
Abstract
A tyrosine phosphatase-like protein, IA-2, is a major autoantigen in Type 1 diabetes but its role in islet function is unclear. Tyrosine phosphorylation mediates regulation of cellular processes such as exocytosis, cell growth, and cell differentiation. To investigate the potential involvement of IA-2 in islet differentiation and insulin secretion, we analyzed by immunohistochemistry expression of IA-2 during islet development in fetal rats and during the maturation of insulin secretory responses after birth. In the fetus, IA-2 immunoreactivity was detected in primitive islets positive for insulin and glucagon at 12 days' gestation. Subsequently, IA-2 was only weakly detectable in the fetal pancreas. In neonatal rat, a progressive increase in IA-2 immunoreactivity was observed in islets from very low levels at 1 day of age to moderate labeling at 10 days. In the adult, relatively high levels of IA-2 were detected in islets, with heterogeneous expression in individual cells within each islet. IA-2 marks a population of endocrine cells that transiently appear early in pancreatic ontogeny. Islet IA-2 expression reappears after birth concomitant with the development of mature insulin secretory responses, consistent with a role for this protein in regulated hormone secretion.
Phosphorylation of proteins on serine, threonine, or tyrosine residues is a major mechanism for the regulation of cellular processes, including exocytosis, cell growth, and cell differentiation. Members of a novel family of protein tyrosine phosphatase (PTP)-like proteins, IA-2 (also known as ICA512) and phogrin (IA-2β /PTP-NP), were originally identified as islet cell autoantigens in Type 1 diabetes (Lan et al. 1994; Rabin et al. 1994; Payton et al. 1995; Hawkes et al. 1996; Lu et al. 1996). The roles of IA-2 and phogrin in islet function are unknown. Expression of both PTP-like molecules is restricted to islets and other neuroendocrine cell types, including pituitary, adrenal, and some regions of the brain. Both have been localized in these tissues to the secretory granules (Solimena et al. 1996; Wasmeier and Hutton 1996). This subcellular localization and tissue distribution has led to the suggestion that the PTP-like proteins may play a general role in the regulated secretion of granule contents from these neuroendocrine organs. The recent finding that phogrin is phosphorylated in pancreatic islets and β-cell lines exposed to insulin secretagogues supports a role for this family of proteins in the secretory process (Wasmeier and Hutton 1999).
In addition to their potential role in peptide hormone secretion, other observations have implicated the IA-2 family of PTP-like proteins in pancreatic development. In situ hybridization studies have shown that mRNA for phogrin (PTP-NP) is detected very early in the development of the embryonic pancreas and subsequently is expressed in the developing nervous system (Chiang and Flanagan 1996). In the pancreas, phogrin expression was observed in embryonic pancreatic epithelium before the expression of islet hormones, thus identifying the protein as an early and specific marker of islet development. Whether or not the closely related IA-2 is also expressed in the developing pancreas has not been studied. To further investigate the role of the IA-2 family of PTPs in pancreatic islet function, we have analyzed the expression of IA-2 protein during pancreatic islet development in the fetal rat and during the development of regulated insulin secretory responses that occurs after birth (Asplund 1973; Grill et al. 1981).
Materials and Methods
Antibodies
Mouse hybridoma 76F secreting monoclonal antibody to IA-2 was generated after immunization of mice with full-length recombinant human IA-2. An additional five monoclonal antibodies (MAbs) to IA-2 (clones 3C12, 3C11, 4H6, 4C11, 2D8) obtained after immunization of mice with protein representing the cytoplasmic domain of IA-2 were kind gifts from Dr. N. Morgenthaler (Berlin, Germany) and Dr. B. Ziegler (Karlsburg, Germany). A rabbit polyclonal antibody to Pdx-1 (Oster et al. 1998) was a kind gift from Dr. J. Jensen (Gentofte, Denmark), a guinea-pig polyclonal antibody to insulin was from ICN (Thame, UK), and a rabbit polyclonal antibody to glucagon from Sigma (Poole, UK). A mouse MAb to cytokeratin 20 (clone Ks20.8) was from Dako (Cambridge, UK).
Animals and Tissues
We examined pancreas sections for IA-2 expression at the following stages of rat pancreas development: at the time of first appearance of the pancreatic bud at 11 days post conception (dpc) but before the appearance of islet hormones; at the time of the first appearance of primitive endocrine cells at 12 dpc; during the appearance of more mature single hormone positive islets at 13, 14, 15, and 17 dpc; during the rapid expansion of islet cell mass and development of normal insulin secretory responses to secretagogues after birth (at 1, 3, 5, 7, and 10 days of age); and in the adult pancreas.
Wistar rats were mated overnight and the day after mating was considered to be 1 dpc. All animals were kept according to the guidelines of the UK Home Office. Pregnant rats at 11–17 dpc were sacrificed by cervical dislocation and fetuses fixed by immersion in 4% formaldehyde in 0.85% sodium chloride solution (formal saline) for 18 hr before dehydration and embedding for light microscopy. In some experiments the fetal pancreas was removed by dissection under the microscope and fixed for 4 hr in formal saline or extracted for Western blotting as described below. Pancreata from rats of 1–10 days of age or from adult rats were dissected and fixed for 4 hr in formal saline or processed for islet isolation as previously described (Brundstedt et al. 1984). Pituitary tissue was obtained from adult male Wistar rats and transformed β-cells of the βTC3 cell line were harvested by scraping from tissue culture flasks after maintenance in tissue culture in Dulbecco's modified Eagle's medium (DMEM) containing 25 mM glucose and 10% fetal calf serum.
Immunohistochemistry
Sensitive detection of IA-2 was achieved by immunoperoxidase labeling with signal amplification using streptavidinbiotin complexes. Paraffin sections 5–6 μm thick of tissues or rat embryos were dewaxed and rehydrated for immunohistochemistry. Endogenous peroxidase was blocked by treatment with 0.3% hydrogen peroxide in methanol and sections treated with normal swine serum (20% in Tris-buffered saline) to prevent nonspecific labeling. Sections were analyzed for reactivity with antibodies to IA-2, using at least two different MAbs at each stage of pancreatic development. Consecutive sections were labeled with antibodies to Pdx-1, glucagon, insulin, and cytokeratin-20 as markers for endocrine and ductal pancreatic cells. For detection of Pdx-1, sections were cut onto Vectorbond-coated slides (Vector Labs; Peterborough, UK) and heated in a pressure cooker at full pressure for 2 min in 0.01 M sodium citrate, pH 6.0, before incubation with specific antibody. For the detection of cytokeratin-20, sections were treated with trypsin (0.1% in 0.125% CaCl2 for 4 min at 37C) before antibody labeling. Sections were incubated overnight at 4C with monoclonal or polyclonal antibodies as described in the text or figure legends. Antibody reaction was visualized by the peroxidase-labeled streptavidin method with 3,3′ diaminobenzidine substrate using a commercial kit (LSAB Plus; Dako, Cambridge, UK). Sections were counterstained with hematoxylin, dehydrated, and mounted for microscopic inspection. In dual-labeling experiments to examine potential co-localization of IA-2 with endocrine cell markers, sections were first labeled with IA-2 antibodies and detected with with 3,3′ diaminobenzidine substrate. Sections were then incubated with the second primary antibody (to insulin or glucagon) for 1 hr at room temperature, washed, and antibody binding detected with FITC-conjugated anti-guinea pig or anti-rabbit immunoglobulin (Stratech; Luton, UK).
Western Blotting
Tissues or cultured cells were gently homogenized in lysis buffer (50 mM Tris, pH 7.5, 10 mM sodium phosphate, 150 mM NaCl, 1% Triton X-100, 5 mM EDTA, 10 mM NaF, 5 mM iodoacetate, 1 mM benzamidine, 5 μg/ml leupeptin, 5 μg/ml aprotinin) and incubated for 30 min on ice. The lysate was centrifuged at 10,000 × g for 20 min to remove insoluble material and the protein concentration determined. Aliquots of tissue lysates representing 10 μg of protein or 20 ng of recombinant protein representing the cytoplasmic domains of IA-2 or phogrin, purified as previously described (Payton et al. 1995; Hawkes et al. 1996), were separated by SDS-PAGE on 10% polyacrylamide gels and transferred to nitrocellulose membranes. Membranes were blocked in Tris-buffered saline (TBS) containing 0.05% Tween-20 and 2% skimmed milk powder, then incubated with tissue culture supernatants containing MAbs to IA-2 at 1:10 dilution. After washing in TBS–0.05% Tween-20, antibody binding was visualized with a horseradish peroxidase-conjugated antibody to mouse IgG (Sigma) using a chemiluminescent substrate (ECL; Amersham Pharmacia Biotech, Amersham, UK).
Results
Specificity of Antibodies for IA-2
Six mouse MAbs recognizing the cytoplasmic domain of IA-2 were available for analysis of IA-2 expression. The antibodies were initially tested for their specificity by Western blotting against extracts of rat pituitary and βTC cells, and against purified recombinant proteins representing the cytoplasmic domains of IA-2 and phogrin. All MAbs gave a similar pattern of reactivity against proteins in pituitary and β-TC cell extracts, with a predominant doublet or multiple bands at Mr 60,000–65,000 (Figure 1). All antibodies reacted strongly to the recombinant IA-2 preparation and only antibodies from clones 4H6 and 2D8 (weakly) crossreacted with the closely related protein phogrin. Antibody reactivity was confirmed by immunoprecipitation of recombinant protein (data not shown). Antibodies showing specificity for IA-2 (clones 3C12, 3C11, 4C11, and 76F) were selected for further studies of IA-2 expression by immunohistochemistry.
Expression of IA-2 During Fetal Pancreas Development
Consecutive sections of formalin-fixed rat fetuses of 11–17 dpc were labeled with antibodies to Pdx-1, cytokeratin 20, glucagon, and insulin to identify the developing pancreas and primitive endocrine cells, and with two or more MAbs to IA-2. No IA-2 labeling was detected in the pancreatic primordia appearing as Pdx-1-positive structures at 11 dpc before the appearance of endocrine cells (Figures 2A and 2B). However, weak IA-2 labeling was observed in primitive glucagon-positive cells appearing early at 12 dpc (Figures 2C and 2D). These primitive endocrine structures were negative for Pdx-1, whereas surrounding pancreatic epithelium forming the major part of the pancreatic bud was positive (Figure 2E). Larger islet-like structures appearing at 12 dpc, positive for both glucagon and insulin, showed strong positivity for IA-2 immunoreactivity (Figures 2F-2H). IA-2 expression was confirmed with two different MAbs to the protein, whereas controls performed by omitting the primary antibody step showed no labeling on any of the sections. In contrast to the observations at 12 dpc, examination of sections from fetal rats at 14 dpc onwards revealed very little IA-2 labeling in the fetal islets, with only sporadic cells positive (Figures 2I and 2K). Single-hormone positive cells, predominantly glucagon rather than insulin, were visible at this stage (Figures 2J and 2L). No labeling with IA-2 antibodies was detected outside of the pancreas, including neuronal tissue, at any gestational age in the sections analyzed. Therefore, throughout fetal development, IA-2 expression appears restricted to the primitive Pdx-1-negative islet-like structures appearing at 12 dpc.
Expression of IA-2 After Birth
Islet development continues after birth and, in the rat, glucose-induced insulin secretion is not apparent until a few days after birth (Asplund 1973; Grill et al. 1981). To evaluate the expression of IA-2 during this period, formalin-fixed sections of dissected pancreas from 1–10-day-old and adult rats were labeled for IA-2 expression using the selected MAbs. Very weak IA-2 labeling was detected in islets of the 1-day-old rat (Figure 3A). On sections from older rats, a progressive increase in the intensity of IA-2 labeling was observed with increasing age, with moderate IA-2 expression detectable in the 3–10-day-old rat (Figures 3B and 3C) and strong labeling in the adult (Figure 3D).
In the neonatal rat, some endocrine cells, predominantly located at the periphery of islets, co-express the epithelial marker cytokeratin-20, suggestive of cells that are transitional in the differentiation from duct cells to islets (Bouwens et al. 1994). In our study, we confirmed the presence of cytokeratin-20 in both ducts and peripheral islet cells in the pancreas of 1- and 3-day-old rats (Figure 3E). IA-2 was largely absent from ducts and peripheral islet cells in the 1- and 3-day-old rat but was weakly expressed in cells at the core of the islet (Figure 3F). The majority of the IA-2-positive cells expressed Pdx-1 (Figures 3G and 3H), which by this stage of development is restricted to endocrine cells (Guz et al. 1995).
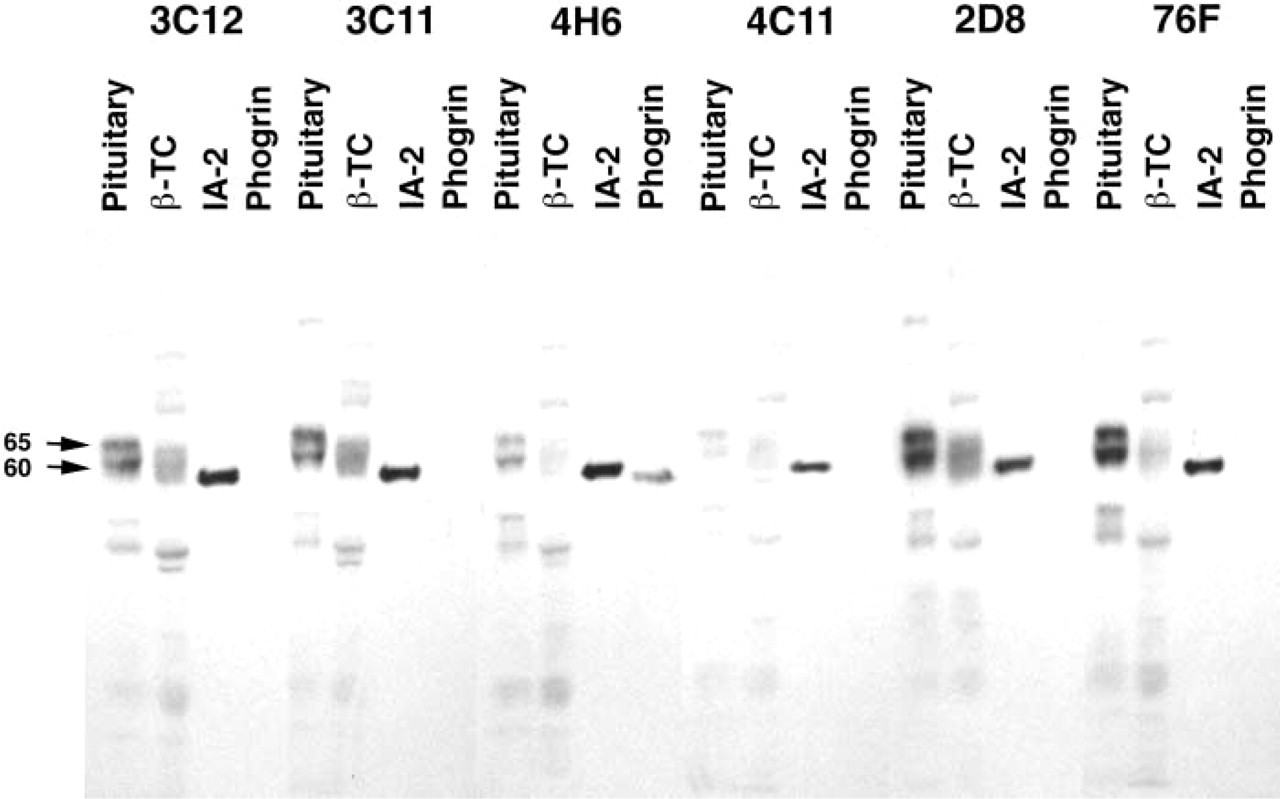
Specificity of IA-2 MAbs by Western blotting. MAbs were tested for their ability to recognize proteins in extracts of rat pituitary and βTC cells. Reactivity with purified recombinant fusion protein representing the cytoplasmic domain of IA-2 or phogrin was also analyzed. The locations of the dominant 60,000–65,000 Mr proteins in pituitary and βTC extracts are marked (left arrows).
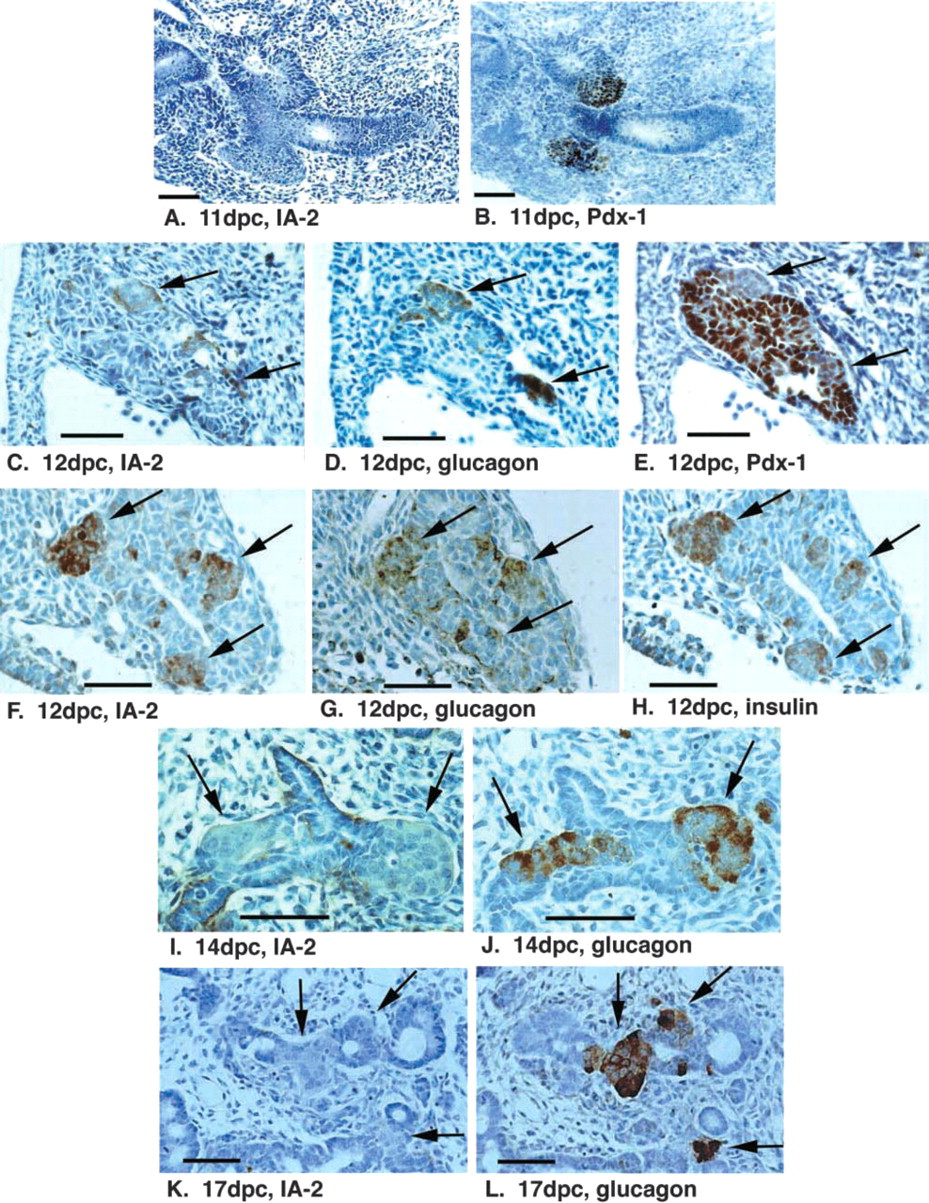
Expression of IA-2 in early pancreas development. Sections of rat fetal pancreas at 11 dpc (
Within each islet there was considerable heterogeneity among cells in the intensity of IA-2 labeling at all ages, and this was particularly evident in the adult pancreas (Figure 3J). Although less evident than for IA-2, there was also heterogeneity in insulin labeling within individual islets, with strongest labeling occurring in β-cells at the periphery of the islet (Figures 3I and 3K). Double labeling of sections to co-localize IA-2 (by peroxidase labeling) and insulin (by immunofluorescence) in β-cells revealed that cells expressing highest levels of IA-2 did not necessarily have high insulin content (compare Figures 3I and 3J). In case of potential artifacts caused by quenching of immunofluorescence by the DAB precipitate, the discordance between high IA-2 and insulin expression was confirmed by labeling consecutive mirror sections with antibodies to insulin and IA-2 (Figures 3K and 3L). IA-2 was present predominantly in β-cells (e.g., the cell labeled by large arrow in Figures 3K and L) but also non-β-cells (small arrow) of the endocrine pancreas.
Western Blotting Analysis of IA-2 in Tissues
To confirm the presence of IA-2 in the fetal and neonatal pancreas, Western blotting studies were performed with tissue extracts (10 μg total extracted protein) prepared from dissected pancreata of 12 and 14 dpc rat fetuses, isolated islets from 5- and 10-day-old newborn rats, and adult rat islets. Predominant reactivities of the MAbs to IA-2 were to multiple bands of Mr 60,000–65,000 (Figure 4). These multiple bands were visible in the 12 dpc fetal rat pancreas but were weak in the 14 dpc pancreas. Increased levels of staining of these bands were observed in isolated islets from 5 days of age to adult. These results, taken together with the histochemical observations, are consistent with a transient expression of IA-2 in the 12 dpc fetal pancreas and increased islet expression of the protein after birth.
Discussion
In this study, immunohistochemical techniques were used to visualize IA-2 expression during rat pancreas development using selected MAbs raised to the cytoplasmic domain of the molecule. Analysis of reactivity to deletion mutants of IA-2 indicated that these antibodies recognize different epitopes within either the PTP-like domain or the juxta-membrane domain of the molecule (data not shown). We selected for our study antibodies that did not crossreact with recombinant phogrin on Western blots. The MAbs showed similar patterns of binding to proteins in extracts of adult rat pituitary and β-TC cells, with predominant reactivity towards multiple bands of Mr 60,000–65,000 and minor bands of Mr 110,000, 130,000, and 40,000. Similar multiple forms of IA-2 have been reported in Western blotting studies of bovine pituitary (Hermel et al. 1999). Partial characterization of these IA-2 species suggest that the higher molecular weight variants represent precursor forms that are differentially glycosylated and that are subsequently subject to proteolytic processing to the 60,000–65,000 Mr forms (Hermel et al. 1999). Studies on phogrin have identified 65,000 and 60,000 Mr bands on SDS-PAGE as phosphorylated and dephosphorylated forms of the protein (Wasmeier and Hutton 1999). The multiple bands around 65,000 Mr may represent differentially phosphorylated or other post-translationally modified forms of IA-2.
Pancreatic islets develop from progenitor cells localized within duct-like structures formed by proliferation of epithelial cells in the pancreatic bud (Pictet and Rutter 1976). In early fetal life, the pancreatic epithelium transiently expresses the insulin gene transcription factor Pdx-1 (Guz et al. 1995). Primitive islets are first detectable by hormone labeling (glucagon, insulin) as isolated cells or groups of cells close to duct-like structures at around 7 weeks' gestation in humans and 11 dpc in the rat. These primitive islets have low levels of Pdx-1 (Oster et al. 1998). Later in fetal development, islets express high levels of Pdx-1 and the transcription factor is eventually restricted to islets late in fetal life and after birth (Guz et al. 1995).
Expansion of islet cell mass continues throughout fetal and early neonatal life and is believed to occur both as a result of growth of pre-formed islets and by neo-islet formation from ductal epithelium (Bouwens et al. 1994). During fetal life, IA-2 was transiently expressed in primitive islets at 12 dpc, at the time of first appearance of glucagon- and insulin-positive cells. Analysis of consecutive sections of fetal pancreas suggested that this population of cells co-expressed insulin and glucagon but was negative for the homeobox protein Pdx-1, consistent with a previous study of hormone-expressing cells in the pancreas at this stage of development (Oster et al. 1998). IA-2 expression in the pancreas at 12 dpc was confirmed by Western blotting, whereby IA-2 was detected as multiple bands of 60,000–65,000 Mr, similar to those detected in the neonatal and adult islet. IA-2 was not detected by immunocytochemistry at 11 dpc, before the appearance of glucagon and insulin, and was only weakly visible or undetectable from 14 dpc onwards, when single hormone positive cells are apparent. The role of the early IA-2-positive cell population is unclear. Multiple hormone-expressing cells appearing early in fetal development may represent precursors of the single hormone-positive cells that appear later (Teitelman et al. 1993). Alternatively, these may represent distinct cell lineages, with the majority of mature islets being derived from hormone-negative precursors in pancreatic epithelium (Pang et al. 1994; Karaka–Gantenbein et al. 1995; Oster et al. 1998). The multiple hormone-expressing cells do store insulin and glucagon in secretory vesicles (Teitelman et al. 1993). It remains to be established whether IA-2 expressed by these cells is localized within these vesicles and whether regulated secretion of hormones or other products from the cells contributes to the further growth and differentiation of the fetal endocrine pancreas.
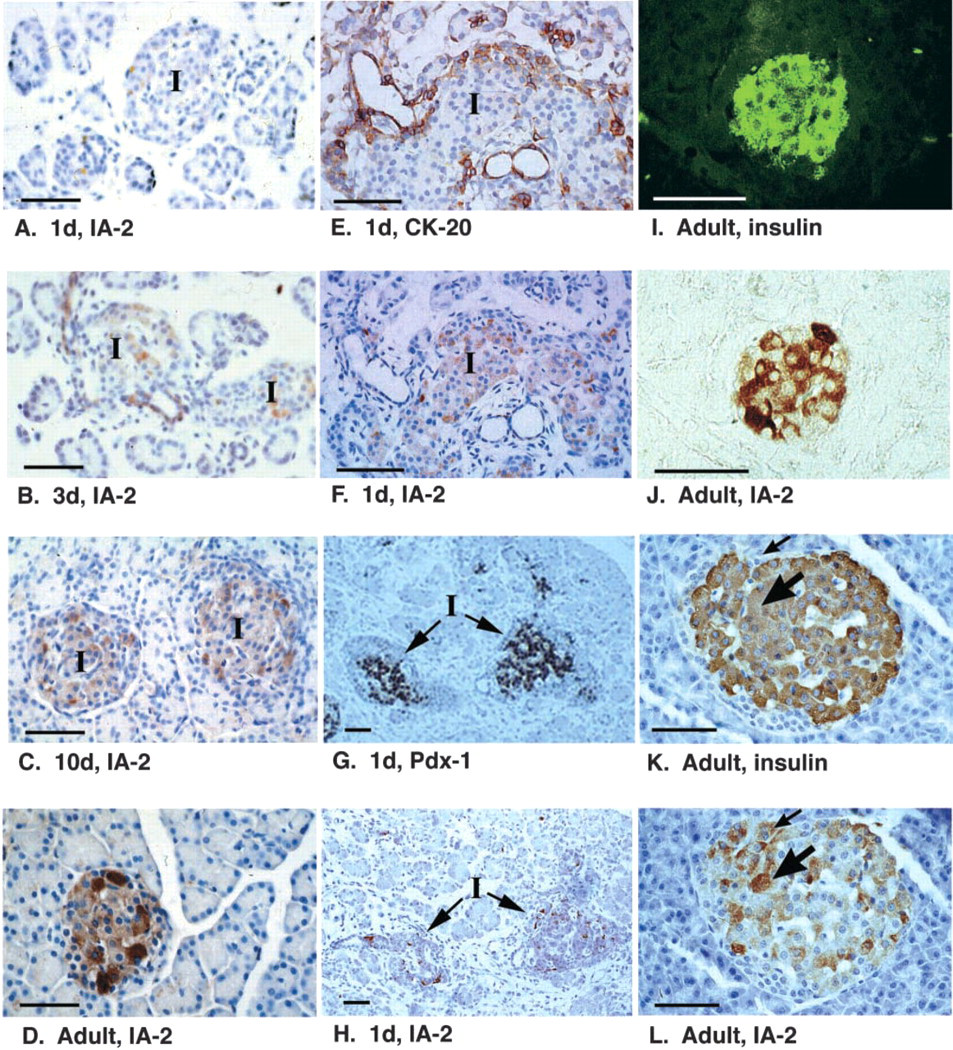
Expression of IA-2 after birth. Sections of pancreas from rats of progressively older age (
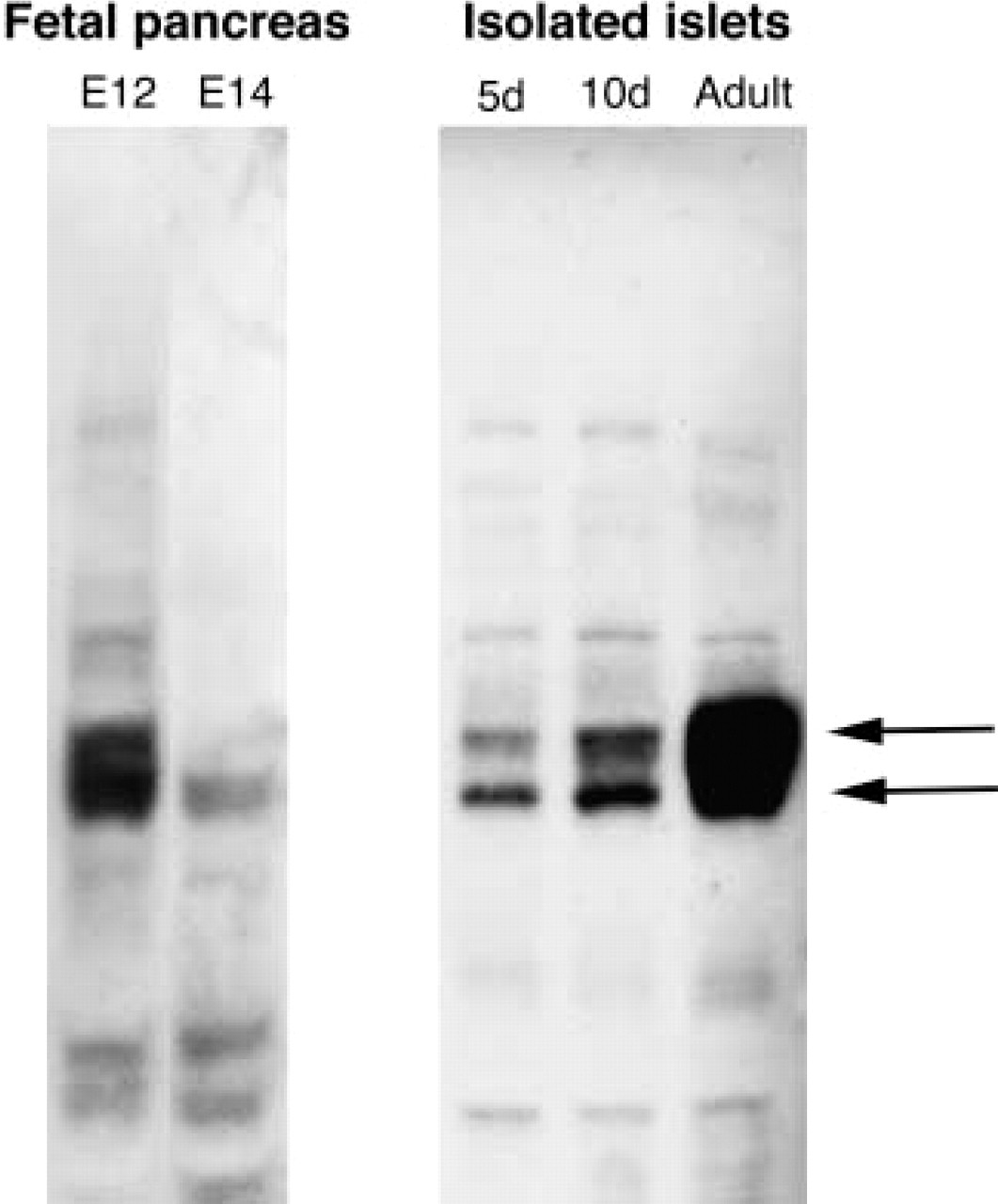
Western blotting analysis of IA-2 expression in fetal and neonatal pancreas. Extracts (10 μg protein) of 12 dpc or 14 dpc fetal pancreas, or isolated islets from 5-day-old, 10-day-old, or adult rats were used in Western blotting experiments with monoclonal IA-2 antibody clone 76F. Arrows (right) show the position of the major 60,000–65,000 Mr proteins on the blot.
The transient appearance of IA-2 in a subpopulation of pancreatic endocrine cells contrasts with a different pattern of expression of the structurally related phogrin (PTP-NP) reported by Chiang and Flanagan (1996) in the developing mouse. The authors used in situ hybridization to show expression of phogrin throughout the developing central nervous system and in the midgut endoderm and embryonic pancreatic epithelium before the expression of islet hormones. Phogrin was subsequently expressed also in insulin- and glucagon-positive cells early in development (9.5 dpc), and islet phogrin expression (in all pancreatic endocrine cells) persisted throughout fetal life. Although confirmation of expression is required at the protein level, phogrin may show an earlier and more persistent pattern of expression during fetal life than IA-2. Nevertheless, our finding that IA-2 is expressed in a specific population of endocrine cells appearing very early in pancreatic development offers support for a role for IA-2 in the developmental process.
In fetal life, IA-2 expression was only transient; from 14 dpc, IA-2 expression in islets was very weak or absent. However, IA-2 labeling was again detectable 1 day after birth, and a progressive increase in islet IA-2 expression was observed during the first few days of life. In the neonate, IA-2 expression was restricted to mature endocrine cells. Cytokeratin-20-positive cells at the periphery of islets, which may represent immature endocrine cells at a transitional stage of differentiation from ductal precursors to islets (Bouwens et al. 1994), were negative. In the adult pancreas, strong but heterogeneous expression of IA-2 was observed in both β- and non-β-cells of the islet. The progressive increase in IA-2 expression after birth occurs in parallel with the maturation of the insulin secretory response to secretagogues. It is well established that isolated islets from fetal and early neonatal rats show relatively poor insulin secretory responses to glucose and other nutrient secretagogues compared to islets from adult animals (Asplund 1973; Grill et al. 1981). The poor response does not appear to be a consequence of low insulin reserves, because islets appear fully granulated by this stage of development and the insulin content of islets from neonatal rats is similar to that from adults (Grill et al. 1981). If IA-2 represents a critical element of regulated secretory pathways in neuroendocrine cells, then the low levels of expression of the protein in pancreatic islets during fetal and neonatal life may contribute to the generalized diminution of insulin secretory responses to secretagogues during this period. Upregulation of IA-2 expression may be necessary for maturation of the secretory response.
Within individual islets in the neonatal and adult pancreas, there was considerable heterogeneity among cells in levels of expression of IA-2. In vitro observations have established that individual β-cells show heterogeneity in a number of characteristics, including insulin biosynthesis, insulin release, activity of ATP-sensitive K+ channels, glucose utilization and oxidation, and morphological appearance of insulin secretory granules by electron microscopy (Misler et al. 1986; Kiekens et al. 1992; Pipeleers 1992). In situ, intercellular differences in hormone distribution have been observed (Stefan et al. 1987), as well as differences in the relative proportions of insulin and proinsulin (Orci 1985). In this study, dual labeling with antibodies to IA-2 and insulin, or comparison of adjacent mirror sections labeled with these antibodies, did not reveal common patterns of high and low expression within individual islets. The diversity of IA-2 expression within individual islets may not simply reflect differences in insulin content and granulation. IA-2 expression thus represents an additional marker for islet cell heterogeneity independent of hormone content.
IA-2 is a major target for autoimmunity in human Type 1 diabetes. The finding that IA-2 is expressed only at low levels throughout most of fetal and early neonatal life, whereas phogrin expression may be more persistent, may be relevant to the initiation of autoimmune responses to these proteins in Type 1 diabetes. It has long been recognized that neonatal mice are more susceptible to tolerance induction than adult mice (Billingham et al. 1953), and the fetal and neonatal period has been suggested to be important for the induction of immunological self-tolerance (Burnet 1959). Studies on transgenic mice expressing SV40 large T-antigen in pancreatic β-cells demonstrated that mice develop autoimmune responses to the protein, as well as lymphocytic infiltration into pancreatic islets, when transgene expression is delayed until post-neonatal life (Adams et al. 1987). Immunity to T-antigen was not observed in mice expressing the transgene in the neonatal period. The delay in expression of IA-2 in pancreatic islets until after the neonatal period may therefore result in ineffective induction of tolerance to the protein. These observations may, at least in part, explain why IA-2, rather than phogrin, is the dominant PTP-like autoantigen in diabetes (Bonifacio et al. 1998), despite similar structure and subcellular localization of the two proteins in the adult.
Footnotes
Acknowledgements
Supported by grants from the British Diabetic Association (RD96/0001250) and the European Union (TMR CT 970142). GR was supported by a Medical Research Council Clinical Training Fellowship.
We thank Dr Nils Morgenthaler, Dr Brigitte Ziegler, and Dr Jan Jensen for gifts of antibodies.