogenic (Moses et al. 1990; Johnson et al. 1994) and they possess growth-promoting activity towards a number of cell types (Hayakawa et al. 1992,1994). The role of TIMP-3 in development and connective tissue turnover has thus far been studied mostly in cell cultures, in animal models, and by analyzing mRNA from tissues by Northern hybridization. In the present study we have extended the analysis of TIMP-3 in vivo by examining its expression in human tissues. We also show that the levels of TIMP-3 transcripts are induced in human primary keratinocytes by TGF-β.
Various metalloproteinases and their inhibitors are involved in mammalian development. During the murine peri-implantation period, interstitial collagenase and stromelysin-1 mRNAs are produced by the embryo (Brenner et al. 1989). In later stages of murine embryogenesis, collagenase expression is restricted to hypertrophied chondrocytes, osteoblasts, endothelial cells, and osteoclasts of developing bones (Mattot et al. 1995). Rodent collagenase is suggested to be analogous to human collagenase-3, and these findings are consistent with our recent report on the expression of human collagenase-3 in hypertrophic chondrocytes and osteoblasts during human fetal bone development (Johansson et al. 1997). The 72-kD gelatinase is expressed widely in murine mesenchymal tissues, whereas 92-kD gelatinase mRNA production is limited to osteoclastic cells (Reponen et al. 1994). TIMP-1 and -2 are also predominantly expressed in osteogenic tissues, with some expression of TIMP-1 mRNA also in kidney, lung, ovary, and amnion (Nomura et al. 1989; Flenniken and Williams 1990; Mattot et al. 1995). TIMP-3 transcripts were found at sites of active matrix remodeling, such as developing bone and hair follicles. Co-localization of MMPs and their inhibitors in these areas suggests a coordinate process of ECM formation and degradation in the development and maintenance of normal tissue architecture. TIMP-3 does not appear to have a major role in the epithelial folding and branching during human fetal development, in contrast to the results of Apte et al. (Apte et al. 1994a) in mouse embryos. However, their samples represented murine embryos of 12.5 to 14.5 days of gestation as well as newborn mice, and we cannot exclude the possibility that in human development TIMP-3 is very transiently expressed during a time period that is not covered by our samples.
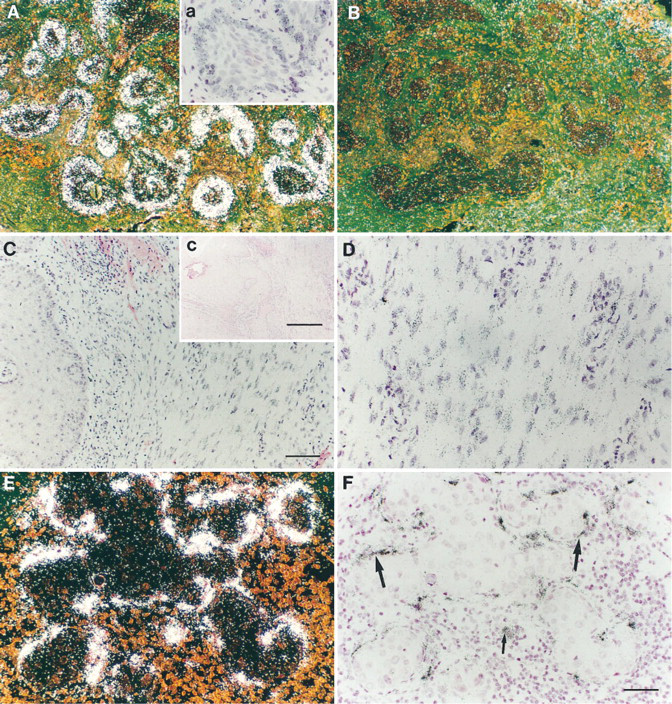
Expression of TIMP-3 mRNA in malignant tumor tissues. Samples were hybridized with TIMP-3 anti-sense and sense probes as described. (
Expression of TIMP-3 was detected throughout fetal hair development, starting from the hair germ stage. Hair germ formation begins in the scalp and face during the third month of gestation and gradually extends in a cephalocaudal direction. The germ consists of a group of epidermal basal cells that protrude into the dermis, forming a hair bud. Mesenchymal cells beneath each bud give rise to the dermal papilla. Further differentiation leads to the formation of the hair cuticle with the surrounding layers of inner and outer root sheath. From the beginning of the fifth month of gestation, different developmental stages are found, ranging from mature follicles to new developing ones (Lever and Schaumburg–Lever 1990). The factors that regulate the process of follicle development and growth cycle are not well understood, but they include epithelial–mesenchymal interactions and cytokines, such as epidermal growth factor (EGF), transforming growth factor-α (TGF-α), and members of the TGF-β family (Messenger 1993). The expression of TIMP-3 is stimulated in mouse cells by various agents, including PMA, EGF, TGF-β, dexamethasone, and TNF-α (Leco et al. 1994; Sun et al. 1995). However, in human primary keratinocytes only TGF-β induced expression of TIMP-3, while IL-1β and TNF-α had no effect. None of these cytokines markedly affected TIMP-1 expression. TGF-β2 mRNA co-localizes with TIMP-3 in the basal cells of the outer root sheath (Schmid et al. 1996), and since TGF-β inhibits growth of cultured hair follicles (Philpott et al. 1990) it may induce TIMP-3 to control follicle growth and degradation of the surrounding ECM.
Immunoreactivity for interstitial collagenase and matrilysin (MMP-7) has been detected in human hair follicles (Karelina et al. 1994; McGowan et al. 1994). Cultured hair follicles synthesize and secrete various MMPs, including interstitial collagenase, stromelysin-1, and gelatinases (Weinberg et al. 1990; Goodman and Ledbetter 1992; Paus et al. 1994). Cyclic expression of TIMP-1 in the inner root sheath of mouse hair follicles has been reported, with gene activation during midanagen phase (Kawabe et al. 1991). Our results demonstrate constitutive expression of TIMP-3 in both fetal and adult anagen follicles. TIMP-3 appears to contribute to the inhibition of proteolysis associated with degradation of the dermal matrix during the initial hair follicle formation, and later during the early anagen phase as the cells proliferate and invade the deeper dermis. On the basis of our studies, TIMP-1 does not have a consistent role in the growth cycle of human hair follicle. Our findings on the expression patterns of TIMP-1 and -3 in human tissues indicate that there are differences among species, and therefore the results obtained using animal models can not be directly applied to humans.
In cutaneous squamous cell carcinomas as well as mammary and intestinal carcinomas, TIMP-3 mRNA was detected in stromal cells adjacent to the malignant tumor, which is a common expression pattern for both MMPs and TIMPs. TIMP-3 was not expressed in the benign tumors and the nonaggressive keratotic basal cell carcinomas, but the signal was distinct in the infiltrative subtype and was localized to the malignant cells at the margins of tumor islands. In contrast, both TIMP-1 and -2 are detected only in the stromal cells of surrounding basal cell carcinoma (Childers et al. 1987; Wagner et al. 1996). Furthermore, expression of TIMP-2 mRNA is lower in cutaneous squamous cell carcinomas and infiltrative subtypes of basal cell carcinoma, compared to less infiltrative subtypes (Poulsom et al. 1993; Wagner et al. 1996). In cutaneous malignant melanomas, induction of both TIMP-1 and -3 mRNA expression in vivo correlates with increased depth of invasion (manuscript in preparation). Therefore, malignant transformation appears to induce the TIMP-3 gene, and in skin tumors invasive growth is associated with enhanced TIMP-3 expression, concomitantly with increased expression of matrix metalloproteinases (Ray and Stetler-Stevenson 1994). This suggests a role for TIMP-3 in the inhibition of metalloproteinase-mediated basement membrane and matrix degradation required for malignant growth.
In this study, TIMP-3 and -1 were differentially expressed during human fetal development. Furthermore, they were differently regulated in keratinocyte cultures by various growth factors. Further studies are needed to determine the level and specific functions of TIMP-3 protein in various human tissues. On the basis of the expression pattern of TIMP-3 mRNA, TIMP-3 appears to protect the matrix from proteolytic activity and thus regulate normal tissue turnover and inhibit malignant growth. Our results demonstrate a role for TIMP-3 in human fetal development and in remodeling processes of the extracellular matrix.
Footnotes
Acknowledgements
Supported by the Sigrid Jusélius Foundation, the Academy of Finland, the Finnish Cancer Foundation, the Paulo Foundation, the Finska Läkaresällskapet, and grants from Helsinki and Turku University Central Hospitals.
We thank Dr David Carmichael for the TIMP-1 cDNA, Dr Jorma Keski–Oja for the HaCaT cells and TGF-β1, Dr Juha Peltonen for primary keratinocytes, Dr Riitta Herva for pathology expertise, and Ms Alli Tallqvist for excellent technical assistance.