
Abstract
Flax fibers have been the subject of many biochemical studies, which revealed that cellulose and pectins are the major constituents of their walls. In contrast, little is known about the location of these polymers within the walls of mature fibers by microscopic methods. This has been technically hampered by the very thick secondary wall of fibers, resulting in inadequate tissue preservation unsuitable for immunogold microscopy. In this study, we adapted the basic chemical fixation, dehydration and infiltration methods to achieve a good preservation of the cell structures of mature fibers and reduced damage to antigens. We were able to apply postembedding immunocytochemical techniques to map the location of various pectic epitopes within the walls of mature fibers. Our immunolabeling data show that homogalacturonans were exclusively found in the middle lamellae and the cell junctions but were not detectable in the secondary wall. In contrast, rhamnogalacturonan I (RG I)-associated epitopes, as well as galactan and arabinan epitopes, were abundantly distributed over the secondary wall of mature fibers.
Because of our long-standing interest in flax fiber as a model for secondary cell-wall synthesis, we wanted to use immunogold microscopy to study the location and distribution of polysaccharides in the walls of immature and mature fibers using highly specific probes. Immunocytochemistry (ICC) is a powerful technique that allows the identification of specific macromolecules within a single cell. It also provides unique information on how the macromolecules are distributed within a specific cell domain (e.g., cell wall), which cannot be achieved by biochemical methods (Monaghan and Moss 1995). The successful application of ICC depends on achieving a balance between structural preservation and retention of the antigenicity of the molecules that are to be located by specific antibodies (Brorson 1998).
Recently, we have been able to perform ICC analyses on LR White-embedded immature flax fibers (1- to 2-week-old plants) using classical fixation and embedding techniques (His 1999; Andème-Onzighi et al. 2000). In contrast, this could not be achieved with more mature fibers (e.g., 8–10-week-old plants). Therefore, we have sought to modify the basic fixation, dehydration, and embedding steps to prepare mature flax tissues for immunogold microscopy.
Our first task was to achieve a good preservation of the ultrastructure, particularly for the fiber cell tissue, and the second was to map the distribution of different epitopes within the greatly thickened secondary walls of mature fiber cells. To the best of our knowledge this represents the first attempt to obtain information on the occurrence and location of polysaccharides in mature fiber walls (capsulation stage) using immunogold microscopy.
Materials and Methods
Plant Material
Flax plants (Linum usitatissimum L., variety Ariane) were harvested from a field near Rouen (Motteville, France) at different stages of their development.
Sample Preparation for Microscopy
Small fragments from the 18th internode were excised and immediately vacuum-fixed for 90 min in 2% (w/v) paraformaldehyde (Prolabo; Fontenay-Sous-Bois, France), 0.5% (v/v) glutaraldehyde (Oxford Agar), 0.1 M phosphate buffer (pH 7.2). The samples were rinsed in the same buffer, dehydrated in a graded aqueous ethanol series (10, 20, 35, 40, 50, 60, 70, 80, 90, 100%: 30 min each step) and further dehydrated with two washes in propylene oxide for 60 min each. They were then progressively infiltrated with LR White resin (LRW), according to the following schedule: 10, 30, 50, 70, 90% LRW/propylene oxide for at least 24 hr each step, and several incubations in 100% LRW. This last step was adapted to each sample depending on the age of the plant. It lasted about 90 days for mature samples and 30 days for younger ones. The first two infiltration steps were done under vacuum. The infiltrated samples were then embedded in gelatin capsules and polymerized for 24 hr at 60C.
Light Microscopy and Histochemical Staining
Staining of Fresh Samples with Safranin-Light Green. Hand-cut cross-sections from fresh stems were brought to 50% ethanol solution (v/v), then stained in safranin solution (Sigma; St Louis, MO) for 2 hr as described by Johansen (1940). The sections were thoroughly washed in water for 5 min or longer before being transferred to a solution of 95% ethanol and counterstained with Light Green staining solution for 15 sec. The sections were rinsed in alcohol before examination with a light microscope (Axioskop microscope; Zeiss, Oberkochen, Germany). Lignified and cutinized cell walls were stained brilliant green, cytoplasm and cellulose cell walls brilliant red.
Staining of LRW-embedded Samples with Methylene Blue-Basic Fuchsin. The staining was performed according to the protocol of Humphrey and Pittman (1974). Semi-thin sections (2.5 μm) were cut from LRW-embedded samples and mounted on glass microscope slides. Slides were covered with methylene blue (Sigma) for 20 min at 60C and rinsed thoroughly with distilled water to eliminate the non-fixed dye. The slides were then incubated with basic fuchsin (Sigma) for 30 sec to 5 min and rinsed thoroughly with water before observation. Lignified cell walls were stained green, cytoplasm and chloroplasts gray-blue, and non-lignified/cellulose cell walls pink.
Electron Microscopy and Immunogold Labeling
Ultrastructural Observations. Ultrathin sections (90 nm) from LRW-embedded samples were submitted to PATAg (periodic acid thiocarbohydrazide silver proteinate) test, which allowed polysaccharide detection as previously described (Roland and Vian 1991).
Detection of Anionic Sites with the Cationic Gold Probe. Labeling with cationic gold probe was performed on ultrathin sections (90 nm) from LRW-embedded samples collected on gold grids and treated with 3% acetic acid at pH 2.6 for 15 min at room temperature. The sections were incubated on a drop of 10-nm cationic poly-
Immunolabeling Procedure
The antibodies used in this study included: (a) the polyclonal antibodies anti-PGA/RG I, which recognize the unesterified polygalacturonic acids of the complex polygalacturonic acid (PGA)/rhamnogalacturonan I (RG I) (Moore et al. 1986; Moore and Staehelin 1988; Lynch and Staehelin 1992), (b) the monoclonal antibody (MAb) JIM5, characterized by Knox et al. (1990) and Willats et al. (2000), specific for ho-mogalacturonans, (c) the MAb LM5 specific for β-1,4-galactans (Jones et al. 1997), (d) the MAb LM6 specific for α-1,5-arabinan-epitopes (Willats et al. 1998), and (e) the MAb CCRCM2 specific for an RG I-associated epitope (Puhlmann et al. 1994).
Thin sections (90–95 nm) mounted on gold grids were incubated in a blocking solution of 3% non-fat dried milk in PBST 0.1% (10 mM Na-phosphate (Sigma), 500 mM NaCl (Sigma), 0.1% Tween-20 (Sigma) for 30 min. All incubations were carried out overnight at 4C, or for 4 hr at room temperature as previously described (Vicré et al. 1998). Grids were washed in PBST, dried, and incubated on droplets for undiluted JIM5, LM6, or LM5 (diluted 1:5 in PBST) and anti-PGA/RG I (diluted 1:10 in PBST). After washing with PBST 0.5%, the grids were transferred to (a) a droplet of goat anti-rat IgG conjugated to 20-nm colloidal gold (Biocell) diluted 1:25 in PBST (for JIM-5, LM-5, and LM6-labeled sections) or to (b) a goat anti-rabbit IgG conjugated to 20-nm colloidal gold (Biocell) solution diluted 1:25 in PBST (for anti-PGA/RG I-labeled sections) for 1 hr at room temperature. The sections were first rinsed with PBST (0.5%), followed by distilled water. After immunolabeling, the sections were submitted to a mild PATAg staining as previously described (His et al. 1997; Jauneau et al. 1997). These sections were examined at 80 kV on either a Zeiss EM109 or a Philips Tecnai 12 electron microscope. All micrographs illustrate representative labeling patterns. Controls were performed either by omission of primary antibodies or by pre-incubation of the probe for 1 hr with polygalacturonic acid (1 mg/ml) for JIM5, anti-PGA/RG I, and the cationic gold probe.
Results
Fiber Differentiation
Figure 1 shows the average of stem length at different growth stages of flax plants. After a period of active growth (42–80 days; 5–85 cm), the elongation of the stem slowed down at the onset of floral bud development (80–98 days; 85–90 cm; see also Morvan et al. 1989, 1990). At maturity (~100 days), the plants usually have a stem length of 90–110 cm.
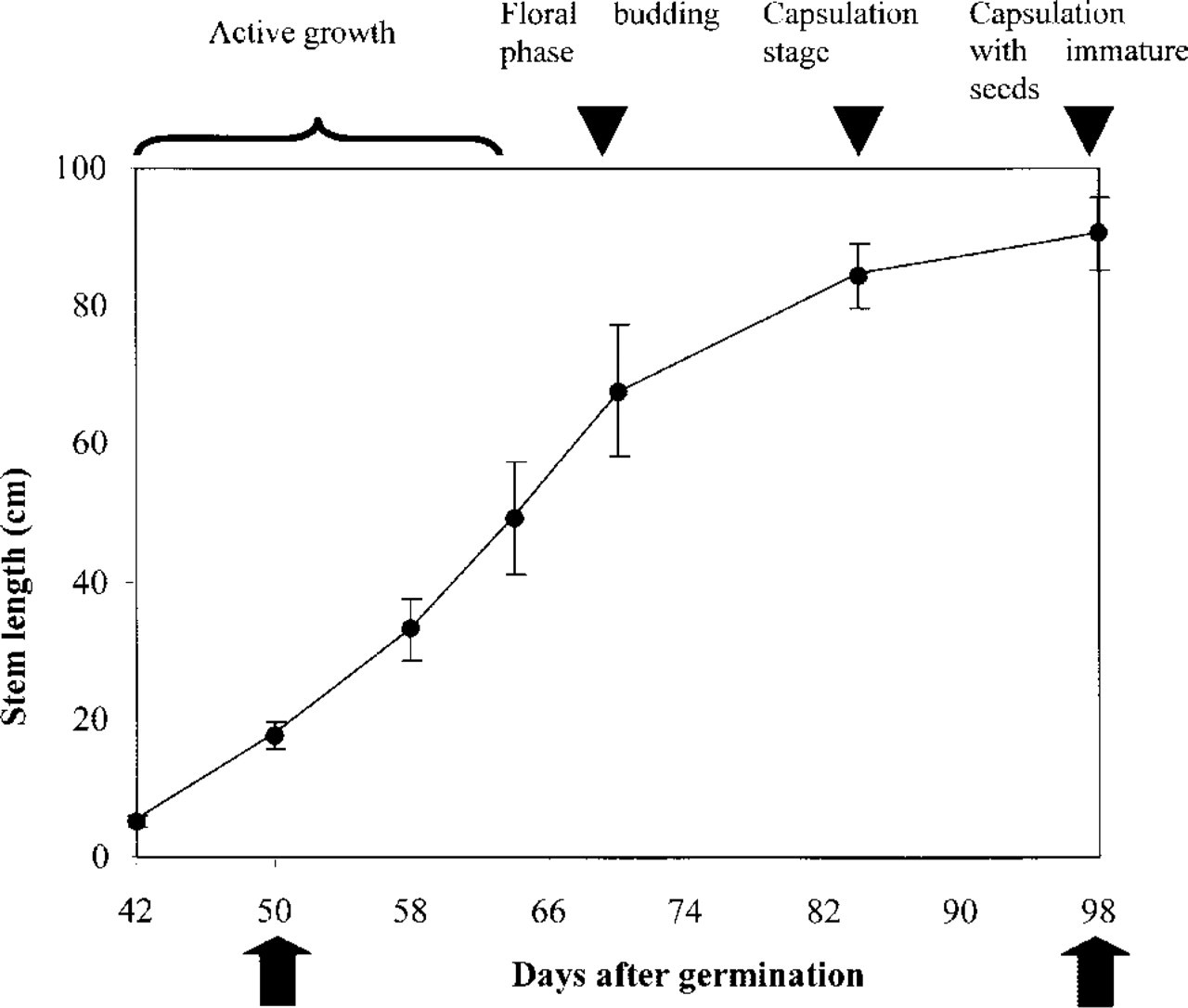
Growth rate of flax plants. The plants were grown in the field. The stem length of 100 freshly harvested plants from different stages of development was measured. Black arrowheads denote different developmental stages of flax. Arrows indicate the two specific stages from which plants in this study were used.
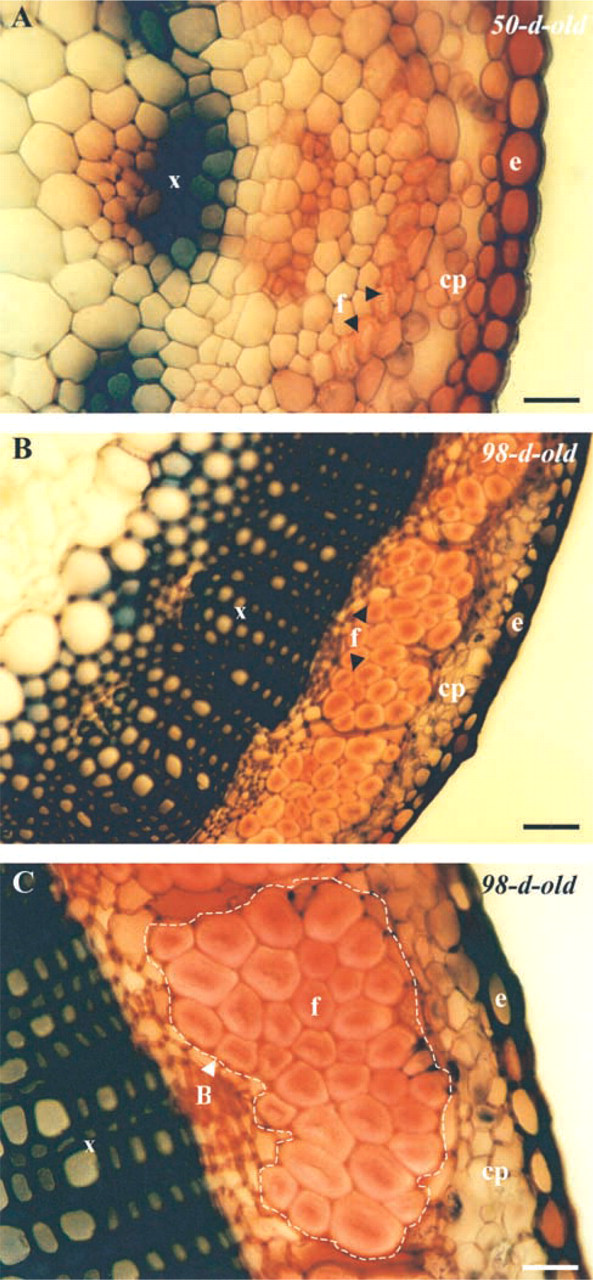
Hand-cut cross-sections from fresh stems stained with safranin, showing the organization of the tissue in 50-day (
Fiber differentiation, characterized by deposition of a cellulose-rich secondary wall, starts very early during flax development (Esau 1977; His 1999; Andème-Onzighi et al. 2000) and continues throughout all plant growth stages. The filling of the fibers occurs progressively during development and, at maturity, fiber cells have a very thick secondary wall (Esau 1977).
In this study, plants were field-harvested at two stages of development, i.e., 50 and 98 days, corresponding to two stages of fiber differentiation. Hand-cut transverse sections from fresh stems, stained with safranin-light green, showing the anatomic organization at both stages are seen in Figure 2. In 50-day-old plants, differentiating fibers (not mature) are arranged in one layer at the periphery of the stem (Figure 2A). They have a relatively thick secondary wall and a wide lumen. In 98-day-old plants, mature fibers (Figures 2B and 2C), are organized into bundles, with up to 20 cells per bundle, located between the cortical parenchyma (cp) and the vascular cylinder, which consists mostly of xylem cells (x). The secondary wall is remarkably thick and the lumen width has undergone a pronounced reduction.
Ultrastructure of LRW-embedded Stems
To improve the quality of ultrastructural preservation of mature fibers and to perform immunogold labeling, we devised a modified method based on a vacuum fixation combining both formaldehyde and glutaraldehyde to enhance the rate of penetration. Vacuum was also applied during early stages of resin infiltration, and LRW embedding duration was increased, especially in the later stages (see Discussion).
To check for the structural preservation of fibers at both stages of maturity after fixation and LRW-embed-ding, stem sections were examined using both light and transmission electron microscopy. As illustrated in Figures 3A and 3B, transverse semithin sections stained with methylene blue-basic fuchsin reveal well-preserved tissues, including xylem and fiber cells. The good quality of preservation is confirmed by transmission electron microscopy (TEM) observations of 90-nm ultrathin sections (Figures 3C and 3D). It is interesting to note that the cohesion of fiber bundles in chemically fixed and LRW-embedded stems is well maintained as in the native state (compare with hand-cut transverse sections of fresh, non-fixed tissue in Figure 2). The primary and secondary cell walls (Figure 3D), as well as cell junctions and the narrow lumen of fibers are clearly distinguishable (Figure 3D).
After checking the preservation quality of fibers, we proceeded to gold labeling of pectic polysaccharides using different gold probes.
Cell wall Localization of Anionic Sites
The poly-
Immunogold Labeling of Homogalacturonans
To check for the presence of homogalacturonans and to examine their distribution in fiber walls, we used the MAb JIM5, which recognizes pectin epitopes containing up to 40% of methylesterified PGA (Knox et al. 1990; Willats et al. 2000). The antibody binds almost not at all to fully de-esterified pectins. As seen in Figure 5, labeling with this probe is confined to the middle lamellae and the cell junctions. No labeling is seen over the primary or the secondary cell walls. Similar data were also obtained with anti-PGA/RG I polyclonal antibodies, which recognize pectin epitopes consisting of de-esterified PGA domains (Moore and Staehelin 1988; Lynch and Staehelin 1992) (Figure 5C). In contrast, when labeling was performed with JIM7, an MAb that binds to methylesterified pectin epitopes with a degree of esterification ranging between 15 and 81% (Knox et al. 1990; Willats et al. 2000), no gold particles were detected anywhere in the fiber walls (not shown), suggesting that the JIM7 epitope is either absent or not accessible to the antibody.
Immunolocalization of Rhamnogalacturonan I-associated Epitopes
Complex polysaccharides such as RG I have been reported to be present in the primary walls of flax tissues including fibers (Davis et al. 1990). In this study, we have taken advantage of three antibodies specific for RG I, i.e., CCRCM2, LM5, and LM6, and the good preservation of the tissues to examine whether these polysaccharides are present and to map their distribution in mature fibers. Labeling of the fiber cells with the MAb CCRCM2, recognizing an epitope of the RG I backbone (Puhlmann et al. 1994), differed between 50- and 98-day-old plants (Figure 6). Whereas it was absent from the secondary wall and mostly associated with the cytoplasm, probably with Golgi-derived vesicles, in 50-day-old samples (Figure 6A), it was abundant all over the secondary wall in more mature fibers (Figure 6B). This indicates a possible developmental regulation of the CCRCM2-epitope deposition in the wall of fibers.
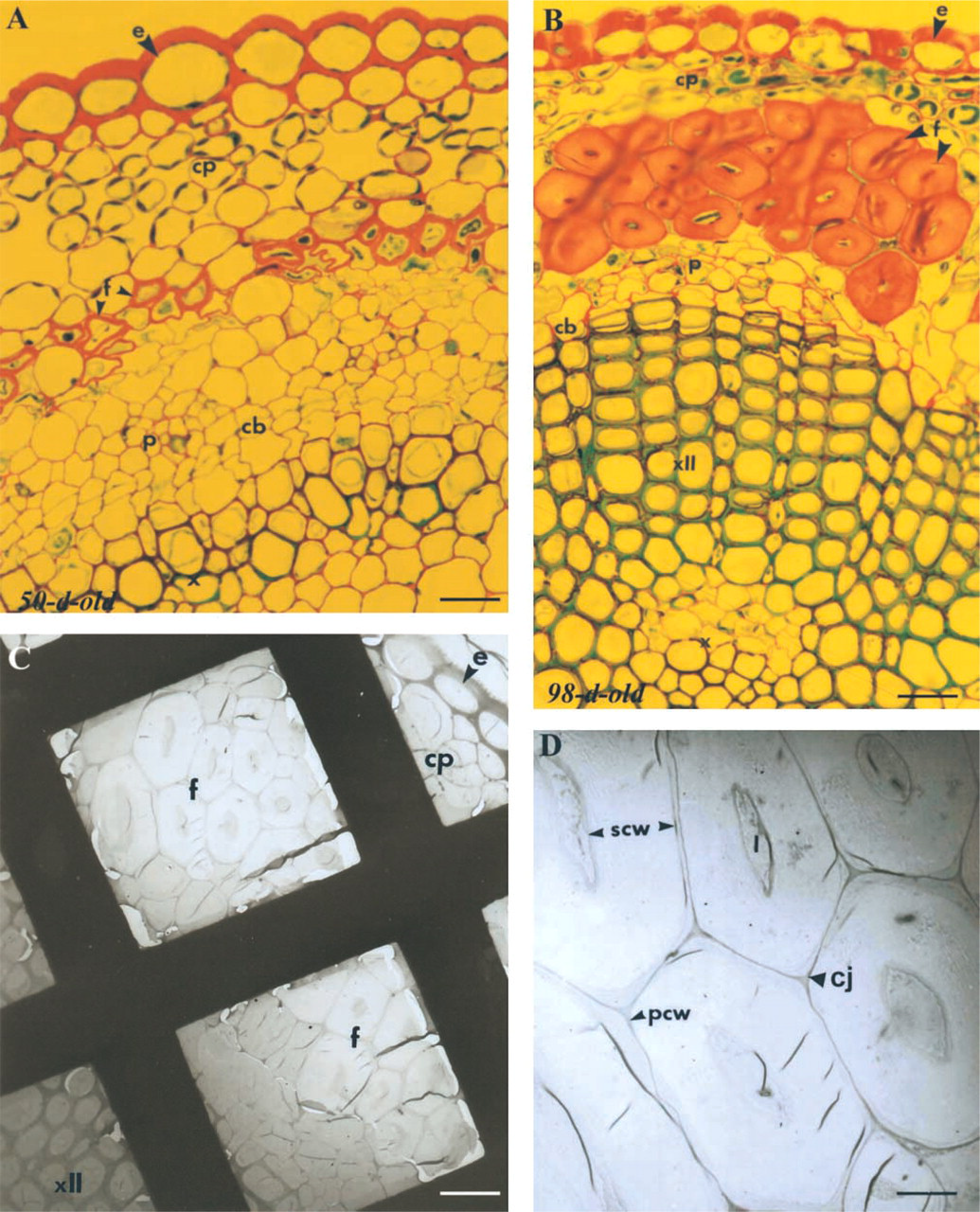
Light and electron micrographs of LRW-embedded fiber bundles. Stem cross-sections of 50-day- (
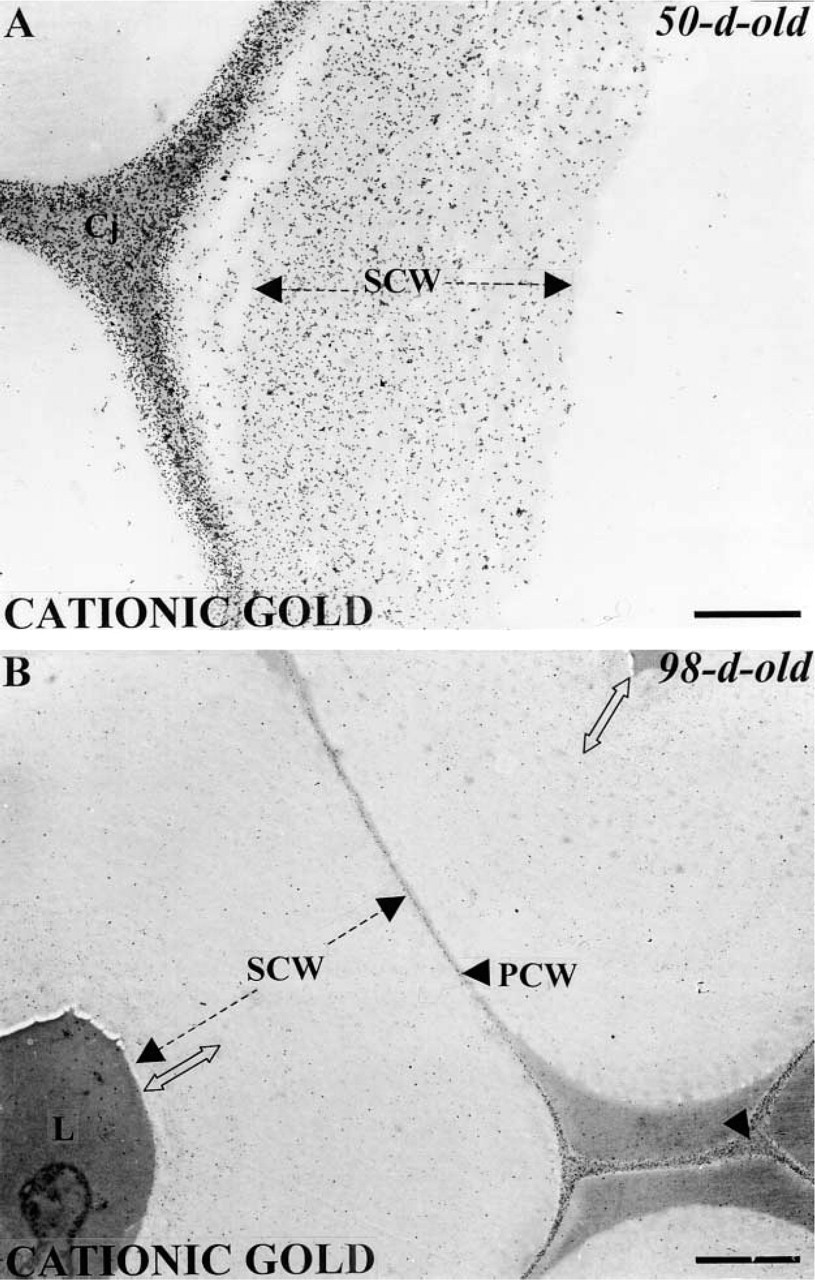
Localization of anionic sites with the cationic gold probe in mature fiber walls. The secondary walls are heavily and uniformly labeled in 50-day-old plants (
Labeling of fiber walls with LM5 and LM6, two MAbs that recognize (1–4)-β-
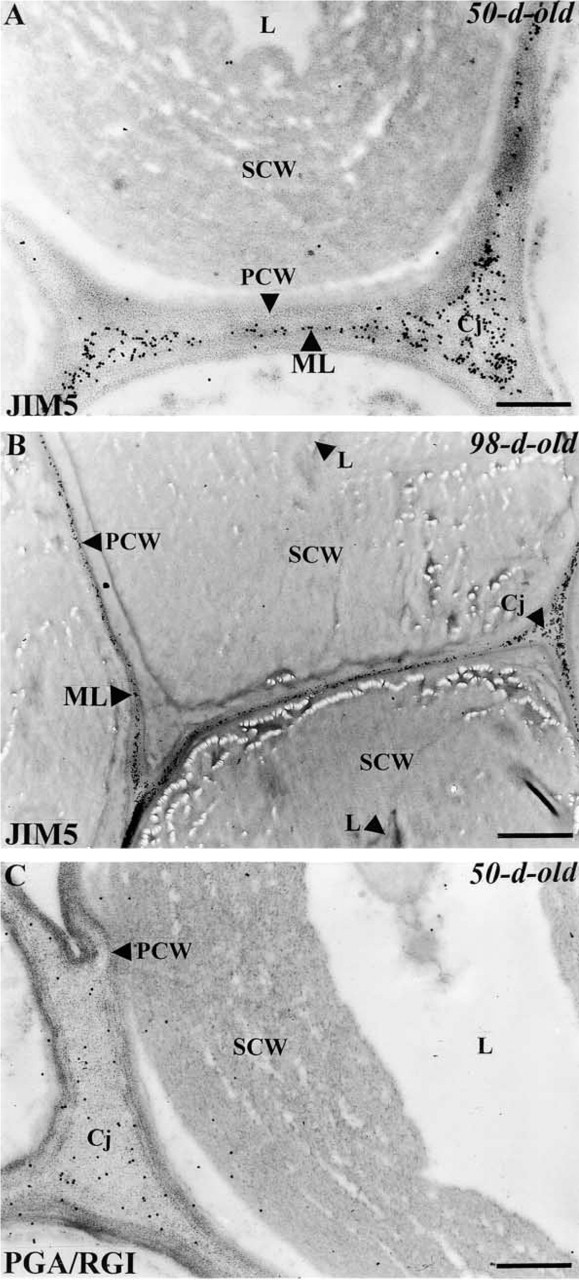
Immunogold labeling of homogalacturonans in mature fibers with JIM5 (
Control experiments performed with the omission of all the primary antibodies recognizing pectins show no labeling over fiber cell walls (data not shown).
Discussion
The goal of the present study was to find a tissue preparation and embedding method that could be applied to mature flax fibers so that they could be used in immunogold microscopy. Working on stems harvested at two growth stages (i.e., rapid phase of growth and maturation) of flax plants made it possible to show that several polysaccharides of the pectic class are present in different cell wall domains and undergo developmental changes during bundle formation and fiber maturation.
Preservation of Fiber Cell Walls
In the past we have been able to achieve good preservation of mature fibers embedded in Spurr's resin (Jauneau et al. 1994,1995; Bert et al. 1996), but immunogold labeling remained problematic. Recently, it was also possible to achieve good preservation of young flax fibers embedded in LRW, which allowed an ICC characterization of their wall components (His 1999; Andème-Onzighi et al. 2000). Unfortunately, the application of the same method to mature fibers was not satisfactory. The structural preservation was poor, resulting in significant separation between primary and secondary wall and loss of stability of fiber bundles during sectioning and under the electron beam. This is mostly due to the organization of individual fiber cells into bundles, in which they are tightly associated one to another, creating a very hard compact tissue compared to the parenchyma or xylem cells, and to the narrow lumen and the thick secondary wall, which tend to resist resin penetration. Recently, good ultrastructural preservation was difficult to achieve even on non-completely mature fibers (later stage of fast growth) using the classical fixation/embedding method (Gorshkova et al. 2000). By comparison, other secondary wall-containing tissues, such as xylem, are less difficult to prepare because of the large lumen and thinner secondary wall, and there are many reports of TEM studies on xylem cells (Shindler et al. 1995; Ruel et al. 1999). To facilitate infiltration and improve the preservation of very hard tissues, Roland and Vian (1991) recommended the use of extractions with chemicals or enzymes before postfixation and embedding. We did not apply such a technique to flax fibers because wall components such as pectins might also be removed and therefore their distribution could not be studied.
Therefore, to circumvent the difficulties encountered, a modification of the classical method, based mainly on an extended vacuum infiltration/fixation step before embedding in LRW resin, was used. This enabled flax fibers, which are reputed to be difficult to infiltrate, to be successfully and homogeneously embedded in LRW. The fixation definitely yielded better preservation of fiber bundles than that obtained with the basic method used for young fibers, and enhanced their structural stability. Although some minor alterations such as cracks were apparent, the secondary walls were uniformly and reproducibly well preserved.
Location of Wall Polymers
By in situ localization using different gold probes, we show that there are differences in the distribution of polysaccharides within the cell walls of mature fibers (see diagram in Figure 7). A large amount of cationic gold labeling is seen within the secondary wall and all other wall domains of fibers at both stages of differentiation. The specificity of the cationic gold probe was previously characterized. It was shown to bind mainly to the COO- groups of galacturonic acid of un-esterified pectins, although it can also recognize other acidic molecules such as proteins or phenolics (Roy et al. 1994). Thus it appears that mature fibers might contain substantial amounts of pectins as has been shown by biochemical studies (Morvan et al. 1989,1990; Davis et al. 1990; Gorshkova et al. 1996). JIM5 and anti-PGA/RG I were then used to locate homogalacturonans (HGs). JIM5 antibodies were characterized recently and were shown to bind weakly to fully de-esterified pectins but methylesterified GalA residues up to 40% enhanced binding (Willats et al. 2000). Whereas PGA/RG-I antibodies were shown to recognize unesterified domains of PGA (Lynch and Staehelin 1992). No labeling was detected in the primary and secondary walls of fibers with both probes suggesting that HGs are poorly represented in flax fibers. In contrast, the observed intense binding to cell corners and middle lamellae indicates that these domains are mainly composed of HGs with either methyesterified and unesterifed PGA domains. This was not surprising as HGs have the capacity to bind calcium ions thereby forming calcium pectates, which ensure the intercellular cohesion of adjacent cells (Goldberg et al. 1996). Calcium ions were abundantly detected in these regions in both young and mature fibers (Jauneau et al. 1992; Ripoll et al. 1993; His et al. unpublished results). In contrast, the segregation of the location for cationic gold and JIM5/PGA-RG I binding was surprising because these two probes should target the same component. This suggests that sensitivity of cationic gold to galacturonic acids of HGs is probably higher than that of the antibodies. However, the difference in labeling might also be explained by differences in the steric environment created by each probe. Thus, the limited labeling observed by the antibodies may result from the greater steric environment created by the primary and the secondary antibody complexes. Alternatively, PGA/RG I and JIM5 epitopes may have been masked through interaction of pectins with other polymers. Although we do not have direct evidence, interaction of pectins with other encrusting components, such as arabinogalactan proteins, or even with cellulose is likely to occur within fiber secondary walls (Girault 1999).
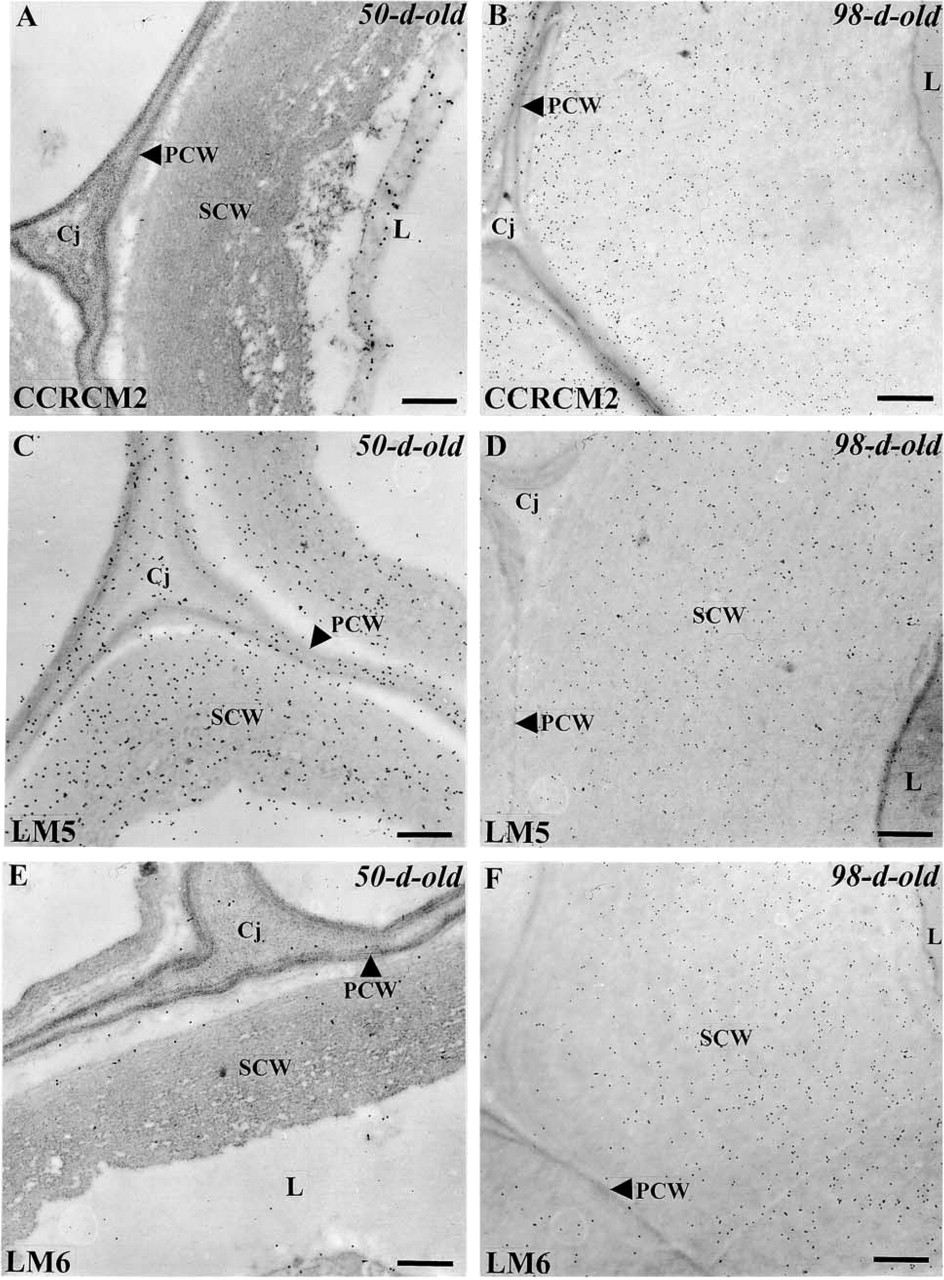
Immunogold localization of epitopes associated with the polysaccharide rhamnogalacturonan I with CCRCM2 (
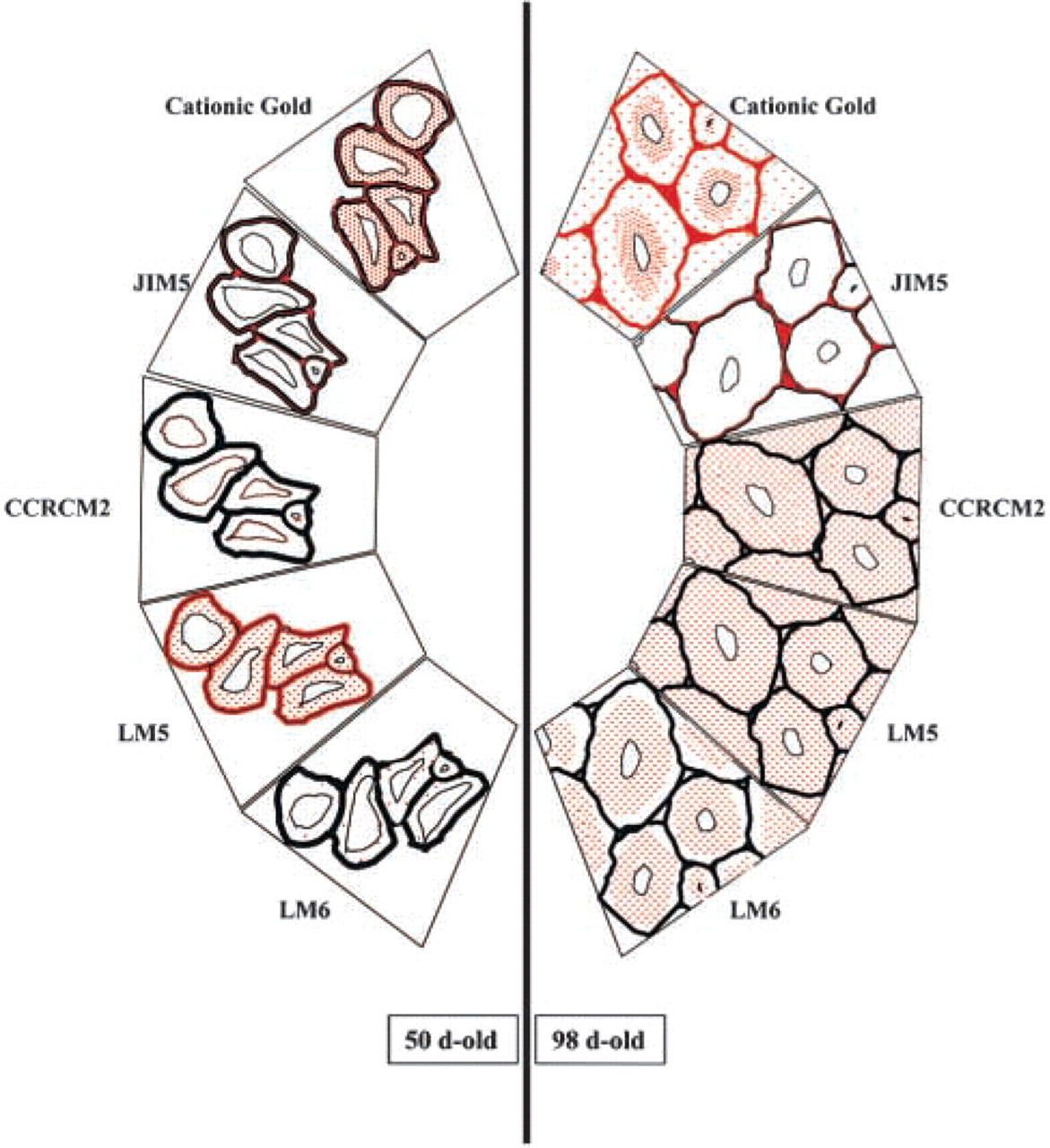
Summary diagram showing the distribution of anionic sites and pectic epitopes in the cell walls of flax fibers at two stages of development (50- and 98-day-old plants). Gold labeling is shown in red. Dotted red represents the labeling found in the secondary wall. Solid red represents the labeling found in the primary cell wall, cell junctions and middle lamellae. Each box represents a section of a fiber bundle. In 50-day-old plants, anionic sites are detected in the primary and secondary walls as well as in the cell junctions and the middle lamellae. JIM5 epitopes are detected only in the middle lamellae and cell junctions but are not detectable within the secondary cell wall. RG I epitopes recognized by the MAb CCRCM2 are found in the cytoplasm but not in the walls. LM5 and LM6 epitopes are detected in the secondary wall. The labeling with MAb LM5 is much more intense than that observed with LM6. In 98-day-old plants, anionic sites and JIM5 epitopes have a similar wall distribution as in 50-day-old plants. Anionic sites are mainly concentrated in the inner part of the secondary wall. CCRCM2, LM5, and LM6 epitopes are all detected in the secondary wall of fiber cells, although with slightly different locations.
The undefined RG I epitope recognized by CCRCM2 was not detected in the fiber walls during the rapid phase of growth, whereas it was abundantly present in the secondary wall at maturity. The absence of labeling in the cell walls of differentiating fibers might be due to the lack of RG I or to a substitution, such as acetyl-esterification, which is not recognized by the antibody. The fact that labeling is reproducibly detected in the cytoplasm indicates that the epitope is synthesized and possibly secreted into the cell wall. Therefore, RG I pectin is probably present in the wall but the epitope could not be detected. RG I polymers decorated with short side chains of galactosyl residues, which are partially acetylesterified, were found in flax fibers (Davis et al. 1990). In addition, indication of the presence of acetylesterified pectins in fiber walls was also obtained using ferric hydroxylamine staining on ultrathin sections (Jauneau et al. 1994). Therefore, we believe that acetylesterification of RG I pectin may contribute at least partially to the lack of labeling with the CCRCM2 antibody in 50-day-old plants. As bundles were formed and fibers differentiated, the secondary walls became highly labeled with CCRCM2 antibody. This also appeared to happen with the LM6 antibody, which could reflect an enrichment of mature fibers with RG I-containing α-1-5-arabinans in their side chains. β-1-4-galactans (recognized by LM5) may also be part of the side chain of RG I present in mature fiber secondary walls. However, the presence of free galactans and arabinans in the secondary wall cannot be excluded. Girault et al. (1997) and Girault (1999), using 13C NMR and methylation analyses, have clearly established that the encrusting components of the cellulose microfibrils of secondary walls of mature fibers are mainly composed of galactosyl residues arranged in long 1–4-β-
Footnotes
Acknowledgments
We gratefully acknowledge the gift of some of the antibodies from Drs M. Hahn, P. Knox, and L.A. Staehelin. We also thank Drs M. Jarvis and M. Vicré for critical reading of the manuscript.