
Abstract
Cadherin-based molecular complexes play a major role in cell-cell adhesion. At the adherens junctions the intracellular domain of cadherins specifically interacts with β-catenin and p120ctn, members of the Armadillo repeat protein family. Differential splicing and utilization of the alternative translation initiation codons lead to many p120ctn isoforms. Two major p120ctn isoforms are expressed in mouse tissues. In this study we used indirect immunofluorescence to demonstrate significant tissue specificity in expression of the p120ctn isoforms. The short isoform is abundant at cell-cell adhesion junctions in epidermis, palatal, and tongue epithelia, in the ducts of excretory glands, bronchiolar epithelium, and in mucosal epithelia of esophagus, forestomach, and small intestine. In contrast, the long isoform, containing an amino terminus highly conserved within the p120ctn subfamily, is expressed at vascular-endothelial cell junctions in blood vessels, at cell-cell junctions in the serosal epithelium lining the internal organs, in choroid plexus of brain, in the pigment epithelium of retina, and in structures such as the outer limiting membrane of retina and intercalated discs of cardiomyocytes. The tissue- and cell type-specific expression of p120ctn isoforms suggests a role for the long p120ctn isoform in cell structures responsible for stable tissue integrity, compared to the role of the short isoform in cell-cell adhesion in the external epithelia with rapid turnover.
C
Several binding sites have been mapped to different domains of p120ctn. The Armadillo domain interacts with the intracellular juxtamembrane domain of classical cadherins within the adherens junctions (Daniel and Reynolds 1995; Reynolds et al. 1996), and also with Kaiso, a POZ-ZF transcription factor (Daniel and Reynolds 1999). Originally p120ctn was identified as a major substrate for tyrosine phosphorylation (Reynolds et al. 1989). The amino terminus of p120ctn is responsible for the interaction with the cytoplasmic tyrosine kinase FER (Kim and Wong 1995). In addition, the RPTPμ-interacting domain of p120ctn maps to its unique amino terminus, a region distinct from the cadherin-interacting domain (Zondag et al. 2000). The p120ctn isoform 3A was identified as a main interaction partner of the protein-tyrosine phosphatase SHP-1 (Keilhack et al. 2000). The amino terminus of isoform 3 interacts with the amino-terminal globular domain of BPAG2/BP180/Type XVII collagen, a transmembrane protein primarily localized to hemidesmosomal attachment complexes in basal keratinocytes (Aho et al. 1999).
The p120ctn can induce a prominent branching phenotype in cells in culture (Reynolds et al. 1996). The small GTPases of the Rho subfamily are signaling molecules that are primarily involved in remodeling of the cytoskeleton. The interaction of p120ctn with Rho-family GTPases has been recently described (Anastasiadis et al. 2000; Noren et al. 2000). Therefore, p120ctn can regulate cell motility through the actin cytoskeleton via Rho-family GTPases.
Analysis of p120ctn isoforms in a variety of murine cell lines has revealed that motile cells, such as fibroblasts and macrophages, preferentially express the long isoform and epithelial cells preferentially express the short isoform, whereas non-adherent cells do not express detectable levels of p120ctn (Mo and Reynolds 1996). The splicing pattern of the p120ctn transcript is tissue-specific and consistent between each fetal and adult human tissue when studied by an RT-PCR approach (Aho et al. 1999). Analysis of a variety of human tumor cell lines has revealed that p120ctn isoform expression in these cells is heterogeneous and does not reflect the types of isoforms expressed by the tissues or cell types from which each tumor cell line originated (Mo and Reynolds 1996; Aho et al. 1999). MAb pp120, which recognizes all p120ctn isoforms, has recently been used to demonstrate that p120ctn is widely distributed in the mammalian tissues in epithelia, endothelia, cardiomyocytes, and the nervous system (Golenhofen and Drenckhahn 2000). Another set of MAbs that recognize the extreme amino terminus of p120ctn can be used to distinguish the long p120ctn isoform 1 from the entire population of p120ctn isoforms (Wu et al. 1998). We utilized MAbs 6H11 and pp120 to investigate the tissue distribution and cell type-specific expression of p120ctn isoform 1 compared to the entire p120ctn population in mouse tissues.
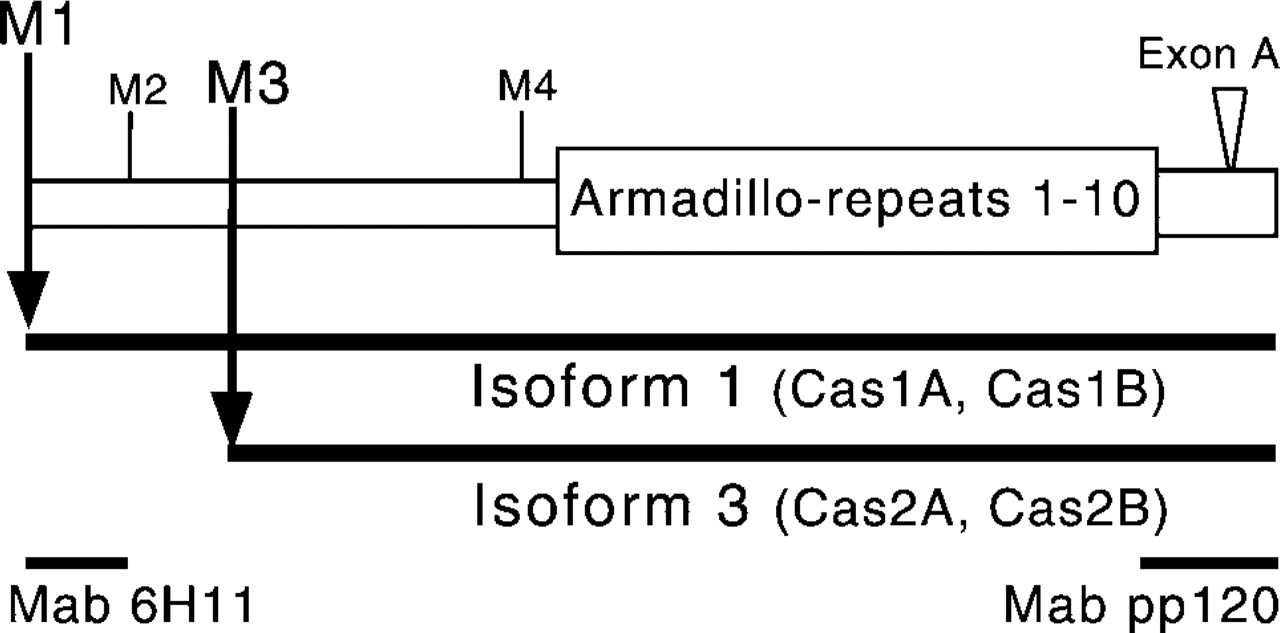
Schematic structure of p120ctn highlighting isoforms 1 and 3. The alternative splicing of 5′-exons gives rise to multiple p120ctn isoforms. In mouse, p120ctn isoform 1 (previously called Cas1B) starts from Met-1 (M1) and isoform 3 (previously called Cas2B) starts from Met-3 (M3). Two additional isoforms, 2 and 4, starting from Met-2 (M2) and Met-4 (M4), have been observed in human cells. At the C-terminus, sequences encoded by the alternatively spliced exon A give rise to additional murine isoforms, previously called Cas1A and Cas2A. MAb 6H11 detects the amino terminus of the isoform 1, while MAb pp120 recognizes a C-terminal epitope common to all p120ctn isoforms.
Materials and Methods
Western Blotting
The tissues and organs were dissected from CO2-asphyxiated mice and immediately homogenized by a Polytron homogenizer into the SDS-gel loading buffer but omitting the tracking dye (100 mg of tissue/ml). The insoluble residue was separated by centrifugation at 10,000 × g for 30 min. The protein concentration was assayed using Bio-Rad Protein Assay reagent (Bio-Rad; Hercules, CA). The tissue ly-sates were mixed 1:1 with SDS sample buffer, proteins separated on 8% SDS-PAGE, and transferred onto PVDF membrane (New England Nuclear Life Science; Boston, MA). The pp120 monoclonal antibody (250 μg/ml) was purchased from Transduction Laboratories (Lexington, KY) and used at 1:2000 dilution. The p120ctn amino terminus-specific MAb 6H11 (Wu et al. 1998) was a generous gift from Dr. A. Reynolds (Vanderbilt University, Nashville, TN). MAb 6H11 (530 μg/ml) was used at 1:500 dilution for Western blotting. Anti-mouse secondary antibody (0.8 mg/ml, used at 1:5000 dilution) was purchased from Jackson Laboratories (West Grove, PA). The secondary antibody, conjugated to HRP, was detected with Renaissance Western blot chemiluminescence reagent (New England Nuclear).
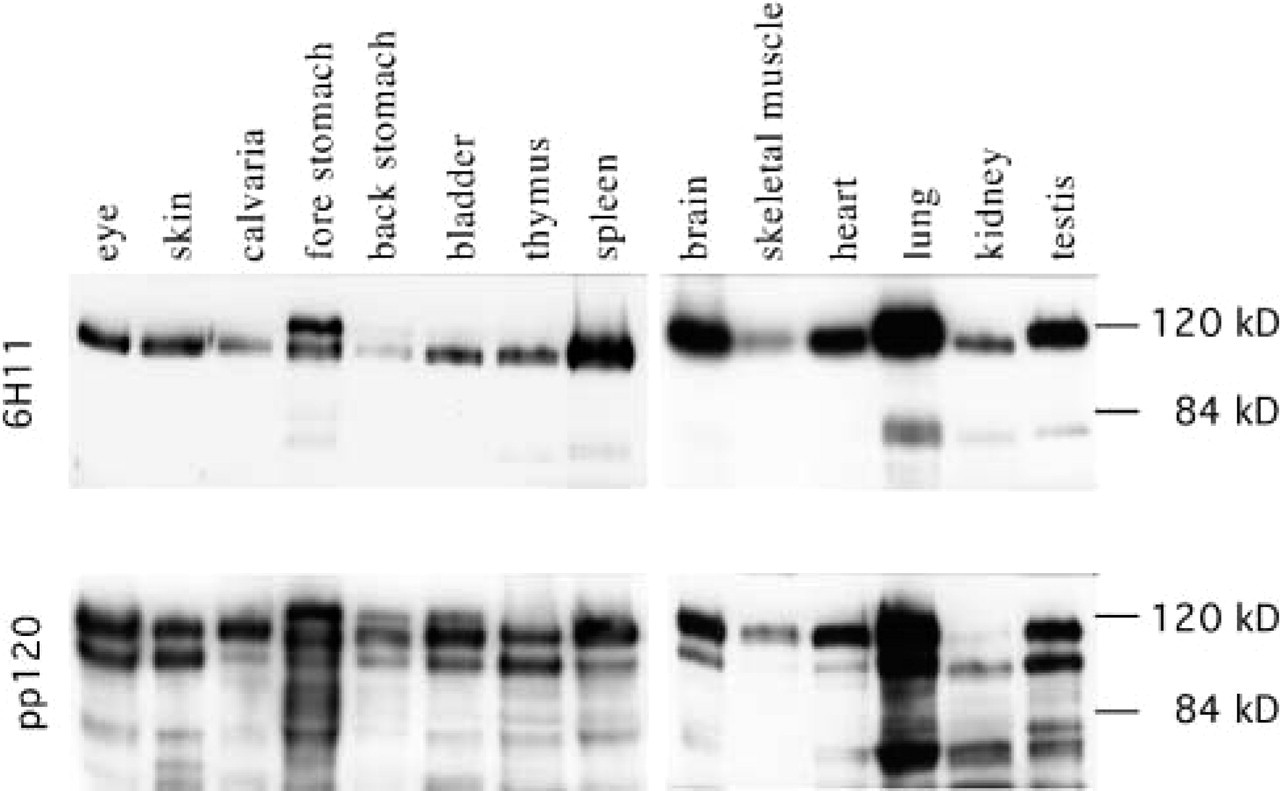
Immunoblotting demonstrates two major p120ctn isoforms in mouse tissues. MAb 6H11 recognized a 120-kD band from all the tissues analyzed. A more slowly migrating band was detected from mouse stomach and an 80-kD band from lung tissue. MAb pp120 revealed a second major polypeptide of 100 kD and several minor bands, which may represent various isoforms and their post-translational modifications or degradation products.
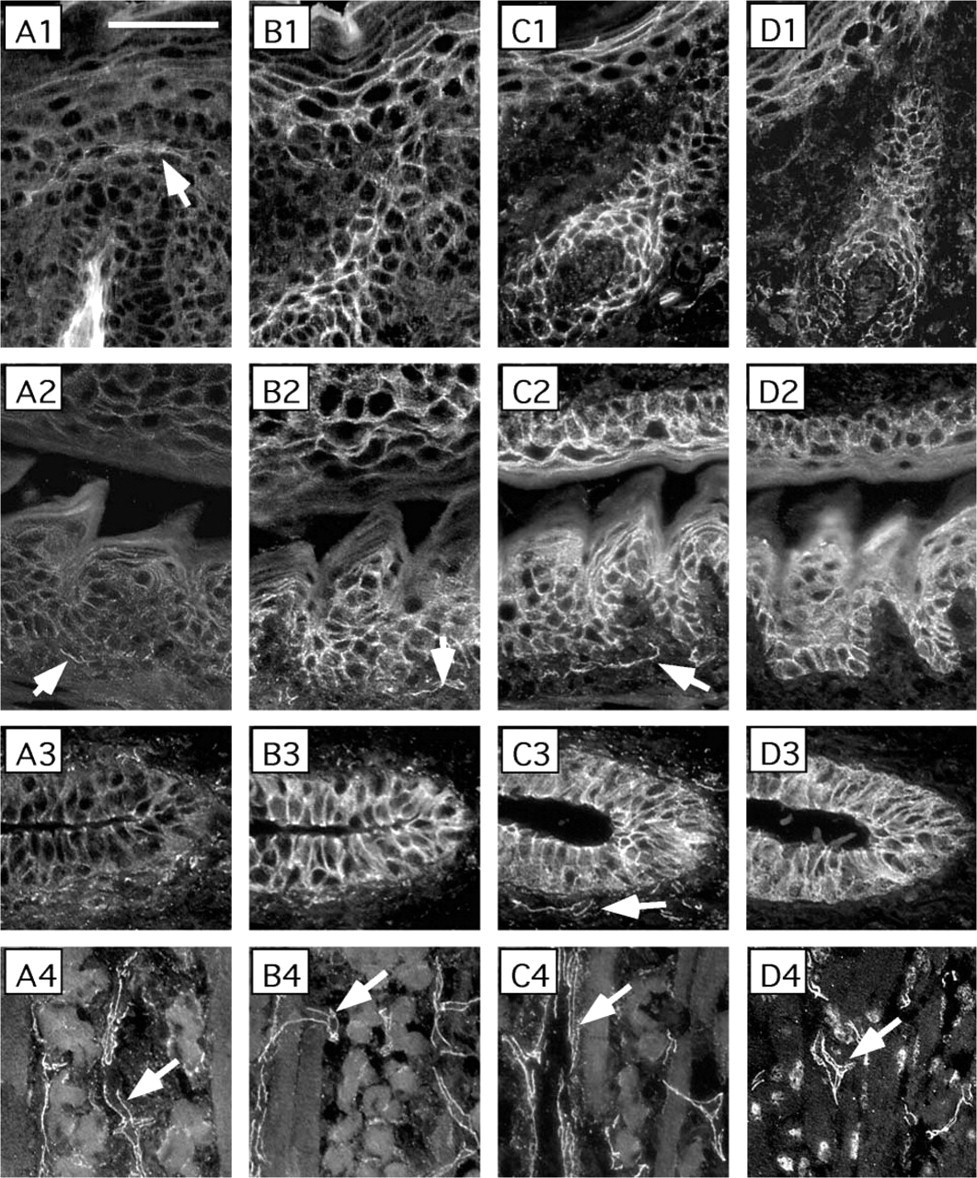
p120ctn isoform 3 localizes to the cell-cell junctions in neonatal mouse epithelia. Adjacent sections of the epidermis of scalp and a hair follicle (Row 1), epithelium of palate and tongue (Row 2), an excretory duct of a salivary gland (Row 3), and tongue muscle (Row 4) were subjected to indirect immunofluorescence with primary antibodies specific for the p120ctn isoform 1 (MAb 6H11, Column
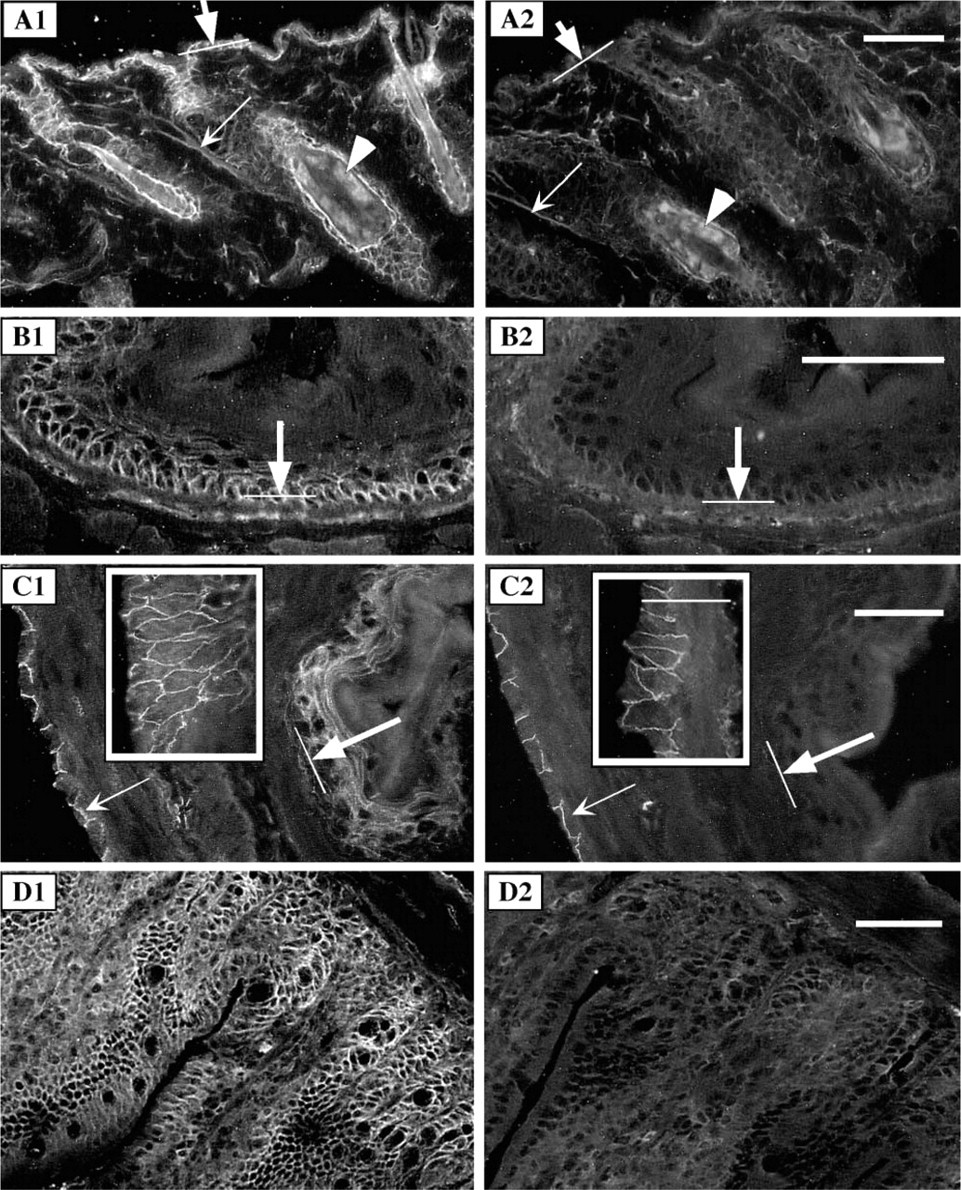
p120ctn isoform 3 is present in adult mouse epidermis and in the epithelia lining the gastrointestinal tract. Parallel sections of adult mouse skin (
Immunocytochemistry
For preparation of cryosections, the tissues from CO2-asphyxiated mice were removed, embedded in TBS Tissue Freezing Medium (Triangle Biomedical Sciences; Durham, NC), and rapidly frozen in liquid nitrogen. Tissue blocks were mounted in TBS and sections 7 μm thick were cut, air-dried, and stored at −20C if not processed immediately. The sections were fixed in methanol at −20C for 5 min, rinsed twice in PBS, and permeabilized with 0.1% Triton X-100 in PBS for 5 min at room temperature (RT). The sections were then washed three times with PBS, blocked with 1% BSA (bovine serum albumin) in PBS for 1 hr at RT, and incubated with each primary antibody in PBS-1% BSA overnight at 4C. For indirect immunofluorescence, the pp120 antibody was used at 1:1000 dilution and the 6H11 antibody at 1:500 dilution. Mouse MAbs for β-catenin (250 μg/ml, diluted 1:500), E-cadherin (250 μg/ml, diluted 1:2000), and N-cadherin (250 μg/ml, diluted 1:500) were purchased from Transduction Laboratories. A goat polyclonal antibody for VE-cadherin (IgG, 200 μg/ml, diluted for slides 1:200) was from Santa Cruz Biotechnology (Santa Cruz, CA). After four washes with PBS, the sections were incubated for 1 hr at RT with Texas Red dye-conjugated affinity-purified donkey anti-mouse or anti-goat IgG (1.4 mg/ml) (Jackson) diluted 1:500 in PBS-1% BSA. Parallel sections of each tissue were processed using mouse IgG (Chemicon International; Temecula, CA) in place of the primary antibody (1 μg/ml, which was the highest concentration of any primary antibody used; data not shown). As a negative control for the goat VE-cadherin antibody, another goat antibody not recognizing the vascular-endothelial proteins was used. To visualize the nuclei, 0.00002% DAPI was included in the secondary antibody incubation. Slides were washed four times with PBS, dehydrated with dipping in 95% ethanol, and mounted for viewing. Sections were studied under a fluorescent microscope (Axioskop; Carl Zeiss), images were captured with ImagePro Plus 4.0 imaging software (Media Cybernetics; Tucson, AZ) and processed with Photoshop 5.0 (Adobe Systems; Tucson, AZ) and Canvas 5 (Deneba Software; Miami, FL).
Results
p120ctn Is Expressed as Multiple Isoforms in Mouse Tissues
Differential splicing and alternative ATG codons give rise to multiple p120ctn isoforms starting with the initiation methionines M1-M4 (Figure 1). In human cells, RT-PCR studies have revealed alternatively spliced transcripts, which can give rise to p120ctn isoforms 1–4 (Keirsebilck et al. 1998; Aho et al. 1999). In mouse cells, two major p120ctn isoforms, corresponding to human isoforms 1 and 3, have been observed (Mo and Reynolds 1996). Both murine isoforms have been identified either with or without a short carboxyl terminal exon A (Mo and Reynolds 1996). Therefore, in mouse cells four isoforms, corresponding to human isoforms 1A, 1, 3A, and 3 have been detected. MAb pp120 has been raised against p120ctn carboxyl terminal amino acids 790–911 and recognizes all known p120ctn isoforms. MAb 6H11 was raised against the amino-terminal region of p120ctn, amino acids 8–349 (Wu et al. 1998) and was later found to recognize specifically isoform 1 of p120ctn (unpublished results; and Daniel and Reynolds 1999).
Western blotting analysis of a set of mouse tissue homogenates confirmed that p120ctn is expressed tissue-wide (Figure 2). In all tissues studied, MAb 6H11 detected a 120-kD band, corresponding to the mouse Cas1B (Mo and Reynolds 1996) and to the human isoform 1 (Keirsebilck et al. 1998). Mouse forestomach contained also a more slowly migrating band, corresponding to the previously characterized mouse Cas1A (Mo and Reynolds 1996) and to the human isoform 1A (Keirsebilck et al. 1998). A more rapidly migrating band around 80 kD, which was detected from mouse lung homogenate, may be a degradation product of p120ctn isoform 1. When detected with the pan-p120ctn-specific antibody pp120, two major isoforms, Cas1 and Cas2 corresponding to human isoforms 1 and 3, were equally represented in eye, skin, bladder, thymus, lung, and testis, whereas Cas1/isoform 1 was dominant in calvaria, spleen, brain, skeletal muscle, and heart (Figure 2, lower panel). These results are in agreement with our previous observation, in which we utilized RT-PCR to demonstrate that the isoform 1 transcript is prominent in human brain, heart, spleen, testis, and ovary (Aho et al. 1999). Because many cell types are present and form distinct functional structures in each tissue or organ, the isoform specificity in situ can be best demonstrated through indirect immunofluorescence analysis.
p120ctn isoform 1 is a component of internal epithelia and stable cell-cell junction of non-epithelial cells. Sections of adult mouse brain (
The p120ctn Isoform 3 (Cas2) Is Present at the Intercellular Junctions of Stratified Squamous Epithelia
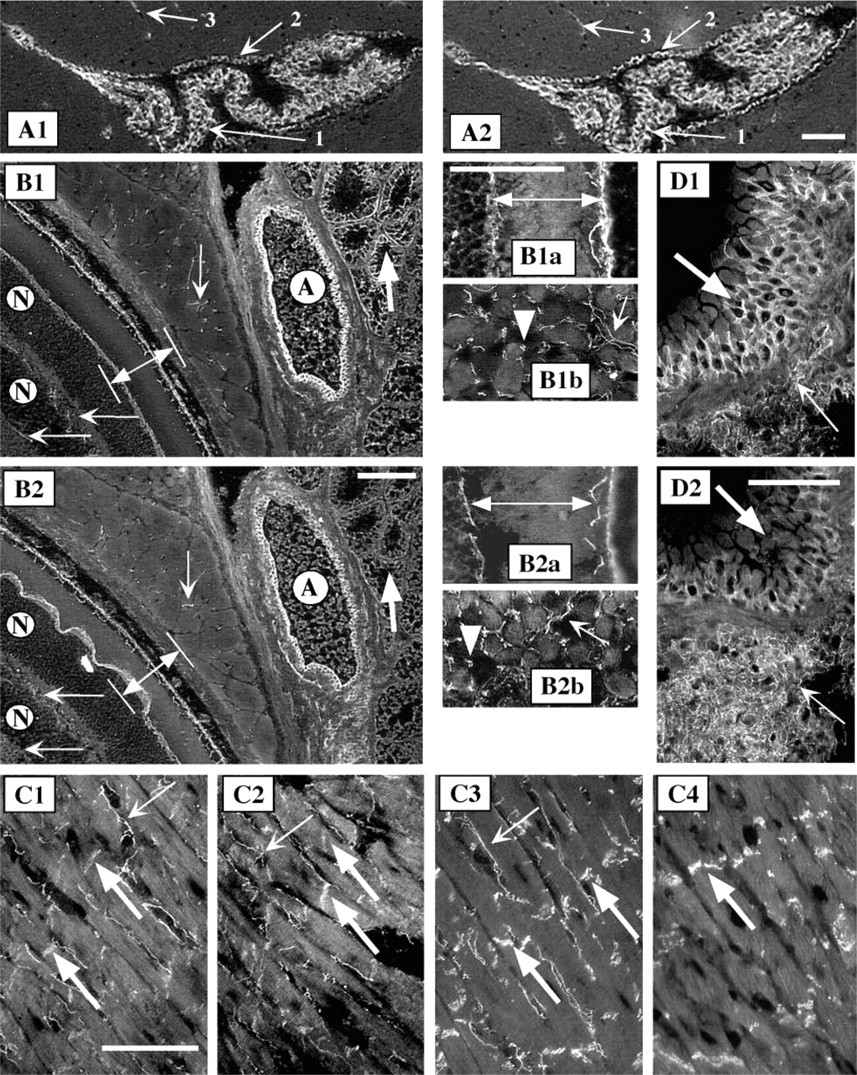
In the adherens junctions, both β-catenin and p120ctn bind to the intracellular domain of cadherins. Because adult mouse epidermis is composed of only two or three layers of epithelial cells, we used neonatal (5-day-old) mouse tissues, including skin, palate, and tongue epithelia, and a duct of a submandibular gland, to study the expression of the components of adherens junctions in stratified epithelia (Figure 3). Both, p120ctn (Figure 3B1–3) and β-catenin (Figure 3C1–3) co-localized with E-cadherin (Figure 3D1–3) when adjacent sections were studied. However, MAb 6H11 detected only occasional weak signal, indicating that the long p120ctn isoform 1 is not a prominent component in the adhesion complexes in epidermis, outer root sheath of hair follicle, the epithelia of palate and tongue, and in the duct of salivary gland (Figure 3A1–3).
However, MAb 6H11 prominently stained distinct structures underlying the epithelial basal cells, especially under the tongue epithelium. These structures, representing microvasculature, were prominent in the tongue muscle (Figure 3A4), where many capillaries run longitudinally through the endomysium and surround each muscle fiber. Similar staining was obtained with MAb pp120, with β-catenin and VE-cadherin antibodies (Figure 3B4-D4), but not with E- or N-cadherin antibodies (data not shown). These results showed that the p120ctn isoform 3 is present at the epithelial cell-cell junctions but that the isoform 1 is present in the endothelial cells in microvascular structures.
The tissue-specific expression of p120ctn isoforms was further studied in adult mouse epithelial tissues. The intercellular junctions of keratinocytes in adult mouse epidermis, contiguous to the hair follicles, stained prominently with pp120 antibody, confirming that the p120ctn isoform 3 is a major adhesion molecule in epidermal keratinocytes (Figure 4A1). The mucosal epithelium of esophagus (Figure 4B1), forestomach (Figure 4C1), and villi of small intestine (Figure 4D1), stained strongly with the pan-p120ctn-specific antibody. The signal with the 6H11 antibody was either absent or hardly detectable in these epithelia with rapid turnover (Figure 4A2-D2). These results are in agreement with the previous findings that p120ctn isoform 3 is abundant in cells with epithelial origin (Staddon et al. 1995; Mo and Reynolds 1996).
However, detailed examination of the mouse fore-stomach revealed distinct staining with MAb 6H11 in the intercellular junctions of serosal epithelium (Figure 4C2 and inset).
The p120ctn Isoform 1 (Cas1) Is Present in Internal Epithelia and Special Cell-Cell Junctions
p120ctn has been demonstrated in choroid plexus and in ventricular ependymal cells of brain, in the pigment cell layer and outer limiting membrane of retina, in capillaries surrounding the muscle fibers, and in the intercalated discs of cardiomyocytes (Staddon et al. 1995; Golenhofen and Drenckhahn 2000).
In this work we used MAb 6H11 for isoform-specific detection of the p120ctn long isoform, isoform 1/Cas1 (Figure 5). In the sections of mouse brain, epithelial cells of the choroid plexus and the ventricular ependymal cells stained strongly, and an occasional capillary in the brain stained with both pp120 and MAb 6H11 (Figures 5A1 and 5A2). While β-catenin was detected in all these structures, N-cadherin could be demonstrated in the ventricular ependymal cell layer but not in the choroid plexus itself (data not shown).
In retina of mouse eye (Figures 5B1 and 5B2) the lateral cell-cell borders of pigment cells and the outer limiting membrane stained prominently with both antibodies (Figures 5B1, 5B2, 5B1a, and 5B2a). The cell-cell junctions in the epithelium of the ciliary body as well as the junctions of the elongated lens cells showed prominent staining with both MAb 6H11 and Mab pp120 (data not shown). The vascular endothelial cells of the arteriole lining the eye (Figures 5B1 and 5B2), as well as the capillaries between the muscle fibers in the transverse section of a muscle lining the eye (Figures 5B1b and 5B2b) gave a distinct staining with both antibodies. However, the cell-cell junctions in the transverse section of the ducts of lacrimal gland stained with MAb pp120, but not with MAb 6H11 (Figures 5B1 and 5B2).
In the cardiac muscle, both pp120 and MAb 6H11 stained the intercalated discs and the endothelial cells of capillaries lining the muscle fibers (Figures 5C1 and 5C2). The immunostaining for β-catenin revealed similar structures (Figure 5C3), while N-cadherin antibody clearly demonstrated the location of intercalated discs between the ends of adjacent cardiac muscle cells (Figure 5C4). Both antibodies also detected the cell-cell junctions in the epicardium, in the layer of endothelial tissue in endocardium, and in the arterioles within the myocardium (data not shown).
In mouse lung tissue, MAb pp120 clearly stained the cell-cell junctions in the epithelium of a bronchiole, whereas the signal obtained with MAb 6H11 was less prominent (Figures 5D1 and 5D2). In the alveolar tissue, where simple squamous epithelium lines the alveolar space and capillaries are abundant, MAb 6H11 signal indicated prominent expression of p120ctn isoform 1.
Discussion
In this work we examined the hypothesis that the p120ctn isoforms are differentially expressed in a tissue- and cell type-specific manner. Our earlier observation on the unique and consistent alternative 5′-splicing of human p120ctn transcripts in each adult and corresponding fetal tissue (Aho et al. 1999) underlined the importance of conducting a study in situ at the protein level. Analysis of a variety of murine cell lines for expression of the p120ctn isoforms, called Cas1 and Cas2, indicated that fibroblasts and macrophages preferentially express isoform 1 (Cas1), and epithelial cells preferentially express isoform 3 (Cas2) (Mo and Reynolds 1996). This led to the conclusion that motile cells preferably express the long isoform 1, where the short isoform is involved in the stable cell-cell adhesion of the epithelial cells. Because of the possibility that the cell type-specific expression of p120ctn isoforms might be affected by the cell culture conditions, we took an in situ approach to distinguish the long isoform from the entire p120ctn population in various mouse tissues.
Stratified squamous epithelium covers body surfaces that are exposed to the external environment. Multiple layers of epithelial cells are held together by the molecular structures called adherens junctions, at which p120ctn binds to the intracellular juxtamembrane region of cadherins, supporting lateral clustering and adhesive strengthening (Yap et al. 1998). Analysis of cells expressing mutated E-cadherin has indicated that p120ctn is required for the E-cadherin-mediated transition from weak to strong adhesion of epithelial cells (Thoreson et al. 2000). Both the long and short isoforms (Cas1 and Cas2) of p120ctn are equally capable of binding to classical cadherins, the interaction requiring an intact Armadillo domain of p120ctn (Reynolds et al. 1996).
In adult mouse skin, the epidermis is composed of only two or three cell layers and it folds into many hair follicles. The neonatal mouse skin is a multilayered tissue, which closely resembles the structure of human epidermis. Our results showed that p120ctn, together with E-cadherin and β-catenin, is a prominent component of cell-cell junctions throughout the epidermis and in the outer root sheath of hair follicles, in the mucosal membranes of palate and tongue, and in the lining of the salivary gland duct. In adult mouse, the stratified epithelium continues to the esophagus and forestomach. In all these structures, the long isoform of p120ctn was hardly detectable, whereas the short isoform was prominent at the cell-cell junctions.
The epithelial sheet of the serous membrane is a thin layer of simple squamous epithelium, lining the body cavities that are not open to the external environment and covering the organs inside these cavities. We found the long p120ctn isoform prominently localized to the lateral cell borders of epithelial cells in the serosa of mouse forestomach, in the epicardium surrounding the heart, in the brain at intercellular junctions of choroid plexus epithelium and ventricular ependymal cells, and in the pigmented epithelial cell layer that separates the retina from the choroid in eye. Remarkable expression of p120ctn long isoform was detected in the cardiovascular system, specifically in the cell-cell junctions of the vascular endothelial cells of endocardium, lining the interior of arterioles, and along the capillaries. In this study we demonstrated that capillaries are abundantly present within various muscles, running in the epimysial space and lining and surrounding the muscle fibers. Capillaries were also detected in other structures, such as the inner layers of retina, and in brain tissue. Both the alveolar tissue and the choroid plexus are rich in capillaries, and we detected prominent staining for the long p120ctn isoform in those structures. The prominent expression of the long isoform in capillaries also explains how the long p120ctn isoform was detected on the Western blot from all the tissues studied. Realizing the fact that almost all the cells in vertebrates are located within 50 μm of a capillary, it is practically impossible to prepare tissue homogenates free of molecular components from capillaries. The long isoform was also demonstrated in two non-epithelial locations, at the intercalated discs joining the adjacent cardiomyocytes and in the outer limiting membrane of retina, which is the line of zonula adherens-like intercellular junctions between Muller cells and the photoreceptor cells.
Our data show that the short isoform of p120ctn is prominently expressed in the stratified squamous epithelium, where cell-cell adhesion is calcium-dependent. The coiled-coil domain at the amino terminus of the long p120ctn isoform can be responsible for specific protein-protein interactions. The function of the p120ctn long isoform, involved in cell-cell adhesion in the internal epithelia composed of a single layer of epithelial or endothelial cells, remains to be studied in the future.
Footnotes
Acknowledgments
Supported by grants from the Dermatology Foundation, and the Maud Kuistila Memorial Foundation, the Finnish Medical Society Duodecim, and the Academy of Finland.
We thank Jari Haikko for tissue cryosectioning and Ms Sue Gotta and Mr Mark Pawlowski for microscopic imaging.