
Abstract
Oxygen insensitivity of the histochemical assay to detect glucose-6-phosphate dehydrogenase (G6PD) activity with NT as tetrazolium salt has been proved to be a powerful tool to discriminate various types of adenocarcinoma from normal tissues. Here we investigated whether this phenomenon can also be applied to differentiate between chemically induced hepatocellular (pre)neoplasms and normal liver tissue in rats. Residual activity (percentage of the amount of final reaction product that is generated in oxygen and that is generated in nitrogen) was 60% in (pre)neoplastic cells and 6% in normal liver parenchymal cells. This means that the oxygen insensitivity test is a useful tool to distinguish (pre)neoplasms from normal rat liver tissue. N-Ethylmaleimide, a blocker of SH groups, did not affect G6PD activity in (pre)neoplastic cells, whereas activity in normal cells was reduced by half. Therefore, the absence of essential SH groups in G6PD in (pre)neoplastic cells is held responsible for the oxygen insensitivity phenomenon. We conclude that oxygen insensitivity of the histochemical assay for G6PD activity is a fast, easy, and cheap tool to diagnose (pre)neoplasms in rat liver. Discrimination is likely to be based on altered properties of the enzyme in (pre)neoplastic cells.
(
Keywords
H
To discriminate between malignant cells and non-malignant cells, the histochemical assay of G6PD is performed with the tetrazolium salt neotetrazolium chloride (NT) (Van Noorden and Frederiks 1992; Van Driel and Van Noorden 1999). The method is based on the capture of electrons that are produced by the enzyme when it oxidizes its specific substrate. With the help of an electron carrier (1-methoxy)phenazine methosulfate (mPMS), these electrons can be picked up by the tetrazolium salt, which is then reduced into a water-insoluble strongly colored formazan. Oxygen is able to interfere in this reduction process when NT is used as tetrazolium salt (Butcher 1978). When the assay is performed in an oxygen-rich environment, hardly any formazan is formed in normal cells, whereas the amount of formazan in cancer cells is less affected by oxygen. Therefore, G6PD activity in normal cells is called oxygen sensitive and G6PD in cancer cells oxygen insensitive. The best measure for the oxygen sensitivity/insensitivity phenomenon is the residual acitivity (RA), which is defined as the percentage of the amount of formazan produced by G6PD activity in oxygen and that produced in nitrogen. When RA in cells is lower than 20%, these cells are considered oxygen sensitive and therefore non-malignant. Cells are considered oxygen insensitive and therefore malignant when the RA is higher than 20% (Best et al. 1990; Griffini et al. 1994; Van Driel et al. 1997, 1999b).
Certain metabolic changes in cancer cells are held responsible for the oxygen insensitivity phenomenon, such as a combination of elevated G6PD activity, decreased superoxide dismutase activity, and decreased lipid peroxidation capacity (Best et al. 1990; Griffini et al. 1994; Van Driel and Van Noorden 1999).
In this study we investigated in rats whether the oxygen insensitivity of the histochemical assay of G6PD activity could be used for diagnosis of HCC. Furthermore, we investigated further chemical backgrounds of the oxygen insensitivity test in these cells and focused on a possible role of changes in SH groups in the active site of G6PD.
Materials and Methods
Induction of HCC
HCC was induced in livers of 15 male Wistar rats by administration of 0.01% diethylnitrosamine (Sigma: www.sigmaaldrich.com, St Louis, MO) via drinking water over 6, 9, or 12 weeks (Wu et al. 1996). After resection, 0.5 cm3 liver pieces were snap-frozen in liquid nitrogen and stored at − 80C until used. Serial cryostat sections 8–m thick were cut on a motor-driven cryostat (Bright: www.hackerinstruments.com, Huntington, UK). Sections were picked up on clean glass slides and stored at −25C until use. For every series of sections, one was used for hematoxylin–eosin staining for histological analysis and one for PAS staining of glycogen to establish the presence of HCC (Bannasch et al. 1982; Hacker et al. 1982). Normal hepatic tissue and preneoplastic foci contain glycogen, whereas hepatocellular carcinoma does not.
Enzyme Histochemical Localization of G6PD Activity
Incubation media for the demonstration of G6PD activity were prepared as previously described (Van Noorden and Frederiks 1992) and consisted of 100 mM phosphate buffer (pH 7.45) containing 18% (w/v) polyvinyl alcohol (PVA; hot water-soluble, weight average Mr 70,000–100,000; Sigma), 10 mM glucose-6-phosphate (G6P; Boehringer: www.roche.com, Mannheim, Germany), 0.8 mM NADP (Boehringer), 4.5 mM neotetrazolium chloride (NT; Polysciences: www.polysciences.com, Poole, UK) and 0.45 mM mPMS (Serva: www.serva.de, Heidelberg, Germany). Control reactions were performed in the absence of substrate and co-enzyme (Butcher and Van Noorden 1985). Incubation media were poured into glass vials and equilibrated for at least 7.5 min at 37C in an atmosphere of either nitrogen or oxygen, using a tonometer to prevent formation of gas bubbles in the viscous media (Butcher 1978). The flow rate of the gases was 500–800 ml/min. Immediately before incubation, sections were taken from the storage cabinet and air-dried for 5 min at 37C. After placement of plastic rings around the sections, media were poured onto the sections and glass coverslips were placed on top of the rings to keep the incubation media saturated with either nitrogen or oxygen. After 7.5 min of incubation at 37C, the reaction was stopped by rinsing off the viscous PVA-containing media from the sections using hot tapwater. Afterwards, sections were mounted in glycerol jelly.
Involvement of SH Groups and GSH in G6PD Activity
To establish the involvement of SH groups in G6PD activity, 10 mM N-ethylmaleimide (NEM; BDH: www.bdh.com, Poole, UK), a blocker of SH groups, was added to incubation media (Neilands 1954; Van Noorden et al. 1985). For this purpose, the oxygen insensitive tetrazolium salt nitroblue tetrazolium (NBT; Sigma) was applied to detect G6PD activity in the presence of 20% oxygen (air). The effect of SH groups on formazan formation was further investigated by addition of GSH to the media. For this purpose, films of GSH were applied onto clean glass slides. The films were prepared by spreading 10–l of an aqueous solution of 0.1 or 1.0 mM GSH over an area of ∼1 cm2. The slides were air-dried at room temperature (RT). Then tissue sections were mounted on top of the GSH film and stored in the cryostat cabinet (−25C) until use.
Histochemical Localization of GSH
GSH was localized using a solution of 5.0 mM Mercury Orange (Sigma) in toluene. Mercury Orange forms a complex with SH groups that are present in GSH and proteins. When short incubation times are used, GSH is preferentially stained (Asghar et al. 1975). The procedure as described by Chieco and Boor (1983) was used, with minor modifications. We obtained optimal results by drying the sections for 5 min at 37C. After drying, sections were kept briefly in steam from boiling water and, as a result, a thin layer of water vapor condensed on the sections. In this way the thickness of the water layer was controlled, which appeared to be important because we found that a thick layer of water caused artifacts such as the formation of Mercury Orange crystals. Immediately after applying the water vapor, incubation in a solution of Mercury Orange was performed at RT for 45 sec. The solution was rinsed off with toluene. Afterwards, sections were mounted in euparal.
Image Analysis to Quantify Histochemical Reactions
Formazan production in sections was measured by image analysis according to Chieco et al. (1994) using a Vanox-T photomicroscope (Olympus: www.olympus.com, Tokyo, Japan) with a ×2 objective (NA 0.08). Cryostat sections were illuminated with white light from a stabilized power supply after filtering by infrared blocking filters (Jonker et al. 1997) and a monochromatic filter of the isobestic wavelength of NT and NBT formazan (585 nm; Van Noorden and Frederiks 1992). Images of sections were captured using a CCD camera (4913 Cohu: www.cohu.com, San Diego, CA), that was attached to a frame grabber (Scion Image 1.59 for Mac; Scion: www.scioncorp.com, Frederick, MD) and a computer (8100: Apple Macintosh: www.apple.com, Cupertino, CA). Gray values were converted to absorbance values by using a set of neutral density filters (Jonker et al. 1997). Absorbance values were converted into amounts (μmoles) of substrate converted per min per gram wet weight of tissue (Van Noorden and Frederiks 1992). Absorbance values of control reactions were subtracted from test values to obtain specific activity (Butcher and Van Noorden 1985). RA was calculated as percentage of the amount of formazan produced in the presence of oxygen and the amount that was produced in the presence of nitrogen, as measured in serial sections (Griffini et al. 1994).
Biochemical Assays
Spectrophotometric analysis of G6PD activity was performed using 3 ml of a buffered 5% PVA solution containing NADP, G6P, mPMS, and NT in the concentrations as described for the histochemical assay. Solutions were saturated with either nitrogen or oxygen. After saturation, 30 μl of a 1:100 diluted G6PD solution (1000 U/ml, from Leuconostoc mesenteroides; Boehringer) in distilled water was added to start the reaction. Cuvettes were covered with glass slides to ensure that the solutions were kept in a saturated oxygen or nitrogen environment. The involvement of SH groups in the reaction was studied by adding 10 mM NEM or 0.1 or 1.0 mM GSH to the media. Increases in absorbance were measured at 585 nm using an Ultrospec III spectrophotometer (Pharmacia: www.apbiotech.com, Uppsala, Sweden).
Statistical Analysis
Statistical processing of data was performed using Excel 97 (Microsoft: www.microsoft.com, San Jose, CA) and SPSS 8.0 for Windows (SPSS: www.spss.com, Chicago, IL). Differences were considered statistically significant when p > 0.05.<
Results
Oxygen Insensitivity of HCC
In all livers investigated, foci of undifferentiated cells were found in hematoxylin–eosin-stained sections. Occasionally these foci were surrounded by connective tissue. Most foci were positive after PAS staining, indicating that these cells contained glycogen (Figure 1A). This means that these cells were preneoplastic and not neoplastic foci (Bannasch et al. 1982; Hacker et al. 1982). Some foci contained hardly any glycogen (Figure 1A), and therefore these foci were considered to be HCC (Bannasch et al. 1982; Hacker et al. 1982). In both types of foci, G6PD activity was very high (Figure 1B). The number of foci and their size increased with time of treatment.
Formazan formation due to G6PD activity, as demonstrated with the oxygen sensitive tetrazolium salt NT in the presence of nitrogen, was high in foci and low in normal liver tissue (Figure 2A). When the assay was performed in the presence of oxygen, formazan was produced in foci of preneoplastic and neoplastic cells but not in normal cells (Figure 2B). In nitrogen, G6PD activity in foci of (pre)neoplastic cells was fourfold higher than in normal cells. The RA was only 6% (95% CI 1.7–9.9) in normal liver tissue and 60% (95% CI 45.5–74.5) in (pre)neoplasms. These differences were significantly different and demonstrate that G6PD activity was oxygen sensitive in normal liver tissue and partly oxygen insensitive in (pre)neoplastic foci (Table 1).
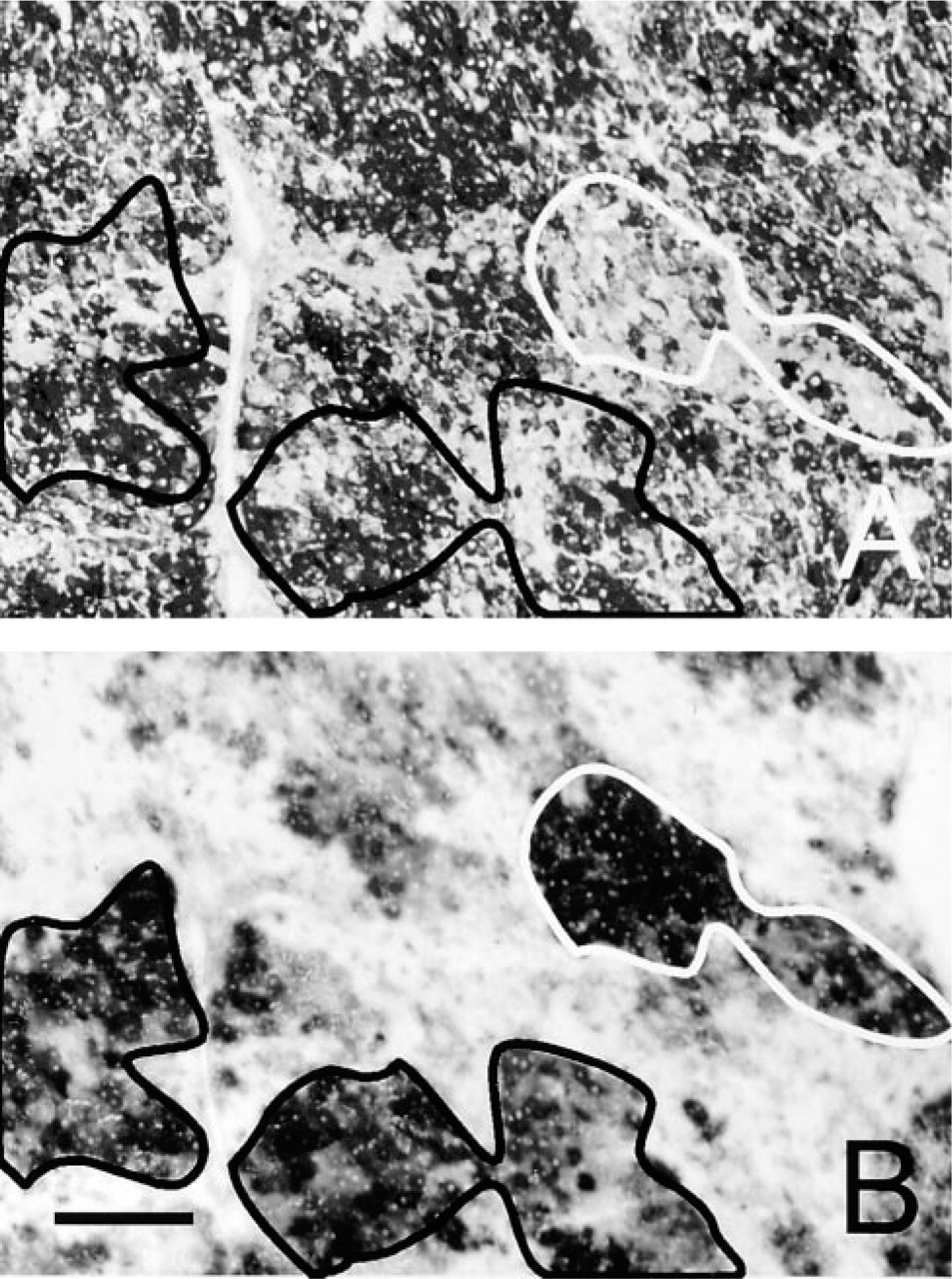
Serial sections of liver of rat treated with diethylnitrosamine for 12 weeks. (
Involvement of SH Groups in G6PD Activity
Incubations in NEM-containing media showed that the activity in normal liver tissue, but not in (pre)neoplastic foci, was sensitive to the blocker of SH groups. Activity in the presence of NEM was half of that in the absence of NEM in normal tissue, whereas in foci, G6PD activity was not affected by NEM (Table 2), indicating that the chemical properties of G6PD are different in normal tissue and (pre)neoplastic foci.
Quantitative histochemical analysis of G6PD activity in normal cells and (pre)neoplastic cells in rat liver in the presence of nitrogen or oxygen a

aG6PD activity is expressed as μmoles G6P converted per g wet weight of tissue per min (± 95% CI) and was calculated on the basis of absorbance of the test minus control reaction for G6PD activity with NT as tetrazolium salt and measured with image analysis. Residual activity (RA) was defined as percentage of the amount of formazan generated in oxygen and that generated in nitrogen. All values of foci and normal tissue were significantly different.
Figure 2C shows intense fluorescence in (preneoplastic tissue but not in normal hepatic tissue due to complex formation of GSH and Mercury Orange. Films made of a solution of 0.1 mM GSH underneath the cryostat sections did not affect the amounts of formazan produced by G6PD activity in control liver tissue and foci in the presence of nitrogen or oxygen, whereas those made of a solution of 1.0 mM GSH abolished most formazan production in all cases (Table 3).
Involvement of SH Groups in G6PD Activity as Determined Biochemically
The involvement of SH groups in the histochemical assay of G6PD activity in the presence and absence of oxygen was investigated further in in vitro experiments. Formation of formazan was followed in cuvettes over time as measured with a spectrophotometer. Activity of purified G6PD was sensitive to oxygen to the same extent as G6PD in normal liver tissue (RA 4%). Moreover, NEM reduced G6PD activity in nitrogen with 86%. This was even stronger than in the histochemical assay applied to normal liver tissue.
Effects of 10 mM NEM on the quantitative histochemical assay of G6PD activity in normal cells and (pre)neoplastic cells in rat liver a

aG6PD activity is expressed as μmoles G6P converted per g wet weight of tissue per min (± 95% CI) and was calculated on the basis of absorbance values of the test minus control reaction for G6PD with NBT as tetrazolium salt and measured with image analysis. Inhibition is defined as percentage of formazan generated by G6PD activity in the presence of NEM (+ NEM) and that in the absence of NEM (–NEM). Inhibition by NEM was statistically significant in normal liver tissue but not in foci.
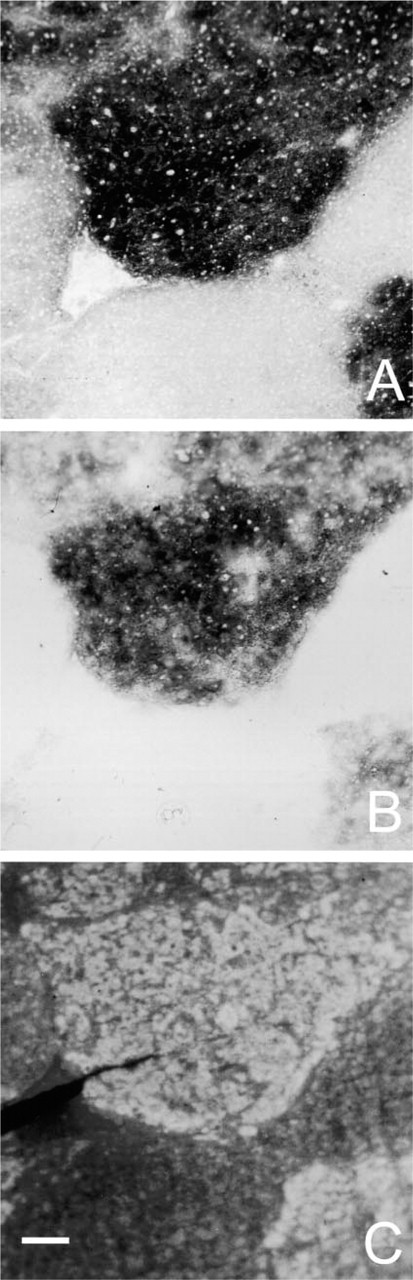
Serial sections of liver of rat treated with diethylnitrosamine for 6 weeks. (
Effects of reduced glutathione (GSH) on the quantitative histochemical assay of G6PD activity in normal cells and (pre)neoplastic cells in rat liver a
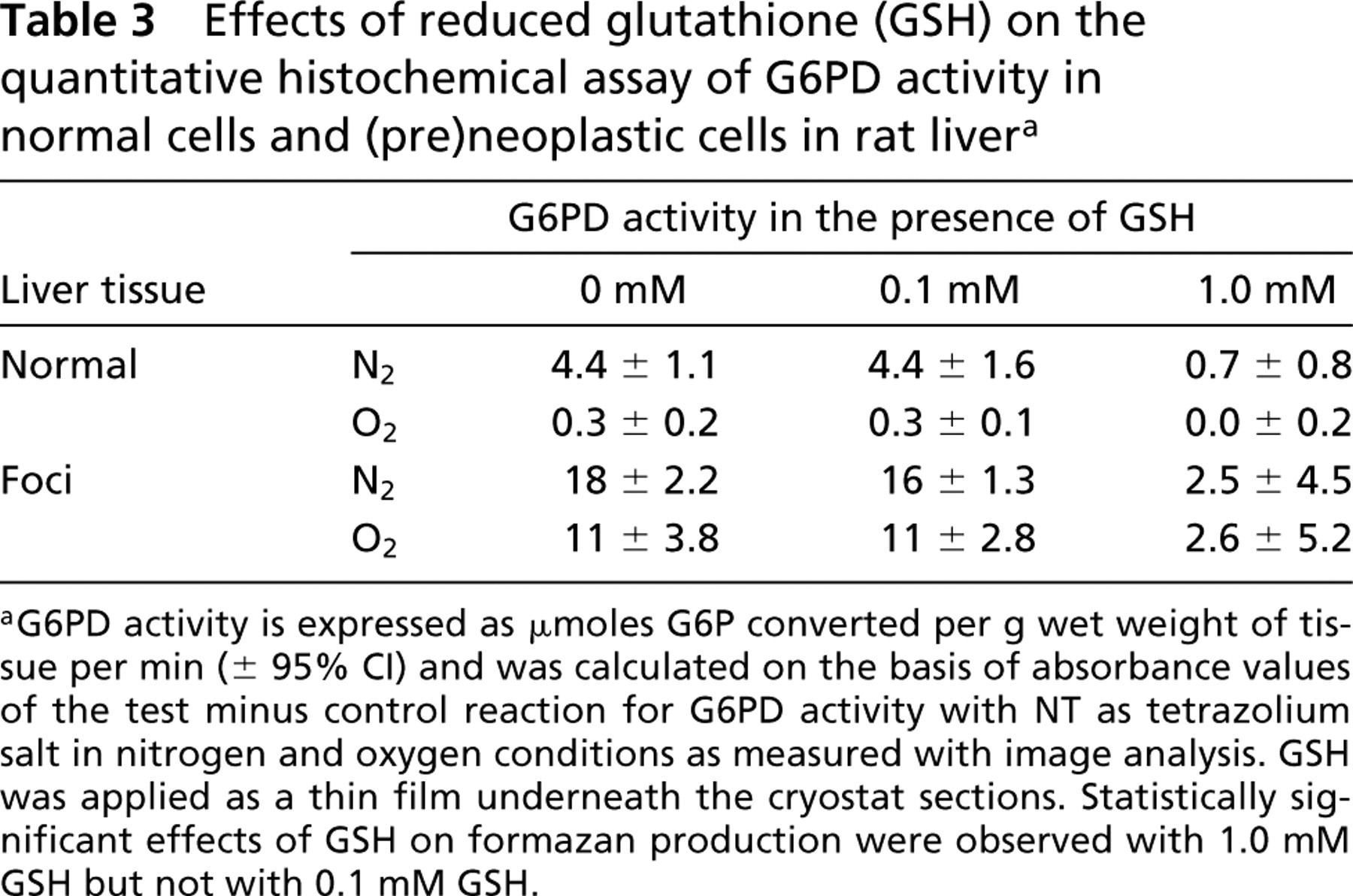
aG6PD activity is expressed as μmoles G6P converted per g wet weight of tissue per min (± 95% CI) and was calculated on the basis of absorbance values of the test minus control reaction for G6PD activity with NT as tetrazolium salt in nitrogen and oxygen conditions as measured with image analysis. GSH was applied as a thin film underneath the cryostat sections. Statistically significant effects of GSH on formazan production were observed with 1.0 mM GSH but not with 0.1 mM GSH.
GSH (1 mM) inhibited formazan production by G6PD activity by 90%. Formazan formation in the presence of oxygen was too low to draw valid conclusions with respect to effects of NEM or GSH.
Discussion
The present study shows that oxygen insensitivity of the histochemical assay for G6PD activity occurs only in (pre)neoplastic cells and not in normal liver tissue in rats (Figures 2A and 2B; Table 1). Therefore, it can be used to detect (pre)neoplasm in rat liver. Oxygen insensitivity was demonstrated previously for cancer cells of human breast, bronchus, stomach, colon, and pancreas (Van Driel and Van Noorden 1999). Chemically induced adenomas and carcinomas of the colon of mice showed that the oxygen insensitivity starts to occur in the adenoma stage (Van Driel et al. 1997). Because the assay is a fast, easy, and cheap diagnostic tool, it should be tested for its diagnostic power in (needle) biopsies for detection of HCC cells in human liver.
The extremely low RA of 6% in control liver is clearly below the limit of 20% that was proposed as borderline between non-malignant tissue (<20%) and malignant tissue (>20%) by Griffini et al. (1994). A value of 60% for the RA in (pre)neoplastic foci is in agreement with values found in adenocarcinomas of colon and pancreatic carcinoma in humans (Van Driel et al. 1997, 1999a, b). These large differences in formazan production between normal liver tissue and (preneoplastic cells make a discrimination very easy, even when only a few altered cells are present in a biopsy.
The present study also shows that foci of (pre)neoplastic cells contain very high G6PD activity (Figure 1B), which was shown previously by others (Hacker et al. 1982; Baba et al. 1989). However, G6PD activity is high not only in (pre)neoplastic cells but also in necrotic areas and inflammatory cells (Elias et al. 1982; Jonges et al. 1993). Therefore, G6PD activity alone cannot be used to diagnose (pre)neoplasm in unknown human liver biopsies.
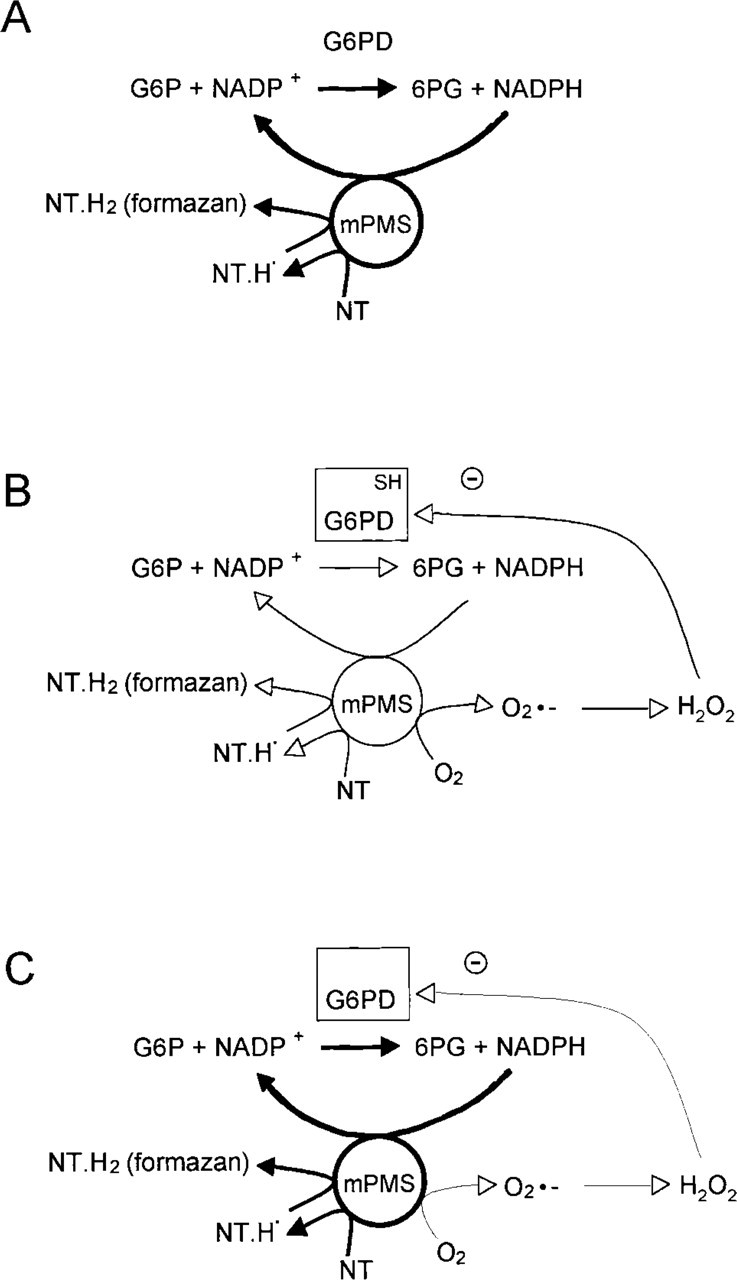
Proposed schemes for the oxygen insensitivity phenomenon in the histochemical assay of G6PD activity using NT in normal cells and (pre)neoplastic cells in nitrogen and oxygen. (
Previous studies indicated that decreased SOD activity and lipid peroxidation capacity, in combination with high G6PD activity in cancer cells, were responsible for the oxygen insensitivity of G6PD in cancer cells (Van Driel and Van Noorden 1999). However, we found that both SOD activity and lipid peroxidation capacity were not reduced in (pre)neoplastic foci (unpublished results). Therefore, we reinvestigated biochemical backgrounds of the oxygen insensitivity phenomenon in this rat model of HCC and focused on the effects of SH groups on G6PD activity in normal and cancer cells.
First, we observed high amounts of GSH in (pre)neoplastic foci in comparison with normal liver tissue (Figure 2C). We hypothesized that GSH plays a role in the oxygen insensitivity phenomenon by prevention of lipid peroxidation due to capture of hydrogen peroxide. It is known that lipid peroxidation products inhibit G6PD activity (Kanazawa and Ashida 1991; Szweda and Stadtman 1992). However, we found that exogenous GSH at high concentrations, but not at low concentrations, prevented and did not induce formazan formation due to G6PD activity, independent of the type of cells or the presence or absence of oxygen (Table 3). Moreover, our in vitro experiments also clearly demonstrated that GSH does not induce oxygen insensitivity and that oxygen sensitivity of G6PD also occurs in the absence of lipid peroxidation. Therefore, GSH and lipid peroxidation products are unlikely to play a role in the discrimination between cancer cells and non-cancer cells on the basis of the oxygen (in)sensitivity phenomenon. It is not clear whether exogenous GSH inhibits G6PD activity directly or prevents formazan formation for other thus far unknown reasons.
We next hypothesized that changes in the G6PD protein render (pre)neoplastic foci oxygen insensitive, and therefore we investigated involvement of SH groups in the activity of G6PD. We found that NEM, as blocker of SH groups, reduced G6PD activity in control liver tissue in situ as well as in vitro. In contrast, G6PD activity in (pre)neoplastic cells was not affected by NEM. This shows that G6PD in normal cells, but not in (pre)neoplastic cells, is dependent on SH groups for its activity. Gel electrophoresis of microdissected tissue should establish whether the properties of the enzyme in normal tissues and (pre)neoplastic foci differ also in other aspects.
The consequences of our results for the chemical backgrounds of the oxygen insensitivity phenomenon are summarized in Figure 3. It is a simplification of the scheme proposed by Van Driel and Van Noorden (1999). Incubations performed for the demonstration of G6PD activity with NT as tetrazolium salt in the absence of oxygen result in proper detection of enzyme activity in normal cells and (pre)neoplastic cells (Figure 3A). However, in the presence of oxygen, some formazan is produced in (pre)neoplastic cells (Figure 3C) but not in normal cells (Figure 3B). The major explanation for differences in oxygen sensitivity of the histochemical assay of G6PD activity in normal cells and (pre)neoplastic cells is the fact that G6PD in (pre)neoplastic cells is less sensitive for oxygen radical species such as H2O2.
We conclude that the oxygen insensitivity of the histochemical assay for G6PD activity is a fast, easy, and cheap, tool to diagnose (pre)neoplastic cells in rat liver. The absence of essential SH groups in G6PD in (pre)neoplastic cells may be held responsible for the oxygen insensitivity of these cells. Our data warrant further investigations to determine whether the assay can be used for detection of (pre)neoplasm in human liver.
Footnotes
Acknowledgements
Acknowledgments
We thank C.G. Wu for donating the animal tissues, Ms K.S. Bosch for technical assistance, Mr J. Peeterse for preparation of the figures, and Ms T.M.S. Pierik for careful checking of the manuscript.