
Abstract
The cellular localization of protein tyrosine phosphatase 51 (PTPIP51) and its in vitro interacting partner protein tyrosine phosphatase 1B (PTP1B) was studied in human placentae of different gestational stages. The expression of PTPIP51 protein and mRNA was observed in the syncytiotrophoblast and cytotrophoblast layer of placentae from the first, second, and third trimesters. In contrast, PTP1B expression was restricted to the syncytiotrophoblast during all gestational stages. Cells of the cytotrophoblasts and parts of the syncytiotrophoblasts expressing high amounts of PTPIP51 were found to execute apoptosis as shown by TdT-mediated dUTP-biotin nick end labeling assay, cytokeratin 18f, and caspase 3 expression. PTPIP51 could also be traced in the endothelium and smooth muscle cells of placental arterial and venous vessels, identified by double immunostainings with antibodies directed against van Willebrand factor and α-smooth muscle actin. Some of these cells showing a high PTPIP51 reactivity were Ki67 positive, indicating proliferation. Additionally, a small population of placental CD14-positive macrophages and mesenchymal cells within the villous stroma were detected as PTPIP51 positive. Our data suggest that both proteins, PTPIP51 and PTP1B, play a role in differentiation and apoptosis of the cytotrophoblast and syncytiotrophoblast, respectively. Moreover, PTPIP51 may also serve as a cellular signaling partner in angiogenesis and vascular remodeling.
Keywords
P
The human Ptpip51 gene is located on chromosome 15 (15q15.1) and encodes the corresponding full-length protein making up 470 amino acids by 12 exons. PTPIP51 is evolutionary conserved and was shown to be expressed in different mammalian tissues and organs (Stenzinger et al. 2005), cancer (Lv et al. 2006; Koch et al. in press), as well as different cell lines including BeWo cells (human choriocarcinoma cell line) (Stenzinger et al. 2005). A regulated layer-dependent expression pattern was particularly observed for epidermal and seminiferous epithelia, both requiring a carefully controlled balance between proliferation, differentiation, and apoptosis (Nickoloff and Denning 2001; Stenzinger et al. 2005, 2006). Moreover, a study conducted by Märker et al. (2008) pointed to a crucial role for PTPIP51 during mouse eye development. In accordance with these data, Roger et al. (2007) described a stage-dependent, biphasic expression pattern of Ptpip51 in postnatal retinal development, which is governed by ciliary neurotrophic factor (CNTF), a member of the interleukin-6 family that mediates its action by the Janus-kinase/signal transducers and activators of transcription (JAK-STAT) signaling pathways. The protein was also found to be expressed during the embryonic development of other organs (Märker D, unpublished data). Interestingly, Lv et al. (2006) found the full-length form of PTPIP51 to harbor a mitochondrial target sequence, thereby enhancing apoptosis in vitro. Our own results on vitamin- and cytokine-regulated expression of PTPIP51 in a human keratinocyte cell line supported these results, and moreover, pointed to a role of PTPIP51 in the differentiation of cells (Stenzinger et al. 2006).
As in the epidermis, the formation and functional integrity of the syncytiotrophoblast, evolving from the cytotrophoblast, essentially requires a finely tuned interplay between proliferation, differentiation, and apoptosis (Huppertz et al. 2006; Kar et al. 2007). The regulation of these processes is dependent on numerous signaling cascades, including tyrosine phosphorylation. Disturbances of this delicate balance may lead to gestational trophoblastic diseases, such as moles and choriocarcinomas (Li et al. 2002; Shih 2007).
For these reasons, we decided to study the histological localization of the novel protein PTPIP51 and its interacting partner PTP1B in human placenta during distinct weeks of gestation.
Development of the placenta begins after implantation of the blastocyst with the decidual reaction and transformation of maternal endometrium. The placenta develops from the single cell layer (trophoblast) of the blastocyst, whereas the embryo evolves from the three to four cell layer region (embryoblast). The trophoblast undergoes rapid proliferation and differentiation into an inner cytotrophoblast and an outer syncytiotrophoblast. Undifferentiated cytotrophoblast cells invade maternal tissue and blood vessels of the decidua and myometrium. By terminal differentiation of mononucleated cytotrophoblast cells, the multinucleated syncytiotrophoblast arises. The syncytiotrophoblast forms an external layer without intercellular boundaries. Its cells form cords infiltrating the endometrium. Through the lytic activity of the syncytiotrophoblast, the maternal capillaries are eroded and anastomose with the trophoblast lacunae, which with progressing pregnancy form a single, connected system. Primary villi are formed by penetration of cytotrophoblast cells into the syncytiotrophoblast, which are invaded by the mesoblast. At the end of the third week, the villous mesoblast differentiates into connective tissue and blood vessels, which get connected to the embryonic blood vessels. The cytotrophoblast of the anchoring villi grows rapidly, forming the cytotrophoblast layers between the syncytiotrophoblast and the uterine endometrium. After the fourth month, the cytotrophoblast slowly disappears from the walls of the placental villi, and the syncytiotrophoblast forms the only continuous layer separating the maternal blood and the fetal capillary endothelium.
This study aims to further elucidate in vivo functional implications of PTPIP51 by double immunostainings detecting apoptotic and proliferative cells, which are known to be essential for placental development.
Materials and Methods
Tissue and Section Preparations
This study was performed with human placenta samples from different weeks of gestation, all obtained from the Department of Obstetrics and Gynecology, Justus-Liebig-University, Giessen, Germany. Normal first trimester placenta samples (n = 4; gestational weeks 9, 9, 10, and 11) were obtained from cases of elective termination of pregnancy for psychosocial reasons. Second trimester placenta specimens (n = 3; gestational weeks 20, 23, and 27) were obtained from stillbirths of the fetus. Third trimester placental specimens (n = 7; gestational weeks 36–40) were taken from women with normal pregnancies who delivered by primary caesarean section because of breech presentation. No placenta had histopathological abnormalities. Unfixed samples were cut out immediately and subsequently snap frozen in melting isopentane. The study was approved by the Ethics Committee, Medical Faculty, Justus-Liebig-University, Giessen, Germany.
PTPIP51 Antibody Production
The cDNA sequence encoding aa 131–470 of PTPIP51 were inserted into the BamHI and HindIII sites of the plasmid pQE30 and expressed as His6-tagged protein in the protease deficient E. coli expression strain AD202 [araD139DE(argF-lac)169 ompT1000:kan flhD5301 fruA25 relA1 rps150(strR) rbsR22 deoC1]. The protein was purified to electrophoretic homogeneity by chromatography on an Ni-agarose column (Porsche 2001). Immunization of rabbits was performed with 0.5 mg of the purified protein in 0.5 ml RIBI adjuvant followed by booster injections with 0.5 and 0.3 mg on days 14 and 21, respectively. The antiserum was collected on day 28. Monospecific antibodies were prepared following the method described by Olmsted (1981). Briefly, 2 mg of purified antigen was blotted on nitrocellulose after SDS electrophoresis. The protein band was marked with Ponceau solution and cut out. After blocking of the membrane strip with 1% low-fat milk powder in PBS, the membrane was incubated with the antiserum for 1 hr followed by extensive washing with Tris-EDTA-buffered saline. The antibodies were eluted with 0.2 M glycine (pH 2.0) for 2 min, followed by immediate neutralization with 1 M triethanolamine.
List of the antibodies used in this study
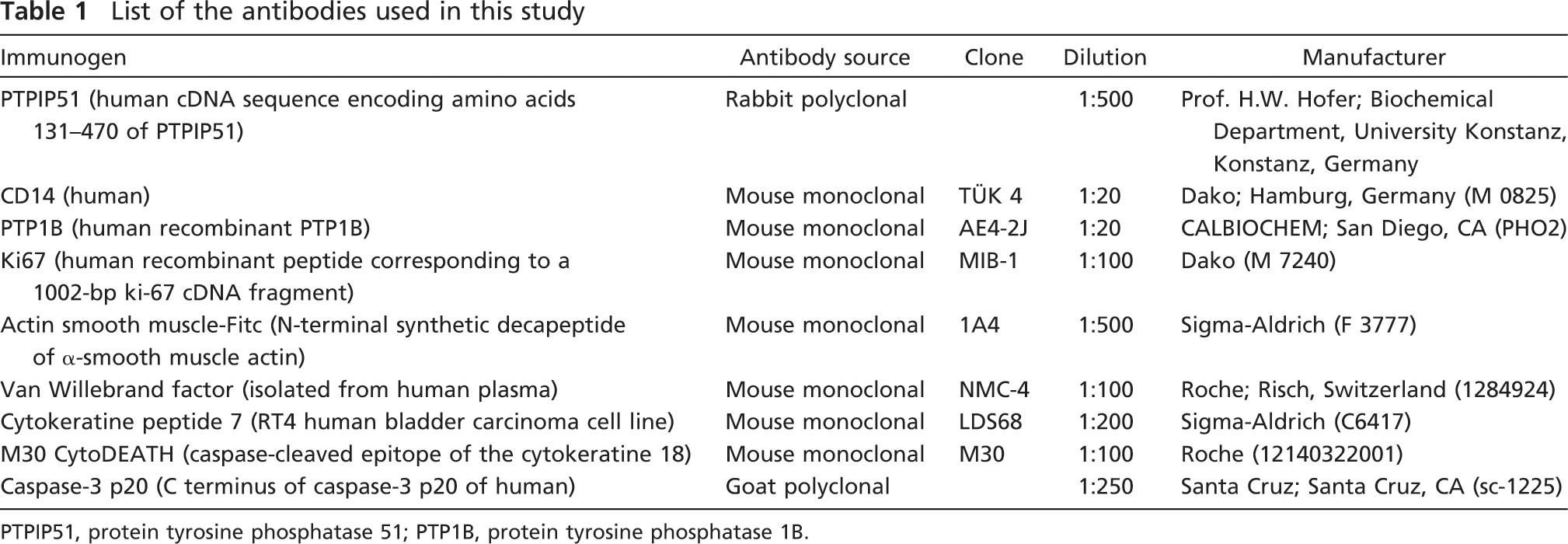
PTPIP51, protein tyrosine phosphatase 51; PTP1B, protein tyrosine phosphatase 1B.
IHC
IHC stainings of 7-μm-thick serial cryosections or Hepes glutamic acid buffer mediated organic solvent protection effect-fixed paraffin sections were performed according to a standard protocol. The sections were fixed for 10 min at −20C in acetone before immunostaining.
Primary antibodies used in this study are listed in Table 1.
They were visualized through Alexa Fluor 488 and Alexa Fluor 555 secondary antibodies (Molecular Probes; Eugene, OR). For each series of antibody staining, sections were incubated with medium lacking PTPIP51 antibody, which served as an internal negative control. Nuclei were shown using 4′-6-diamidino-2-phenylindole.
Apoptosis was detected by an in situ cell death detection kit ApoTag (S7110; Chemicon International, Temecula, CA). The ApopTag Fluorescein In Situ Apoptosis Detection Kit detects apoptotic cells in situ by the indirect TdT-mediated dUTP-biotin nick end labeling (TUNEL) method, using an anti-digoxigenin antibody that is conjugated to a fluorescein reporter molecule. It provides indirect immunofluorescence staining. Results were analyzed by using fluorescence microscopy. It was used according to the manufacturer's instructions.
The Axioplan 2 fluorescence microscope equipped with Plan-Apochromat objectives (Carl Zeiss; Jena, Germany) was used for photo documentation. For visualization of the secondary antibody, Alexa Fluor 555, an excitation filter with a spectrum of 530–560 nm and an emission filter with a spectrum 572.5–647 nm were used. Alexa Fluor 488 was visualized by an excitation filter with a range of 460–500 nm and an emission filter with a range of 512–542 nm.
Hematoxylin and Eosin Counterstaining
After obtaining photographs from immunostained samples, the coverslip of each slide was removed, and the sections were subsequently stained with hematoxylin and eosin (H&E). Light microscopy was used to take H&E-stained pictures of the corresponding IHC section.
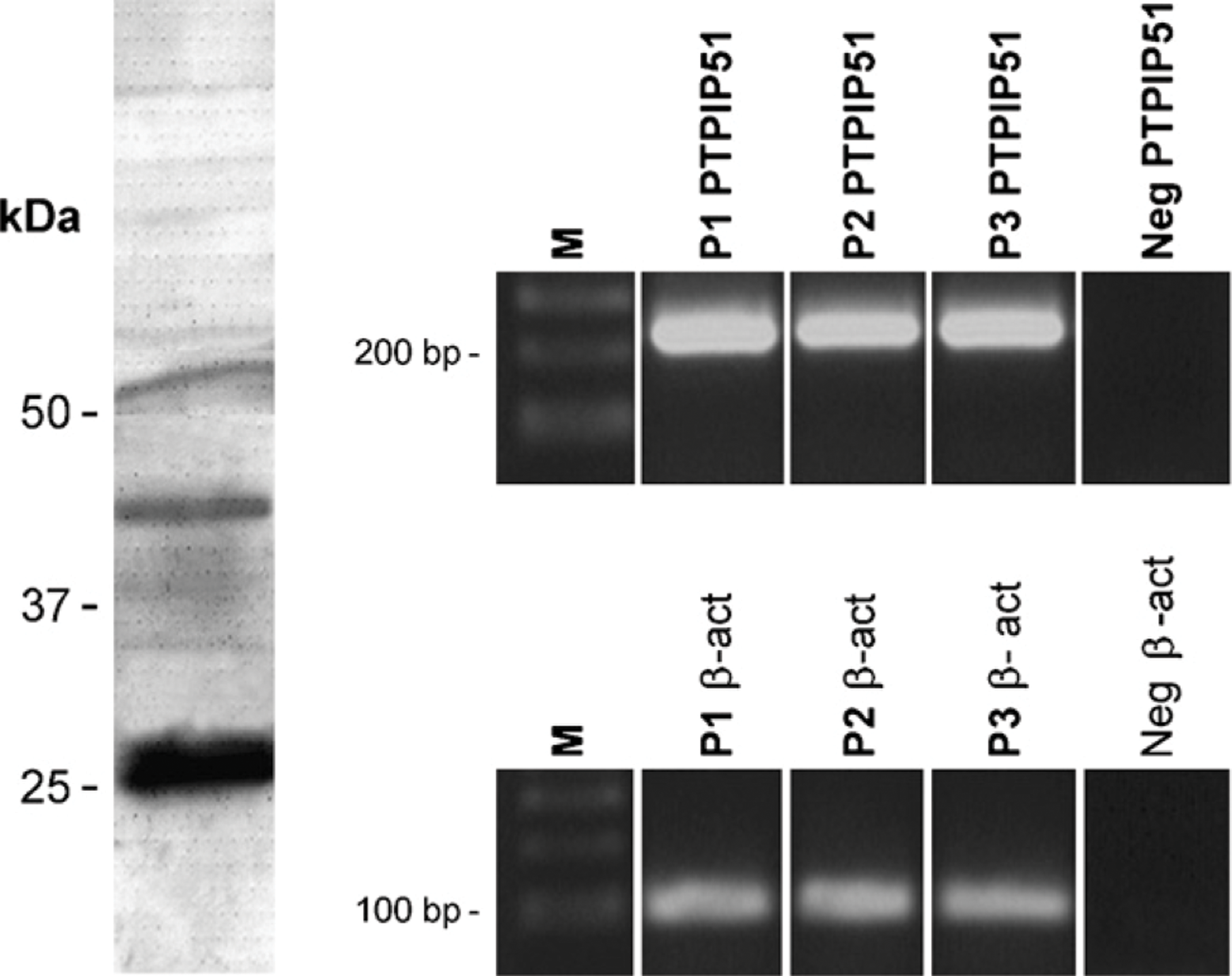
Immunoblotting and RT-PCR of protein tyrosine phosphatase 51 (PTPIP51) in placenta: (Left) Immunoblotting of term placentae probed for PTPIP51 antigen. (Right) RT-PCR was performed using primers specific to PTPIP51 as given in Material and Methods. β-Actin was amplified as an internal positive control, and probes lacking RT served as negative controls (Neg). Marker (M), samples of placenta 1 (P1), placenta 2 (P2), and placenta 3 (P3). The white bar between the marker and the lanes indicates that the lanes were depicted from different gels.
ISH
Production of DIG-labeled cRNA probes was performed as described previously (Steger et al. 1998). PCR conditions were 1 × 94C for 3 min, 35 × 94C for 30 sec, 66C for 30 sec, 72C for 60 sec, and 1 × 72C for 8minusing5′-GTCTCTCACCTGAGCTGGCTAGA-3′ as the forward primer and 5′-GGATAGCCAAATCCTCCTTCGTGA-3′ as the reverse primer. The 247 nucleotide (nt) endoplasmic reticulum product (nt position 1126–1372, corresponding to aa 376–457) of the human Ptpip51 gene was subcloned in pGEM-T (Promega; Heidelberg, Germany). Plasmids were transformed in the XL1-Blue E. coli strain (Stratagene; Heidelberg, Germany) and extracted by column purification (Qiagen; Hilden, Germany). In vitro transcription of DIG-labeled PTPIP 51-cRNA was performed using the 10 × RNA-DIG Labeling-Mix (Boehringer Mannheim; Mannheim, Germany) and RNA-polymerases T3 and SP6. Vectors containing the PTPIP 51 inserts were digested with NcoI and NotI (New England Biolabs; Frankfurt, Germany) for the production of sense cRNA and antisense cRNA, respectively.
ISH was performed as described previously (Steger et al. 1998, 2000) using a stringency to achieve hybridization only with a homology >80%. Briefly, 5-μm sections were partially digested with proteinase K, postfixed in 4% paraformaldehyde, and exposed to 20% acetic acid. After prehybridization in 20% glycerol, sections were covered with the DIG-labeled sense or antisense cRNA probes.
Both cRNAs were used in a dilution of 1:100 (the optimal concentration was determined by a dot blot test) in hybridization buffer containing 50% deionized formamide, 10% dextran sulfate, 2 × saline sodium citrate (SSC), 1 × Denhardt's solution, 10 μg/ml salmon sperm DNA, and 10 μg/ml yeast tRNA. Hybridization was performed overnight at 37C in a humidified chamber containing 50% formamide in 2 × SSC. Post-hybridization washes were performed, according to Lewis and Wells (1992). After blocking with 3% BSA, sections were incubated (overnight at 4C) with the anti-DIG Fab-antibody conjugated to alkaline phosphate (Boehringer Mannheim). Staining was visualized by developing sections with nitrobluetetrazolium/5-bromo-4-chloro-3-indolylphosphate in a humidified chamber protected from light. For each test, negative controls were performed using DIG-labeled cRNA sense probes.
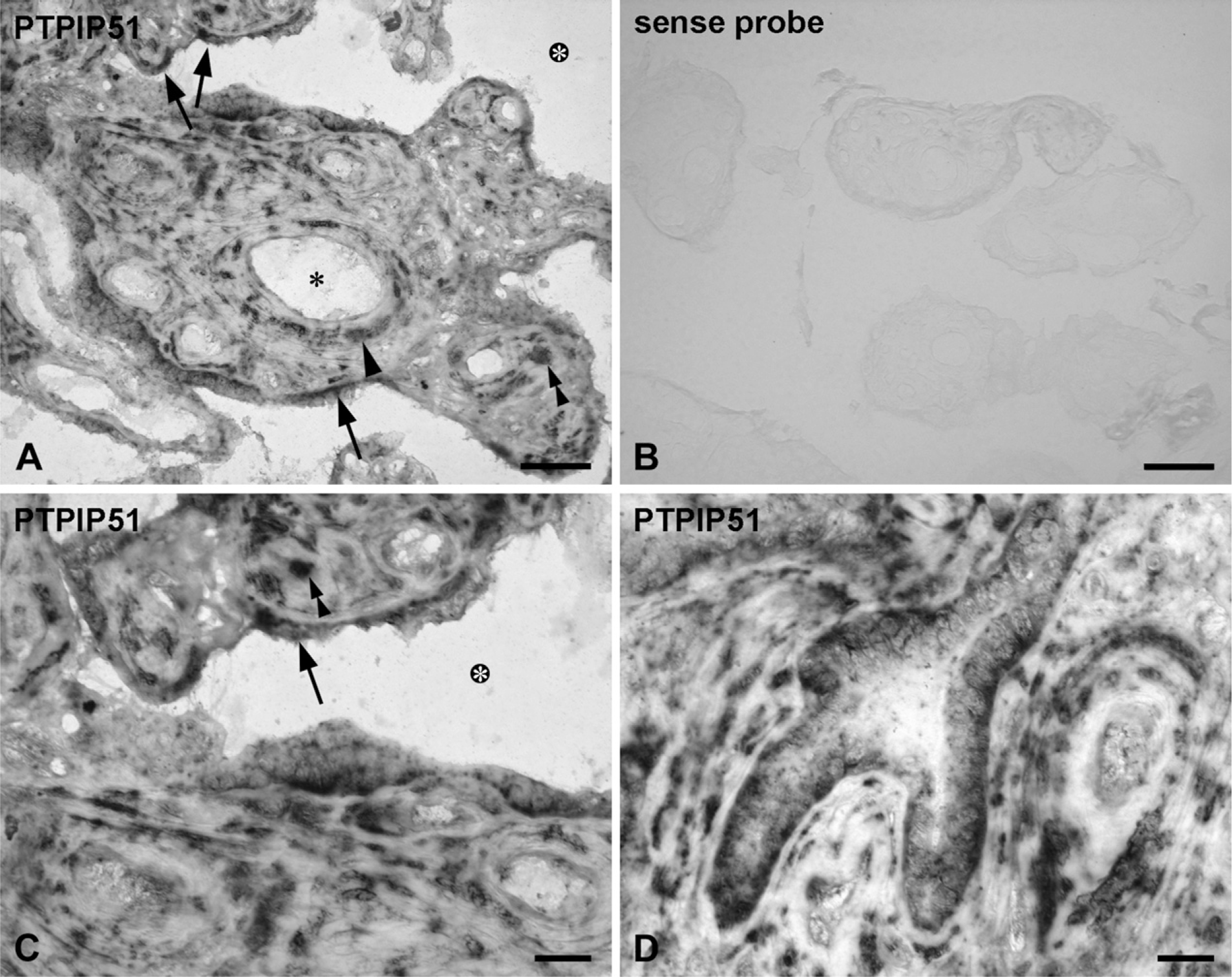
ISH of PTPIP51 in human term placenta.
RNA Extraction
RNA extraction from cryomaterial was performed using the RNA extraction kit RNeasy MINI (Qiagen).
First Strand Synthesis
First strand synthesis was performed using Omniscript according to the manufacturer's protocol (Qiagen).
PCR
PCR was performed on an iCycler using SYBR Green Supermix (BioRad; Munich, Germany). Per sample, 2 μl cDNA was used for amplification of PTPIP51. Cycling conditions were 95C for 3 min, followed by 40 cycles of 95C for 30 sec, 55C for 30 sec, and 72C for 30 sec. The following primers were used (MWG; Ebersberg, Germany): 5′-GCAGGTGGTGCTATCAGGTC-3′ as the forward primer and 5′-AGCTCCAGGGCCAACTTCATC-3′ as the reverse primer, resulting in a 232-bp amplification product.
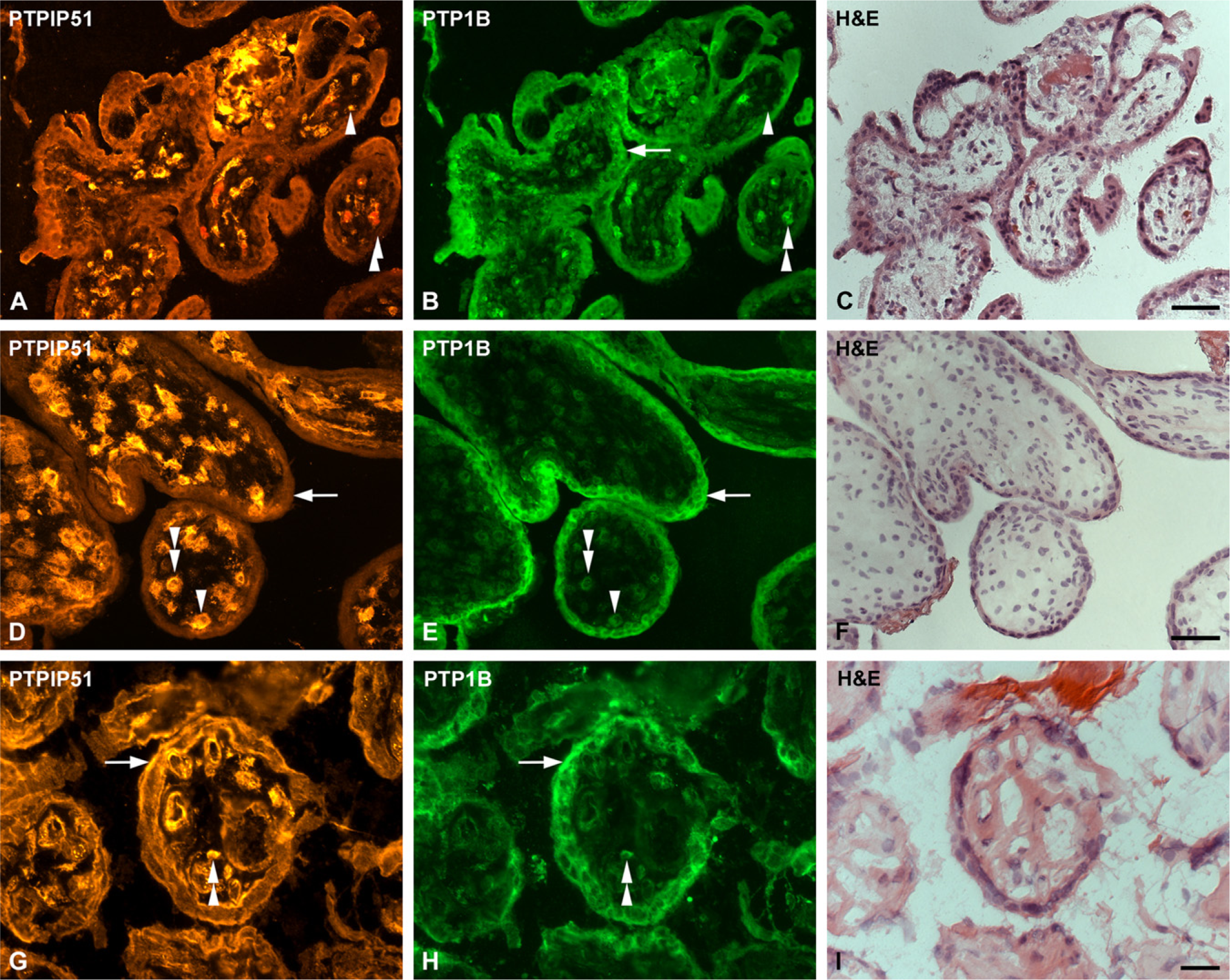
Double immunostaining of PTPIP51 and protein tyrosine phosphatase 1B (PTP1B) in sections of human placenta.
PCR products were visualized by agarose gel electrophoresis. While amplification of a 90-bp β-actin product served as positive control, negative controls included samples lacking RT.
Immunoblotting
Samples of term placentae were separated on a 10% SDS-PAGE gel. Transfer on an Immobilon P membrane (Millipore; Danvers, MA) was performed according to Towbin et al. (1979). The membrane was blocked with 10% fat-free milk powder in PBS and subsequently incubated for 1 hr with antibodies diluted in 0.5% fat-free milk powder in PBS. Incubation with polyclonal anti-PTPIP51 was done overnight at 4C. Alkaline phosphatase-conjugated anti-rabbit immunoglobulins were applied for 1 hr at room temperature, and the reaction was visualized with the SigmaFast 5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium substrate (Sigma-Aldrich; St Louis, MO). A prestained molecular mass marker (Cat. No. SM0431; Fermentas, Burlington, ON, Canada) was used for calibration.
Results
RT-PCR analyses showed the presence of PTPIP51 mRNA in three different samples of placental tissue (Figure 1, right panel). β-actin amplification served as an internal positive control. Immunoblotting of term placentae showed PTPIP51 protein of three different molecular masses of ∼52, 45, and 28 kDa (Figure 1, left panel).
The cell-specific expression of PTPIP51 mRNA in term placenta was detected through ISH (Figure 2). Figure 2B shows placental tissue probed with the sense mRNA serving as an internal control.
Whereas the syncytiotrophoblast of first and second trimester placental tissue showed only a sparse or even no reaction to the PTPIP51 antibody (Figures 3A and 3D), a fraction of villous syncytiotrophoblast of mature placenta strongly expressed PTPIP51 mRNA (Figures 2A, 2C, and 2D) and protein (Figure 3G).
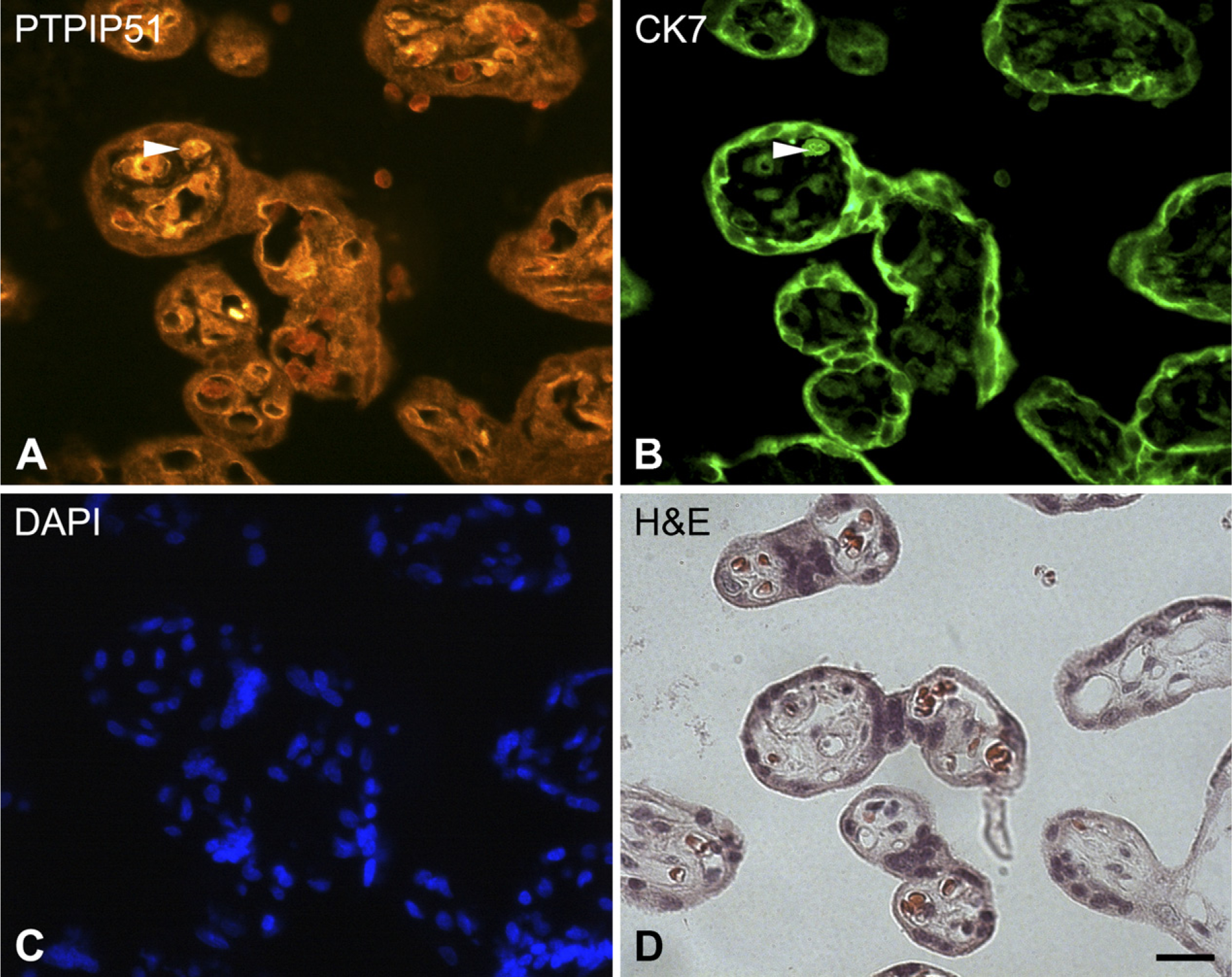
Double immunostaining of PTPIP51 and cytokeratin 7 in sections of human placenta.
In contrast to these findings, a converse expression pattern could be observed: for cytotrophoblasts, immunostainings of first and second trimester cytotrophoblasts showed a strong signal to the PTPIP51 antibody (Figures 3A and 3D), whereas samples of third trimester placentae showed a slightly reduced expression of PTPIP51 protein and mRNA (Figures 2A, 2C, 2D, and 3G). Double immunostainings with cytokeratin 7 antibody, a marker for trophoblast cells (Frank et al. 2000), substantiated the expression pattern of PTPIP51 in cytotrophoblast cells (Figures 4A and 4B).
Co-immunostaining experiments with PTPIP51 and its in vitro interacting partner PTP1B showed a constant and strong expression of PTP1B in the syncytiotrophoblast and a somewhat weaker expression in the majority of cytotrophoblasts during all trimesters (Figures 3B, 3E, and 3H). Hence, first and second trimester cytotrophoblasts were found to coexpress PTPIP51 and PTP1B.
Because of the PTPIP51 expression pattern described above, coexpression of the two respective antigens in the syncytiotrophoblast was only observed in mature placentae.
PTPIP51 could also be traced in arterial and venous small vessels and capillaries. The reaction to the PTPIP51 antibody was confined to smooth muscle cells, identified by α-smooth muscle actin (Figures 5A and 5B), and the endothelia, identified by van Willebrand factor (Figures 5D and 5E). This expression pattern was corroborated by ISH (Figure 2) and could be observed in all gestational stages. Interestingly, some placental macrophages identified by CD14 (Figures 4G and 4H), as well as some mesenchymal cells within the villous stroma, were found to be PTPIP51 positive. This could be observed for the first, second, and third trimesters.
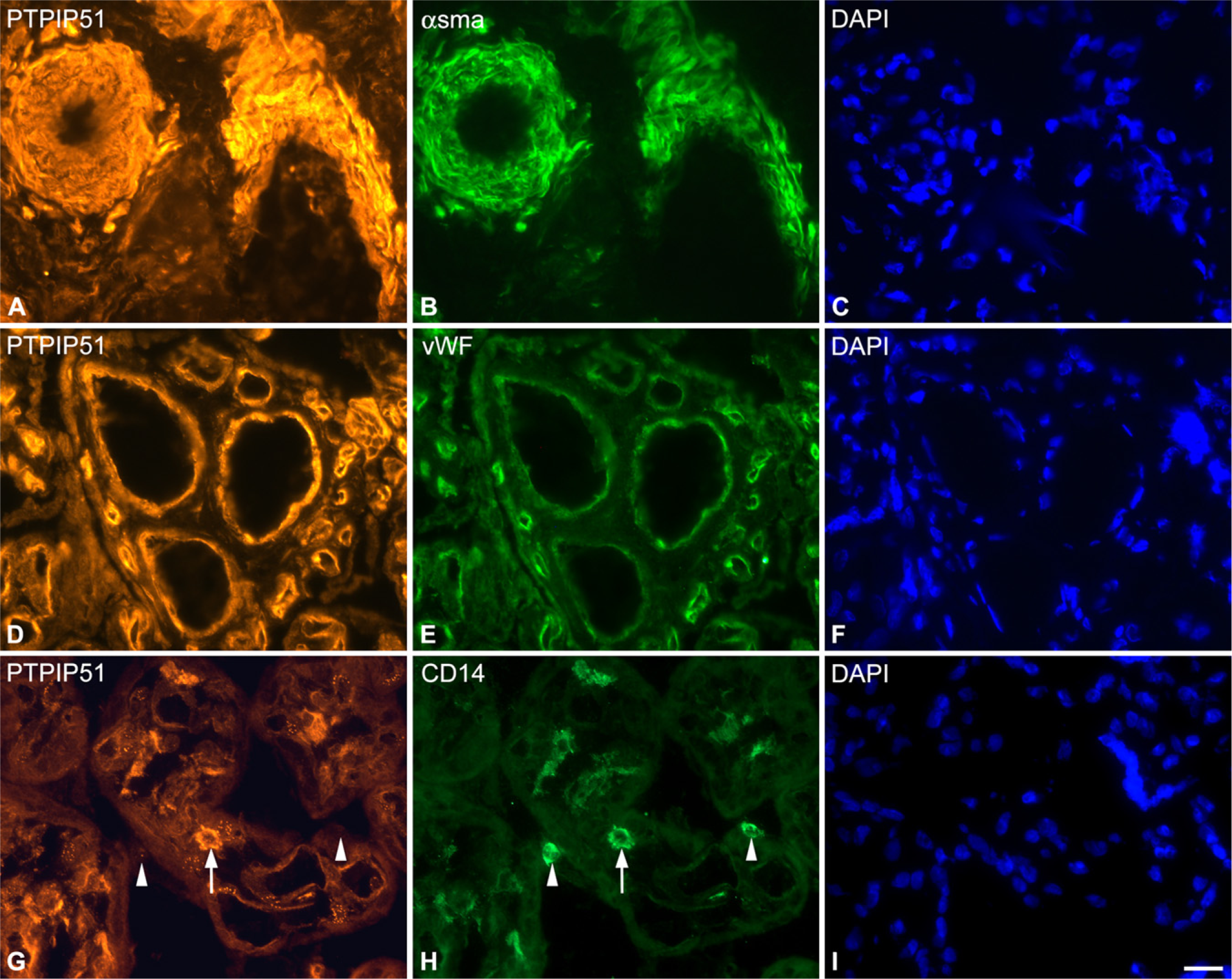
Double immunostudies of PTPIP51, α-smooth muscle actin (α-sma), van Willebrand factor (Vwf), and CD14 in sections of human term placenta.
PTPIP51 was found to be expressed in cells of the syncytiotrophoblast, undergoing late apoptosis, detected by TUNEL assay during different gestational stages. Moreover, a small population of apoptotic villous cytotrophoblasts showed strong PTPIP51 reactivity (Figures 6A-6J). Additionally, early apoptosis was detected by cytokeratin 18f antibody (Figures 7A and 7B) and by an antigen expression of caspase 3 (data not shown), thereby confirming the association of PTPIP51 to apoptosis.
Highly PTPIP51-reactive endothelial and smooth muscle cells were proliferative, as detected by Ki67 double staining (Figures 7D and 7E).
Discussions
This study showed for the first time the cell-specific expression of PTPIP51 and its in vitro interacting partner PTP1B in the human placenta during distinct weeks of gestation. Our data suggest a role for PTPIP51 in apoptotic and differentiation processes mandatory for the functional integrity of the syncytiotrophoblast. Moreover, PTPIP51 expression may also be important for placental macrophages (Hofbauer cells), as well as for the maintenance of vessels and capillaries, both expressing PTPIP51 during all weeks of gestation.
Villous cytotrophoblasts, arising from cytotrophoblastic progenitor cells, differentiate and fuse into the multinucleated syncytiotrophoblast layer (Lee et al. 2007). As meticulously studied by Huppertz and colleagues (Huppertz et al. 1998, 2006; Black et al. 2004) and other groups (Ho et al. 1999; Ka and Hunt 2003), this process is maintained by an elaborate program of delayed apoptosis, ultimately resulting in DNA degradation and the formation of syncytial knots that are released into maternal blood. Interestingly, these events share certain similarities with the regulated cell turnover of epidermal keratinocytes that express PTPIP51 and require activation of caspase 8 and 3, as well as the involvement of Src kinases and the JAK-STAT pathway. PTPIP51 was found to be expressed in cytotrophoblasts and also in cells of the syncytiotrophoblast that execute programmed cell death. The observation of a small number of apoptotic cytotrophoblasts has been described previously (Kar et al. 2007). Interestingly, these cells were highly reactive to the PTPIP51 antibody and may reflect the full-length form of PTPIP51, which has been shown to induce apoptosis when overexpressed in HEK293T cells (Lv et al. 2006). PTPIP51 may also serve as a signaling partner in earlier stages of apoptosis as shown in this study by immunostainings of caspase 3 and cytokeratin 18f.
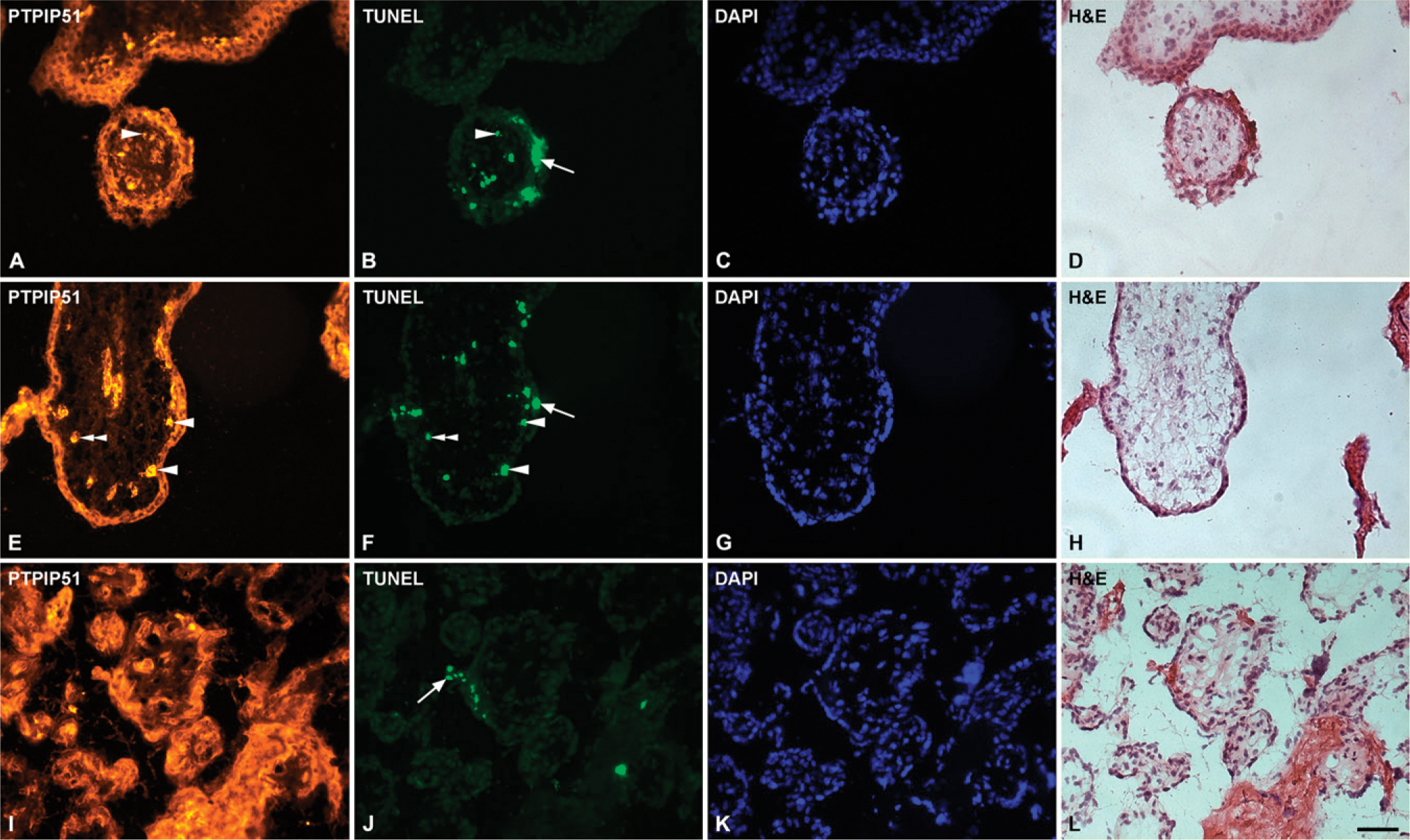
Double immunostaining of PTPIP51 and TdT-mediated dUTP-biotin nick end labeling (TUNEL)-labeled cells in sections of human placenta.
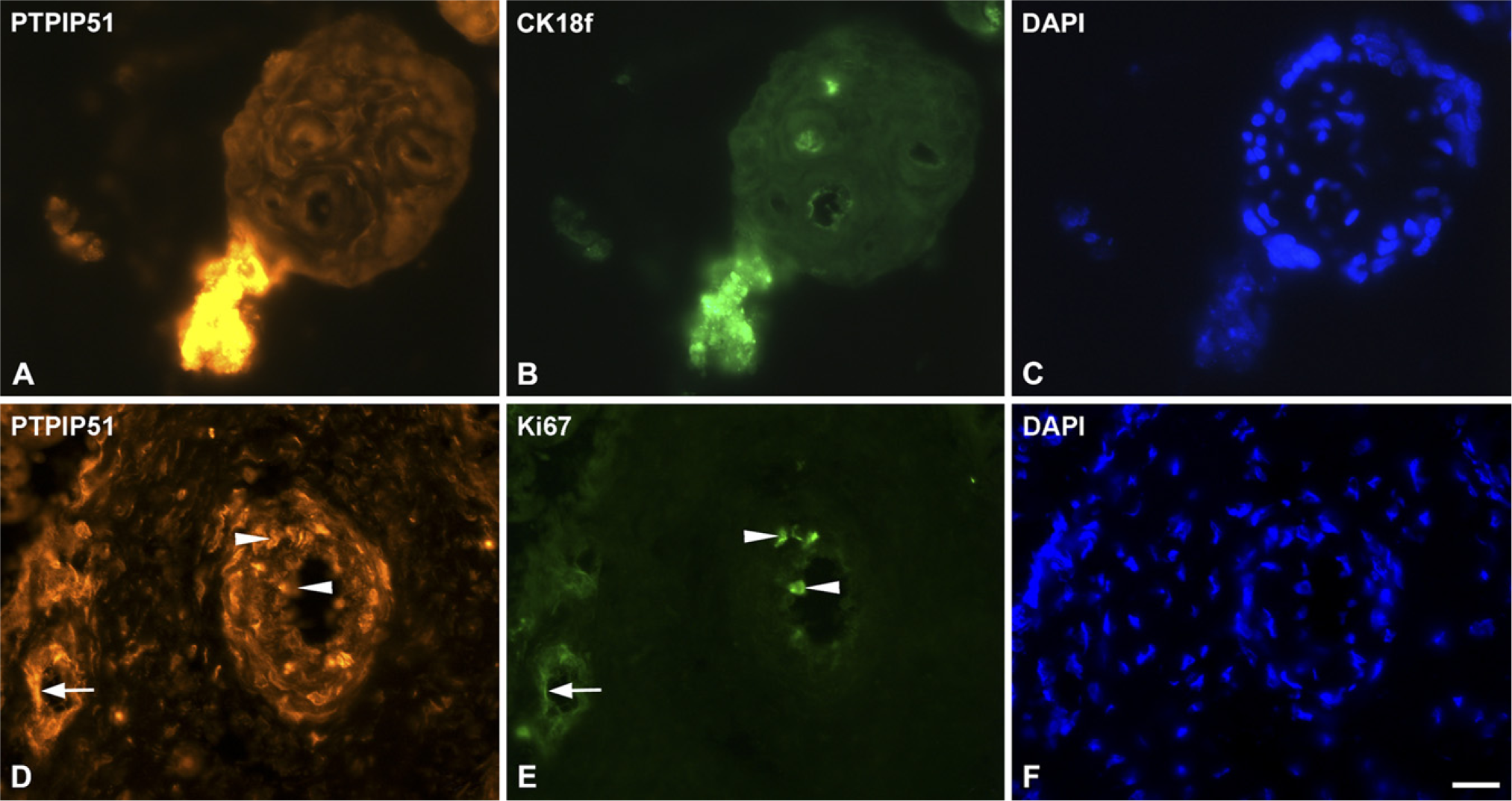
Double immunostaining of PTPIP51, Ki67, and cytokeratin 18f.
PTPIP51 reactivity was also traced in non-apoptotic cells of the cytotrophoblast and in the syncytiotrophoblast, indicating that PTPIP51 is not exclusively responsible for the induction of apoptosis but is also involved in processes of differentiation. These findings may be explained by putative isoforms or splicing variants, expressed in different tissues, that lack this N-terminal sequence, which is mandatory for the execution of apoptosis. This is also reflected by the different molecular mass (∼52, 45, and 28 kDa), detected by immunoblotting of PTPIP51. The known initiation sites in the Ptpip51 gene would lead to molecular protein masses of 52, 45, 38, and 30 kDa, which are close to the apparent masses of the PTPIP51 proteins determined by immunoblotting (Stenzinger et al. 2005).
Until now, no reports have been published regarding the histological localization of PTP1B in the human placenta, although its first description was released almost 20 years ago (Charbonneau et al. 1989). PTP1B, the prototype of protein tyrosine phosphatases, has emerged as an important signaling partner, regulating the immune system, cell cycle, and apoptosis and differentiation, and also plays a crucial role in cancer development and metabolic diseases (Tonks 2003; Dubé and Tremblay 2005; Koyama et al. 2006; Ostman et al. 2006; González-Rodriguez et al. 2007). Interestingly, the Src family kinases, interacting partners of PTP1B (Roskoski 2005; Julien et al. 2007), are reported to be involved in the regulation of trophoblast differentiation (Daoud et al. 2006). Thus, it is not unlikely that PTPIP51, being tyrosine-phosphorylated on Y176 by c-Src and v-Src kinase in vitro (Schreiner D, unpublished data), interacts with these enzymes, thereby promoting trophoblast differentiation.
Cytokine signaling, activating the JAK-STAT pathway, is also known to play a role in trophoblast differentiation. Because Ptpip51 transcription in the rat retina is regulated by CNTF (Roger et al. 2007), a member of the interleukin-6 family, one may speculate that transcription of the PTPIP51 gene in the human placenta is also dependent on cytokine stimulation.
Red-Horse et al. (2006) previously reported that human cytotrophoblasts are capable of inducing apoptosis in arterial smooth muscle and endothelial cells. PTPIP51 was expressed in arterial vessels of placental villi throughout all stages of gestation and may contribute to this process. Angiogenesis and subsequent vascular remodeling in general play a crucial role during placental development, involving numerous anti- and pro-angiogenetic factors. Among them, growth factor—driven signaling pathways, using tyrosine-phosphorylation, are essential for endothelial cell activation and proliferation, both being mandatory for capillary plexus formation and capillary sprouting (Carmeliet 2005; Red-Horse et al. 2006). Interestingly, during placental gestation, distinct cell types constitute a continuous source for vascular endothelial growth factor, which is first produced by villous cytotrophoblasts, whereas during further steps of angiogenesis, cytotrophoblasts, Hofbauer cells, and fibroblasts take over (Charnock-Jones et al. 2004; Demir et al. 2007). PTPIP51 is expressed in arterial and venous endothelia, as well as in the capillaries of human placenta, irrespective of gestational stage. Moreover, arterial smooth muscle cells and small populations of Hofbauer and mesenchymal cells were also PTPIP51 positive. Hence, PTPIP51 may be involved in signal transduction cascades, governing angiogenesis and vessel remodeling.
This study extended further expression analyses of our group and underlined the wide expression of PTPIP51 in many different cell types. Splicing variants and cell-dependent homologous and heterologous interactions of PTPIP51 (Schreiner D, unpublished data) may shed light on the function-dependent expression pattern.
Footnotes
Acknowledgements
We thank Claudia Tag for excellent technical assistance; Karola Michael for image editing; and Phillip Grant for linguistic revision.