
Abstract
Histochemical demonstration of tartrate-resistant acid phosphatase (TRAP) is used for the specific identification of osteoclasts. The enzyme, which we have shown to be critical for normal bone development in mice, is also characteristic of monohistiocytes, including alveolar macrophages, and is associated with diverse pathological conditions such as Gaucher's disease and hairy cell leukemia. TRAP activity is enhanced in serum when bone resorption is increased, and the activity is used routinely to monitor treatment responses in Gaucher's disease. We have lately shown widespread expression of the enzyme in murine tissues with particular reference to the skin, thymus, gut epithelia, and isolated dendritic cells, suggesting a possible role in immunity. To further clarify the significance of TRAP in human physiology, we have examined its distribution in non-skeletal human tissues and in CD34+-derived human dendritic cells. TRAP mRNA determined by Northern blotting analysis was expressed abundantly in spleen, liver, colon, lung, small intestine, kidney, stomach, testis, placenta, lymph node, thymus, peripheral blood leukocyte, bone marrow, and fetal liver. Expression of TRAP protein was investigated by immunohistochemistry, with which the enzyme was identified in multiple tissues. Histochemical staining detected enzymatically active protein in spleen, lung, skin, colon, stomach, and ileum. Active TRAP was identified in CD34+-derived immature dendritic cells and co-localized to intracellular CD63 positive organelles. When these cells were matured by induction with LPS, the TRAP activity increased fivefold and remained within the cell during the phase associated with CD63 surface expression. Our findings demonstrate widespread expression of TRAP in human tissues. Its abundant expression in epithelia and dendritic cells suggests a potential role in antigen processing and in immune responses.
Osteoclasts and other cells of monohistiocytic lineage, including the macrophage, express the band 5 isozyme of tartrate-resistant acid phosphatase, TRAP (EC 3.1.3.2) (Minkin 1982; Oddie et al. 2000). The isozyme is identified by its unique cathodal mobility at pH 4 and is further defined by its resistance to inhibition by L(+)-tartrate (Li et al. 1970). TRAP belongs to a class of purple proteins that contain a catalytically active di-iron-oxo cluster that is essential for its activity (Davis and Averill 1982). In addition to acting as a phosphatase, the enzyme is capable of generating oxygen radicals in the presence of hydrogen peroxide (Hayman and Cox 1994). Hydroxyl radicals are formed in the Fenton reaction (Sibille et al. 1987).
TRAP has long been used as a histochemical marker for the osteoclast (Burstone 1959), but it is clear from studies of its distribution that have emerged over the past decades that the enzyme is also expressed in other cells of macrophage lineage, and occasionally in lymphoproliferative disorders. The enzyme is secreted during bone resorption (Chambers et al. 1987) and active enzyme can be detected in the serum of mammals, where it exists as a complex with α2-macroglobulin (Shaffer–Brehme et al. 1999). Serum TRAP activity is pathologically increased in conditions in which bone resorption is enhanced, such as Gaucher's disease, hyperparathyroidism, osteoporosis (Lau et al 1987; Chamberlain et al. 1995), and some cancers (Lam et al. 1984). TRAP activity is also increased in fluid and tissues associated with the inflamed synovial joint (Luukkainen et al. 1990). Hairy cell transformation of B-lymphocytic leukemia cells is associated with enhanced expression of TRAP (Yam et al. 1972). Occasionally, cells from other lymphoproliferative disorders, such as prolymphocytic leukemia (Loffler et al. 1976)), lymphosarcoma cell leukemia (Katayama and Yang 1977), and Sezary syndrome (Naeim et al. 1979) show a positive reaction for TRAP. Activated human macrophages express the enzyme, particularly those of pulmonary alveolar origin (Efstratiadis and Moss 1985). TRAP has also been identified in human placenta (Ketcham et al. 1989; Janckila et al. 1996), where the cell type expressing the enzyme is unclear, and in odontoclasts that resorb enamel in human deciduous teeth before shedding (Sahara et al. 1998).
Previous studies of acid phosphatase function lacked specificity because of overlapping enzymatic activities of multiple phosphatase isozymes with the capacity to hydrolyze common substrates. The development of specific antibodies as well as gene probes for the Acp 5 isozyme of TRAP has greatly facilitated the systematic study of its enzyme expression in relation to tissue distribution and physiology.
The human TRAP gene maps to chromosome 19 and contains a putative N-terminal lysosomal leader sequence (Lord et al. 1990). In the macrophage, TRAP has a lysosomal localization (Hayman et al. 2000a). In the osteoclast, however, the protein has been thought to be secreted from lysosomes into the resorptive vacuole beneath the ruffled border (Reinholt et al. 1990). Recently, evidence has been presented suggesting that TRAP is not secreted from the ruffled border but is instead present in vesicles that are fused to transcytotic vesicles which transport matrix degradation products from the ruffled border to a specialized region in the basolateral membrane. The enzyme appears to be secreted from this intracellular source together with degradation products of the bone matrix (Halleen et al. 1999).
The enzyme has been purified from a number of sources, including human osteoclastomas, in which it is composed of two subunits initially translated as a single polypeptide (Hayman et al. 1989, 1991). Recent studies with recombinant TRAP species and TRAP purified from rat bone have demonstrated that the enzyme is synthesized as a latent proenzyme with low activity, which is activated by cysteine proteinases that cleave the single-chain 34 kD form into a highly active heterodimer in which the subunits are linked by a single interchain disulfide bond (Ljusberg et al. 2000). TRAP promotes the hydrolysis of a number of substrates, including nucleotides, aryl phosphates, and phosphoproteins (Hayman et al. 1989; Janckila et al. 1992; Nash et al. 1993).
Mice deficient in TRAP generated by selective gene targeting have demonstrated that TRAP is essential for the normal mineralization of cartilage in developing bones and for maintenance of the adult skeleton (Hayman et al. 1996). These knockout mice also display an impairment of macrophage function (Bune et al. 2001) with abnormal immunomodulatory cytokine responses. Transgenic mice overexpressing TRAP exhibit increased osteoclastic resorption of bone, which is largely compensated for by a reciprocal increase in the rate of bone formation (Angel et al. 2000).
In the mouse, TRAP is expressed in many tissues in addition to bone (Angel et al. 2000; Hayman et al. 2000a,b), and the identification of the enzyme in murine dendritic cells suggests a function in T-cell immunity (Hayman et al. 2000a). Because TRAP is also implicated in several human diseases, including those affecting the skeleton, as well as lymphoid organs and macrophages, we sought to investigate further the expression of TRAP in selected human tissues that participate in immunity and in isolated human dendritic cells.
Materials and Methods
Northern Blotting
Multiple-choice Northern blots were purchased from Cambridge Biosciences (Cambridge, UK) and Clontech Laboratories (Basingstoke, UK). Blots were made with 2 μg poly A+ RNA per lane from human tissues. The RNA samples were obtained from healthy adult tissues and from more than a single donor. The membranes from Cambridge Biosciences were wetted in 4 × SSC for 20 min and incubated for 2 hr at 65C in Rapid-hyb buffer (Amersham Pharmacia Biotech; Little Chalfont, UK). Human TRAP cDNA (25 ng) was labelled with [32P]-dCTP using the Prime-a-Gene Labeling System from Promega (Southampton, UK). The probe was purified down a column of Sephadex G-50 and added to the prehybridization solution with the blot. Hybridization was carried out for 2 hr at 65C using 2 × 106 cpm/100 μl. Blots were washed at the same temperature: three times for 5 min with 2 × SSC–0.1% SDS and subsequently twice in 0.25 × SSC–0.1% SDS for 30 min. Blots from Clontech were prehybridized in ExpressHyb at 68C for 30 min. This solution was replaced with radiolabeled probe as previously described in fresh ExpressHyb and incubated for 1 hr at 68C. Washes were for 30–40 min at room temperature (RT) in 2 × SSC–0.05% SDS, followed by twice for 20 min at 50C in 0.1 × SSC–0.1%SDS. All membranes were exposed to Hyperfilm TM-βmax (Amersham Pharmacia Biotech) at –80C for 4 days.
Histochemical Staining for TRAP
Unfixed, undecalcified cryostat sections were stained for TRAP activity using the standard naphthol AS-BI phosphate postcoupling method, using Fast Red as the coupler. Tissue samples were obtained during the course of surgical biopsy for unrelated disorders in separate adult subjects and were found to be disease-free after histological examination. The incubation was carried out for 10 min at RT in 0.4 M sodium acetate buffer, pH 5.6, containing 2 mM naphthol AS-BI phosphate and 100 mM sodium tartrate. The reaction was stopped in distilled water and postcoupled in the same buffer containing 2 mM Fast Red for 2 min or until color developed, followed by washing in distilled water (Burstone 1958).
Immunohistochemistry
Immunohistochemistry was performed using a Vectastain Elite ABC kit and peroxidase staining from Vector Laboratories (Peterborough, UK). Frozen sections obtained from disease-free tissue of adult individuals after surgical biopsy were fixed in acetone at 20C for 20 min. Formalin-fixed, paraffin-embedded sections were treated with two changes of xylene for 10 min each with constant stirring. Sections were rehydrated through a series of ethanol concentrations and washed with water. All sections, irrespective of their method of preparation, were washed in PBS for 5 min and incubated for 20 min in diluted normal blocking serum prepared from the species in which the secondary antibody was made. Excess serum was blotted from sections and primary antibody diluted in PBS was applied. The primary antibodies were polyclonals: rabbit anti uteroferrin (Echetebu et al. 1987) and goat anti-B7 (V17; Santa Cruz Technology, Santa Cruz, CA). Nonimmune immunoglobulins were used as controls. After an overnight incubation at 4C the slides were washed twice in PBS. Diluted biotinylated secondary antibody was applied for 30 min, followed by washing in PBS as before. An avidin and biotinylated horseradish peroxidase macromolecular complex (Vector Laboratories; based on the methods of Hsu et al. 1981a,b) was added to the sections and left for 30 min. After washing in PBS, peroxidase substrate was applied to the sections and incubated at RT until the desired intensity developed. The slides were rinsed in tapwater, counterstained with hematoxylin, and mounted.
Harvesting and Culture of Human Dendritic Cells
CD34+ stem cells were immunomagnetically purified from leukapheresis products obtained from disease-free adult donors for hematopoietic stem cell storage and transplantation with a midiMACs system (Miltenyi Biotec; Bisley, Surrey, UK). The cells were cultured in 24-well plates in serum-free medium containing GM-CSF, TNFα, TGFβ-1, stem cell factor, and F1t3 ligand (Peprotech) (Strobl et al. 1997; Gatti et al. 2000). After a 10-day incubation, clusters were purified on a 7.5% BSA column. These clusters contained approximately 95% immature dendritic cells. For activation of these cells, the media was supplemented with LPS (Sigma-Aldrich; Gillingham, UK) to promote differentiation into mature dendritic cells.
Confocal Microscopy of Human Dendritic Cells
Dendritic cells were cultured on alcian blue-coated coverslips and fixed in methanol/acetone (1:1) at –20C overnight. Cells were washed three times with PBS–0.1% BSA and incubated with mouse antibody directed to the CD63 antigen (Bio-design; Kennebunk, ME) and rabbit antibody to porcine uteroferrin (Echetebu et al. 1987) overnight at 4C. Slides were washed three times with PBS–0.1% BSA and incubated at RT with FITC-conjugated and Texas Red-conjugated secondary antibodies (Jackson; Avondale, PA). Cells were washed three times with PBS–0.1% BSA and mounted with Mowiol (Calbiochem; Nottingham, UK). Cells were examined with a Leica confocal microscope.
Assay of TRAP Activity in Dendritic Cells and Tissues
Dendritic cells and human spleen were homogenized in 0.5 m1 0.4 M sodium acetate, pH 5.6, containing 1% w/v Triton X-100. To assay specifically for TRAP (band 5), immunoabsorption with immobilized rabbit antibodies to porcine uteroferrin was carried out (Echetebu et al. 1987) after neutralizing 13,000 × g supernatant tissue extracts to pH 7.5 with Tris base. TRAP activity was finally determined spectrophotometrically before and after immunoabsorption in the presence of 0.1 M L(+) sodium tartrate at pH 5.6 using 10 mM 4-nitrophenyl phosphate as substrate (Hayman et al. 1989). Assays were carried out in duplicate using at least two dilutions each of the tissue or cell extracts; individual enzymatic determinations were within 6% of the mean value.
Results
Northern Blotting Analysis
An investigation of TRAP mRNA expression in 17 human tissues was carried out by Northern blotting analysis (Figure 1). Abundant expression of TRAP transcripts of the expected size of 1.5 kb was identified after hybridization of radiolabeled full-length human TRAP cDNA in spleen, liver, lung, kidney, stomach, lymph node, bone marrow, and fetal liver, with smaller amounts in colon, small intestine, placenta, and thymus, and a trace in testis and peripheral blood leukocyte. Transcripts were absent in muscle, brain, and heart. The isozyme-specific activities of TRAP in immature human dendritic cells and mature dendritic cells were respectively 0.053 and 0.245 μmols/mg protein/minute, indicating a substantial increase on maturation in vitro. Therefore, the activity of the enzyme is eightfold greater in dendritic cells than in human spleen but increases further on maturation to a value approximately 40-fold greater in mature dendritic cells.
Enzymatic Activity of TRAP in Tissues
An extract of human spleen exhibited a TRAP activity of 0.006 μmols/mg protein/min. Histochemical examination of tissues for TRAP activity revealed a positive enzymatic reaction in several tissues, especially lung, where high activity was widespread (Figure 2A). Activity was present in the skin, with higher activity concentrated around the hair follicles than in the epidermis (Figures 2B and 2C). Here enzyme activity was detected only after prolonged incubation in vitro (see arrows). Weak enzymatic activity was also demonstrated in colon, ileum, stomach, liver, kidney, and spleen (not shown).
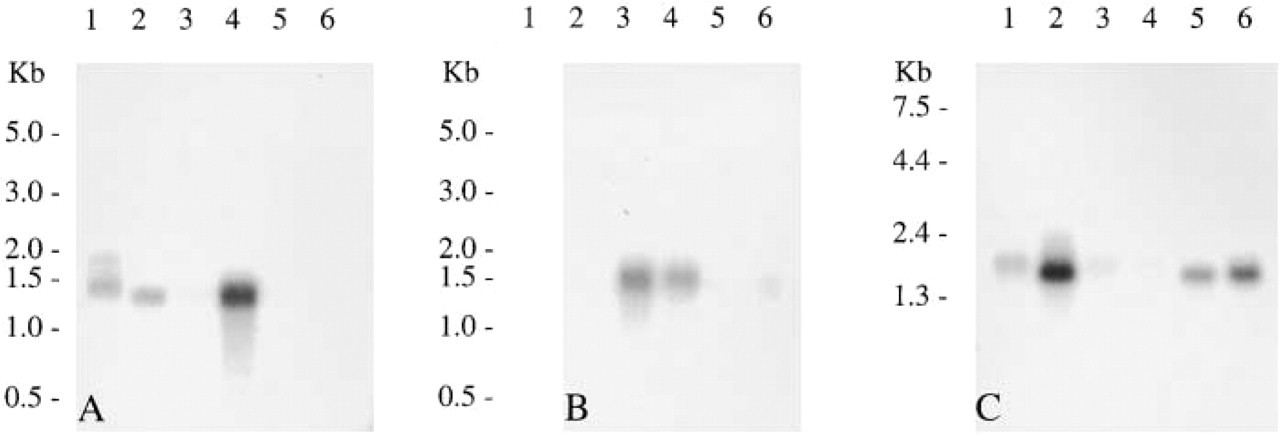
TRAP expression in human tissues. Northern hybridization of human poly A+ RNA with a radiolabeled human TRAP cDNA showing positive transcripts of 1.5 kb. Lanes: Al, spleen; A2, liver; A3, colon; A4, lung; A5, small intestine; A6, muscle; B1, brain; B2, heart; B3, kidney; B4, stomach; B5, testis; B6, placenta; C1, spleen; C2, lymph node; C3, thymus; C4, peripheral blood leukocyte; C5, bone marrow; C6, fetal liver.
Immunohistochemistry
An investigation of TRAP by immunohistochemistry identified protein in several tissues (Figures 3 and 4). In the lung, staining was in cells surrounding the alveolar spaces, comparable with expression in pulmonary alveolar macrophages as well as sparse dendritic cells (Figure 3A). Abundant protein immunostaining was detected in sinusoidal cells surrounding the blood spaces around the vessels in the spleen (Figure 3B). In the liver (Figure 3C), immunohistochemical staining gave a positive reaction in hepatocytes surrounding the hepatic venous radicals. In the stomach, colon, and ileum (Figures 4A-4C), TRAP protein was concentrated in the muscular layer of the submucosal epithelium. In the uterus (Figure 4D), TRAP immunostaining was prominent around the secretory ducts and endometrial glands.
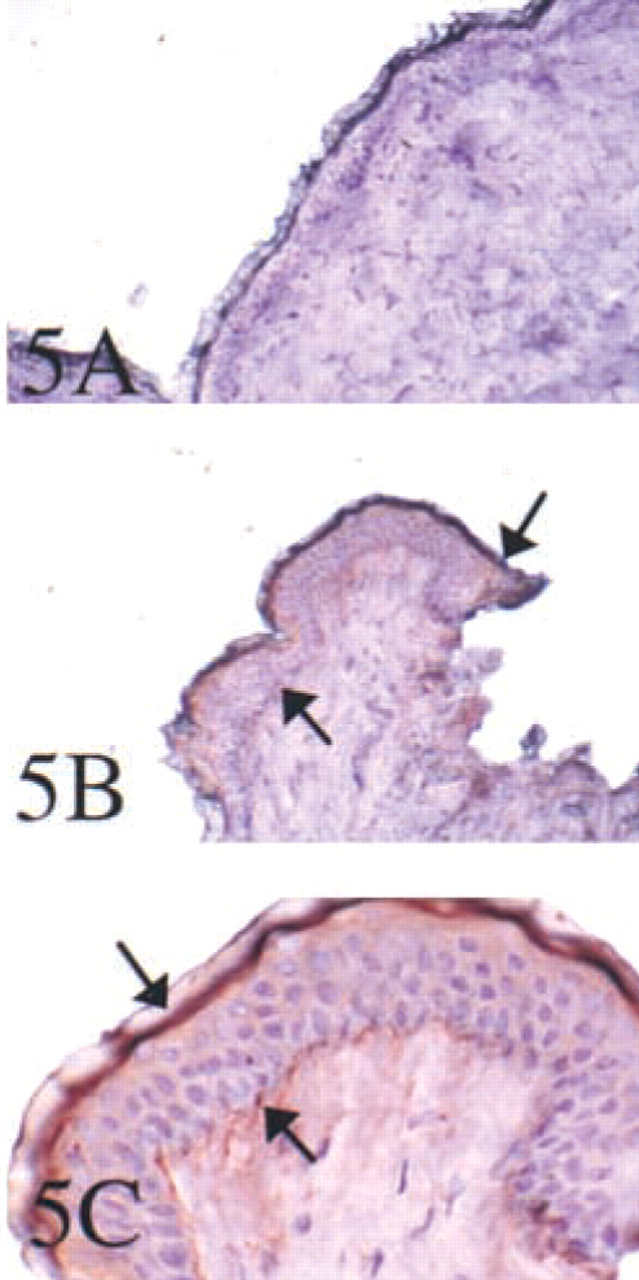
In the skin, as in the mouse, a dual localization of TRAP protein was demonstrated by immunostaining. TRAP protein was identified with strong staining reactions in cells of the stratum basale, in regions of the developing papillae, and in a subepithelial layer to which Langerhans cells migrate (Figure 5).
TRAP Expression in Dendritic Cells
The presence of TRAP in CD34+-derived human dendritic cells was investigated. Human dendritic cells were cultured from CD34+ stem cells obtained at leukapheresis after stem cell mobilization with GM-CSF. Both immature and activated dendritic cells stained positively for TRAP (Figure 6). In immature dendritic cells, as defined by low cell surface expression of MHC class II molecules CD83 and CD86, the TRAP protein co-localized to intracellular CD63-positive organelles. After activation with LPS, the dendritic cells mature into an MHC class II CD83 and CD86 high-expressing population. In these cells, the TRAP protein remained intracellular, whereas CD63 was expressed at the cell surface. Isolated dendritic cells were assayed for TRAP activity after lysis. Activity that could be removed by immobilized antibody was fivefold higher in the activated dendritic cell population compared with the immature cells.
Discussion
Because dendritic cells have been recognized as the professional antigen-presenting cells of the immune system and because of our earlier demonstration that Acp 5 shows widespread expression in murine tissues and specifically can be demonstrated in isolated dendritic cells, a comparative study using human tissues was undertaken to determine the potential involvement of Acp 5 in the human immune system, including tissues with epithelia that interface with the external environment.
We have observed that TRAP expression is widely distributed in human tissues, despite having long been employed as a familiar marker of the osteoclast. Osteoclasts indeed show intense staining for TRAP in normal bone (Burstone 1959), unlike other normal tissues except the lung, in which phagocytosis of airborne microbes and other inhaled particulates is active. The enzyme has been reported in human pulmonary alveolar macrophages (Efstratiadis and Moss 1985). Human TRAP activity is greatly enhanced in splenic macrophages in particular storage diseases, such as Gaucher's disease, in which steady-state levels of TRAP mRNA are at least 10-fold increased, presumably in response to activation induced by the presence of the undigested storage material (Lord et al. 1990; Moran et al. 2000). Studies in the rat have reported phosphoprotein phosphatase activity with biochemi- cal properties corresponding to TRAP in the skin. As in our findings, the enzyme was detected in granular cells and the lower strata of the comified epidermis and was also localized in keratohyaline granules (Ohno et al. 1989). These authors did not remark on the presence of TRAP expression in the Langerhans cells of rat epidermis but suggested that the enzyme dephosphorylates histidine-rich proteins, such as filaggrin, implicated in the control of epidermal differentiation. We recently reported on the expression of TRAP in murine skin, where it was abundant in both the epidermis and in Langerhans cells which are of dendritic origin (Hayman et al. 2000a,b). Striking expression of TRAP (Acp 5) mRNA was also demonstrated in the skin and epithelial surfaces of the oropharyngeal and gastrointestinal cavities by in situ hybridization of developing mouse embryos.
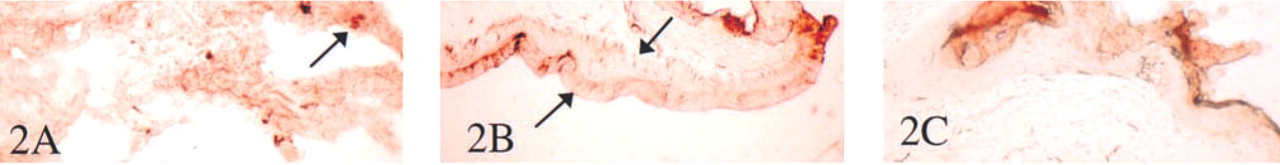
(
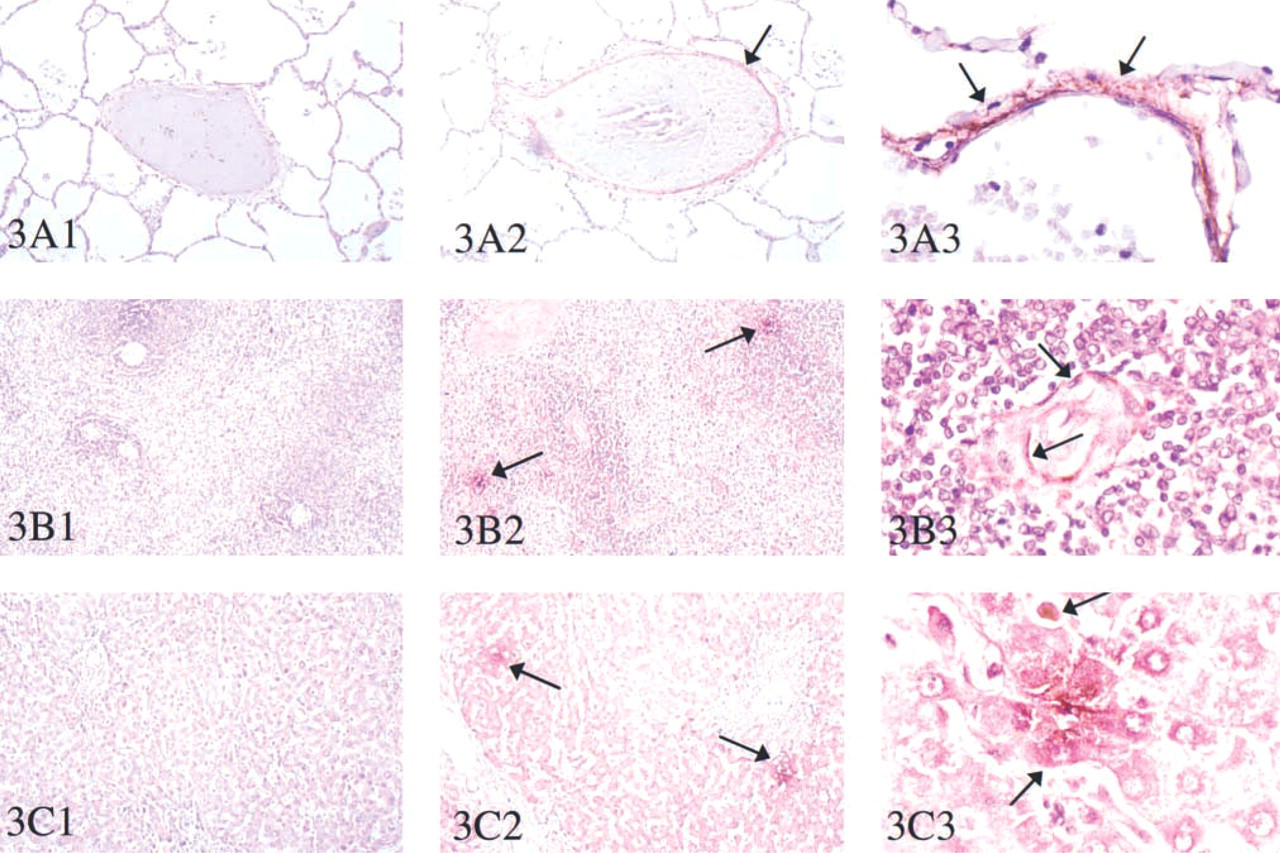
Immunohistochemistry of tartrate-resistant acid phosphatase in human tissues. Immunohistochemistry was performed using peroxidase staining with Vector Red substrate. The primary antibody was a polyclonal rabbit anti uteroferrin used in paraffin sections of lung (
In view of these findings and of the importance of dendritic cells in human immunity, we report here multi-tissue expression of Acp 5 in human organs at sites corresponding to cells of hematopoietic lineage and, specifically, in cells of dendritic phenotype invested with antigen-presenting properties. A striking finding was the extent to which TRAP expression localized to tissues and organs with environmentally exposed surfaces, including the skin, lung, gastrointestinal tract, and endometrium. These are clearly sites at which continual interactions occur between dendritic phagocytes and microbial and other antigens. TRAP expression, as expected, was also detected in visceral tissues that contain macrophages and also contribute component cells of the mononuclear phagocyte system, including the spleen and liver. The skin revealed a particularly interesting dual localization, especially around the hair follicle and appendages.
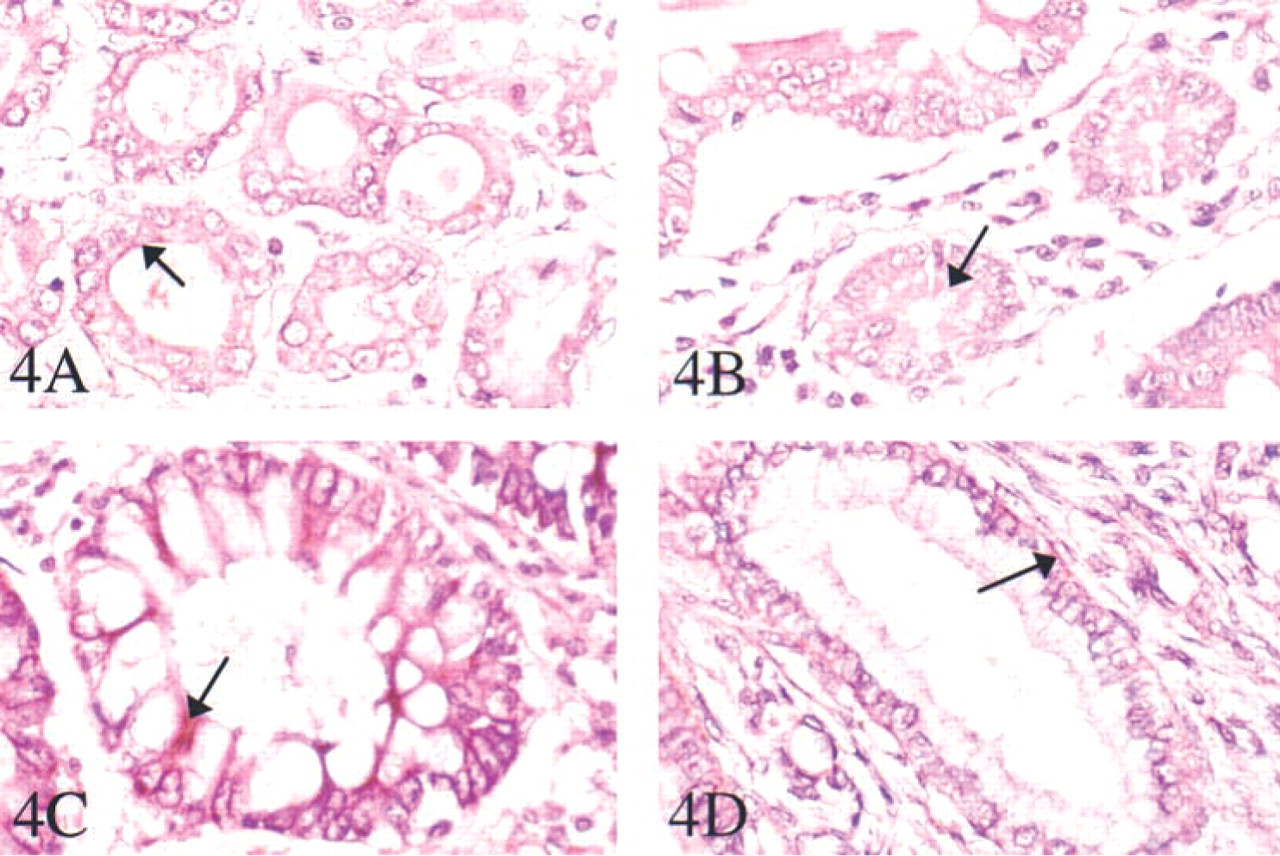
Immunohistochemistry of TRAP in human tissues. Immunohistochemistry was performed using peroxidase staining with Vector Red substrate. The primary antibody was a polyclonal rabbit anti-uteroferrin used in paraffin sections of stomach (
(
In skin and in tissues lining the gastrointestinal tract, TRAP activity was unexpectedly low compared with the amount of protein present, as detected by the strong immunostaining reactions and confirmed by the studies of steady-state transcript abundance. This may be in readiness for activation when triggered by appropriate conditions. The cysteine proteinases are a family of proteases of which members are present in all sites at which TRAP has been identified. In vitro studies show that TRAP enzyme is greatly activated on cleavage of the single-chain pro-enzyme into the disulfide-linked heterodimer by cysteine proteinases (Ljusberg et al. 2000). We propose that this process occurs in vivo, thus permitting activation of latent TRAP at privileged sites when required.
Here for the first time we have identified abundant TRAP expression in cultured human dendritic cells. Located in most tissues, DCs capture and process antigens. They upregulate their co-stimulatory molecules and migrate to the spleen and the lymph nodes, where they activate antigen-specific T-cells (Banchereau and Steinman 1998). We show that the specific enzymatic activity of TRAP in mature DCs was fivefold higher than in immature cells. This may explain why TRAP activity is low in skin. Skin contains many migratory immature dendritic cells or Langerhans cells that are located above the basal layer of proliferating keratinocytes. Immature DCs are weak T-cell stimulators; they express few MHC and accessory molecules but possess many receptors for antigen capture on their membrane surfaces. On maturation, DCs produce large amounts of antigen MHC class II molecules where antigens undergo proteolytic digestion into peptides for immune presentation. This takes place largely in the specialized MHC class II-rich compartments that are abundant in immature DCs. On maturation, these compartments are converted to non-lysosomal vesicles that discharge their MHC–peptide complexes to the surface. The CD63 marker used in our studies is a lysosomal membrane glycoprotein that is translocated to the plasma membrane after dendritic cell activation. This explains why, in immature DCs, TRAP co-localized with the DC marker CD63, whereas in mature cells TRAP remained intracellular and CD63 staining was translocated to the cell surface.
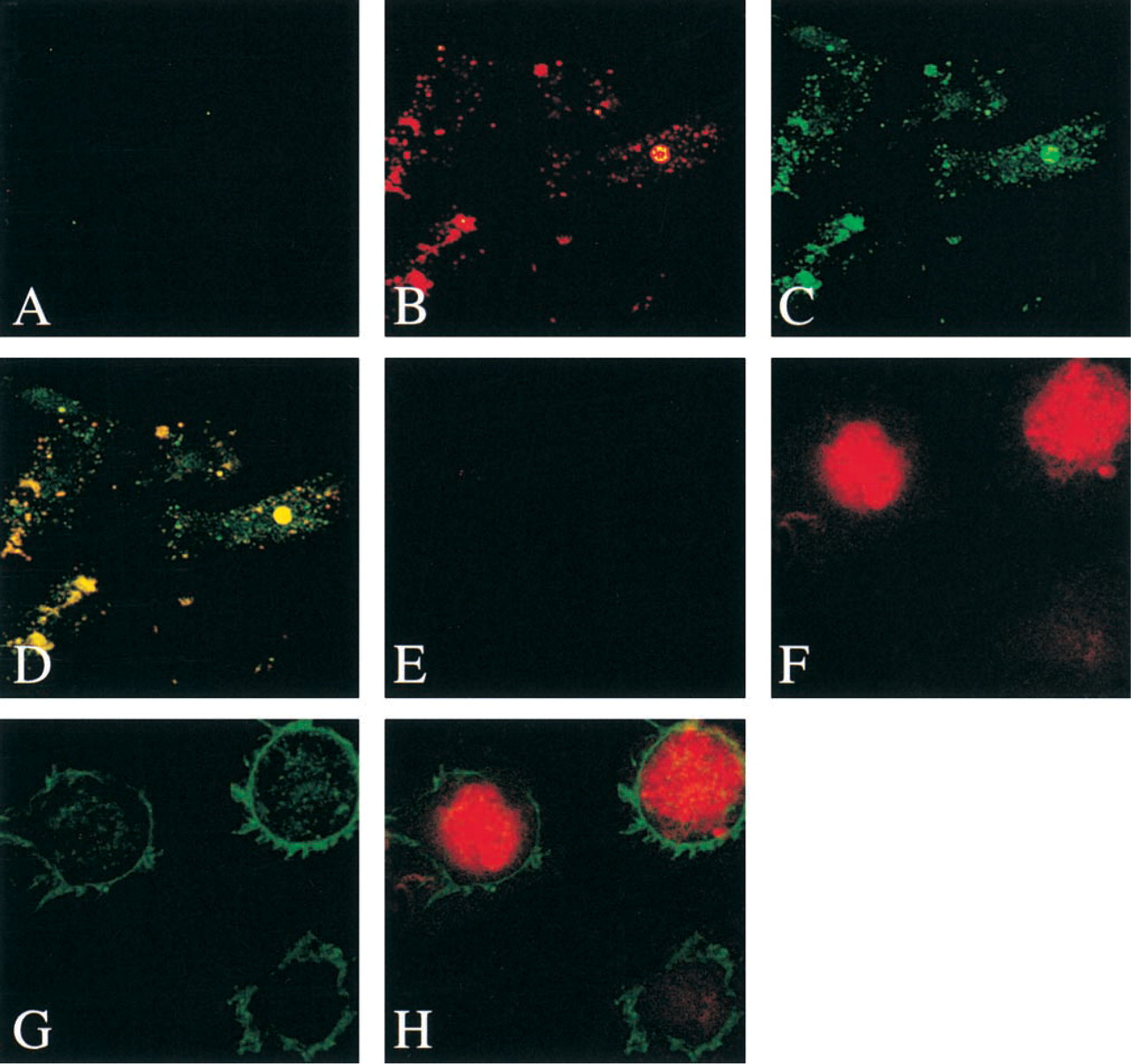
Identification of TRAP in human dendritic cells. Anti-uteroferrin and CD63 staining of immature and LPS-activated human dendritic cells. In mature DCs (
Acp 5 expression in dendritic cells within the muscular layer of the submucosa of the gastrointestinal tract is also at a site where a putative role in cells that mediate antigen capture and processing is likely. In the skin, non-keratinocyte epidermal cells, especially migratory Langerhans cells, are involved in presentation of processed foreign antigens to the T-lymphocytes. Pulmonary alveolar macrophage cells are activated by continual exposure to environmental antigens.
In summary, these studies in humans and previous studies carried out in mice and rats (Hayman et al. 1991, 2000a,b; Reddy et al. 1995; Angel et al. 2000) demonstrate that TRAP is widely distributed in normal tissues. Immunolocalization studies show that the enzyme is expressed in dendritic cells localized to the skin, spleen, thymus, and lymph nodes, as well as epithelial surfaces of the gastrointestinal tract. It therefore appears that the Acp 5 isozyme of TRAP hitherto regarded as a selective marker for osteoclast and certain activated macrophage populations, is in fact a general reagent for the study of mononuclear phagocytes and dendritic cells of bone marrow origin. The consistent expression of this enzyme and its activation by lysosomal proteinases in professional antigen-presenting cells immediately suggests a role in the immune response. Experiments to determine the functional link between TRAP and the systems of innate and acquired immunity are in progress. Our preliminary studies conducted with mice lacking the enzyme as a result of targeted gene disruption indicate an abnormal immunomodulatory response and cytokine secretion profile in cultured bone marrow and peritoneal macrophages, which endorse the predicted relationship between TRAP expression and these immune networks (Bune et al. 2001).
Footnotes
Acknowledgements
ARH is in receipt of a Research Fellowship from the Arthritis Research Campaign. PM and PJL are supported by The Wellcome Trust.
Joan Grantham and Philip Ball kindly prepared the manuscript and figures.