
Abstract
Expression of glucose-6-phosphate dehydrogenase (G6PD) activity is high in tongue epithelium, but its exact function is still unknown. It may be related either to the high proliferation rate of this tissue or to protection against oxidative stress. To elucidate its exact role, we localized quantitatively G6PD activity, protein and mRNA using image analysis in tongue epithelium of rat and rabbit, two species with different diets. Distribution patterns of G6PD activity were largely similar in rat and rabbit but the activities were twofold lower in rabbit. Activity was two to three times higher in upper cell layers of epithelium than in basal cell layers, whereas basal layers, where proliferation takes place, contained twice as much G6PD protein and 40% more mRNA than upper layers. Our findings show that G6PD is synthetized mainly in basal cell layers of tongue epithelium and that it is posttranslationally activated when cells move to upper layers. Therefore, we conclude that the major function of G6PD activity in tongue epithelium is the formation of NADPH for protection against oxidative stress and that diet affects enzyme expression in this tissue.
Keywords
G
Materials and Methods
Tissue
Six male Wistar rats, weighing 200–250 g, and six mature male New Zealand White rabbits, with a mean weight of 5 kg, were used. Animals were sacrificed with an overdose of barbiturates. The tongue was immediately removed and divided into three parts, the anterior, central, and posterior parts. Small fragments of all parts with a thickness of up to 5 mm were frozen in liquid nitrogen and stored at −80C. Cryostat sections 8 μm thick were cut at −25C on a motor-driven Bright cryostat fitted with a rotary retracting microtome, picked up on clean glass slides, and stored at −30C.
Histochemical Localization of G6PD Activity
Cryostat sections were allowed to dry at room temperature (RT) for 5 min and were then incubated for the demonstration of G6PD activity according to Van Noorden and Frederiks (1992). Incubation medium contained 18% polyvinyl alcohol (PVA, average Mr 70,000–100,000; Sigma, St Louis, MO) in 0.1 M phosphate buffer (pH 7.4), 10 mM glucose-6-phosphate (Boehringer; Mannheim, Germany), 0.8 mM NADP+ (Boehringer), 0.4 mM 1-methoxyphenazine methosulfate (Serva; Heidelberg, Germany), 5 mM MgCl2, 5 mM sodium azide, and 5 mM nitro BT (Sigma). The medium was freshly prepared just before incubation and nitro BT was added after being dissolved in a heated mixture of dimethylformamide and ethanol (final dilution of each solvent in the medium was 2% v/v). Sections were incubated for 2 min at RT and then rinsed with hot tapwater to remove the incubation medium and to stop the reaction immediately, and were mounted in glycerin–gelatin. Control reactions were performed in the absence of substrate and co-enzyme (Butcher and Van Noorden 1985). To establish the contribution of SH groups in cells to nonspecific formation of final reaction product, 10 mM N-ethylmaleimide (Sigma) was added to control incubation medium (Van Noorden et al. 1985). The possible involvement of conversion of endogenous substrate by G6PD in the control reaction was determined by incubation in the presence of 10 mM epiandrosterone, a specific inhibitor of G6PD (Shantz et al. 1989).
Immunohistochemistry of G6PD Protein
Rat G6PD protein was demonstrated in rat tongue using a polyclonal rabbit anti-yeast G6PD antibody (Sigma; dilution 1:200) according to Straatsburg and Frederiks (1997). Cryostat sections were air-dried (1 hr or overnight), fixed in cold acetone (10 min, −20C), and dried again (10 min). Antigens were exposed for 30 min by using a solution of Tris (10 mM), EDTA (5 mM), NaCl (150 mM), 0.25% gelatin, and 0.5% Tween-20 (Sigma) at pH 8.0. Between all further incubation steps, sections were thoroughly rinsed in PBS. The antibody was diluted in PBS containing 0.2% (w/v) bovine serum albumin (BSA) and 1% (v/v) normal rat serum to block nonspecific staining. Sections were incubated for 60 min at RT. After rinsing, sections were incubated for 90 min at RT with horseradish peroxidase-labeled goat anti-rabbit IgG (Dako, Glostrup, Denmark; dilution 1:100). Peroxidase activity was detected by incubating the sections for 10 min in a medium containing 0.5 mg/ml diaminobenzidine (Fluka, Buchs, Switzerland), 3 mM H2O2, and 50 mM Tris-HCl buffer (pH 7.6).
Rabbit G6PD protein was demonstrated using a polyclonal goat anti-rabbit G6PD antibody (dilution 1:50) (Ninfali et al. 1990). IgGs were purified from goat serum by DEAE Affi-Gel Blue chromatography (Bio-Rad; Milan, Italy) as reported elsewhere (Ninfali et al. 1994). Cryostat sections were air-dried (1 hr or overnight), fixed in cold acetone (10 min, −20C), and dried again (10 min). Between all further incubation steps, sections were thoroughly rinsed in PBS. To block nonspecific staining, sections were incubated with PBS containing 3% BSA; antibodies were diluted in PBS containing 0.3% (w/v) BSA and 5% (v/v) normal rabbit serum. Sections were incubated with the first antibody for 60 min at RT and then with horseradish peroxidase-labeled rabbit anti-goat IgG (Dako; dilution 1:100). Peroxidase activity was detected by incubating the sections for 10 min in a medium containing 0.5 mg/ml diaminobenzidine, 3 mM H2O2, and 50 mM Tris-HCl buffer (pH 7.6).
In Situ Hybridization of G6PD mRNA
The procedure to localize G6PD mRNA was performed as described by Baumgart et al. (1997). Cryostat sections of rat tongue were air-dried and then fixed with 4% paraformaldehyde in PBS for 20 min at RT. After thorough rinsing, sections were treated with 10 μg proteinase K (Sigma) per ml PBS for 15 min at RT. After rinsing and acetylation, sections were dried thoroughly and incubated overnight with double digoxygenin-labeled RNA probes (Amersham Pharmacia Biotech; Roosendaal, The Netherlands) in hybridization buffer. Two probes were used. The G6PD1 mRNA probe was the antisense sequence of the one that codes for the amino acid sequence tgttgtgctgctgggtcacccggatgaatg and the G6PD3 mRNA probe for catcgagccagacgctgctttgtccccagg. These sequences were derived from the human genome as described by Kanno et al. (1989). As control, sections were incubated with 10 μg/ml RNase for 30 min at RT before hybridization. After rinsing, sections were incubated with anti-digoxygenin–Fab conjugated with alkaline phosphatase (Boehringer; diluted 1:400) for 3 hr at RT. Alkaline phosphatase activity was detected by incubating in 0.1 M Tris HCl buffer, pH 9.0, containing 0.4 mM nitro BT, 0.4 mM 5-bromo-4-chloro-3-indolyl phosphate, and 10 mM levamisole for 10 min at 37C (Van Noorden and Frederiks 1992).
Immunohistochemistry of Proliferating Cell Nuclear Antigen (PCNA)
PCNA was detected immunohistochemically in cryostat sections of rat tongue. They were air-dried (1 hr or overnight) and fixed for 2 min in 4% formaldehyde in phosphate buffer, pH 7.4. After dehydration and rehydration in a series of ethanol (50%–70%–90%–96%–90%–70%–50%), sections were rinsed in PBS and then incubated with PBS containing 10% normal goat serum. Between all further incubation steps, sections were thoroughly rinsed in PBS. Sections were incubated with the mouse monoclonal antibody PC10 against PCNA (Dako; dilution 1:100) for 60 min and then incubated with biotinylated rabbit anti-mouse Ig (Dako; dilution 1:200) in the presence of 10% normal rat serum. Sections were incubated with a streptavidin–biotin–horseradish peroxidase complex (Dako; dilution 1:100) prepared 30 min in advance and mixed shortly before use with an equal volume of PBS and 20% normal rat serum. Peroxidase activity was detected by incubating the sections for 10 min in a medium containing 0.5 mg/ml diaminobenzidine, 3 mM H2O2, and 50 mM Tris-HCl buffer (pH 7.6).
Image Analysis and Processing
Serial sections of anterior, central and posterior parts of tongue from six rats and six rabbits were used to quantify G6PD activity, protein, and mRNA with image analysis. In each section (one section per part of the tongue per animal), three areas were selected in the basal cell layers and three in the upper cell layers of the epithelium. In each selected area, three measurements were made and mean gray values were calculated. Gray values were converted into absorbance values by using a set of neutral density filters (Kodak, Rochester, NY) (Jonker et al. 1997). Values of control reactions were subtracted from test values to obtain specific enzyme activity (Butcher and Van Noorden 1985; Geerts et al 1996). A Vanox-T photomicroscope (Olympus; Tokyo, Japan) with a ×2 objective (NA 0.08) was used. Cryostat sections were illuminated with white light from a stabilized power supply after filtering with infrared blocking filters (Chieco et al. 1994; Jonker et al. 1997) and a monochromatic filter of the isobestic wavelength of NBT–formazan (585 nm; Van Noorden and Frederiks 1992) was used for activity measurements. Amounts of protein and mRNA in basal and upper cell layers of the epithelium were determined in an identical way as activity except for the use of a monochromatic filter at a wavelength of 460 nm for the protein (Geerts et al. 1996). Images of the sections were captured by using a Cohu 4913 CCD camera (Cohu; San Diego, CA) and an 8-bit monochrome frame grabber (LG-3; Scion, Frederick, MD). Data analysis of captured images was performed on a Power Macintosh 8100/110 computer (Apple; Cupertino, CA) using the public domain NIH Imaging software program (written by Wayne Rasband; United States National Institutes of Health) and available via Internet from http://rsb.info.nih.gov/.
Statistics
Results were tested statistically for differences using the Student's t-test, and p=0.05 was taken as level of significance.
Results
Localization of G6PD Activity
G6PD activity was not significantly different in the anterior, central, and posterior parts of tongue epithelium of rat and rabbit (not shown). Figure 1A shows the localization pattern of G6PD activity in the median sulcus of rat tongue. The epithelium in this region is mainly organized as filiform papillae, which consist of stratified squamous epithelium including a basal germinative layer and cornified projections. Among the filiform papillae, scattered fungiform papillae were present, with one taste pore on top covered by a slightly keratinized layer. G6PD activity in epithelium was very high in comparison with that in other tissue components of the tongue such as muscle, connective tissue, and nerves (Figure 1A). Activity was hardly present in the connective core of papillae.
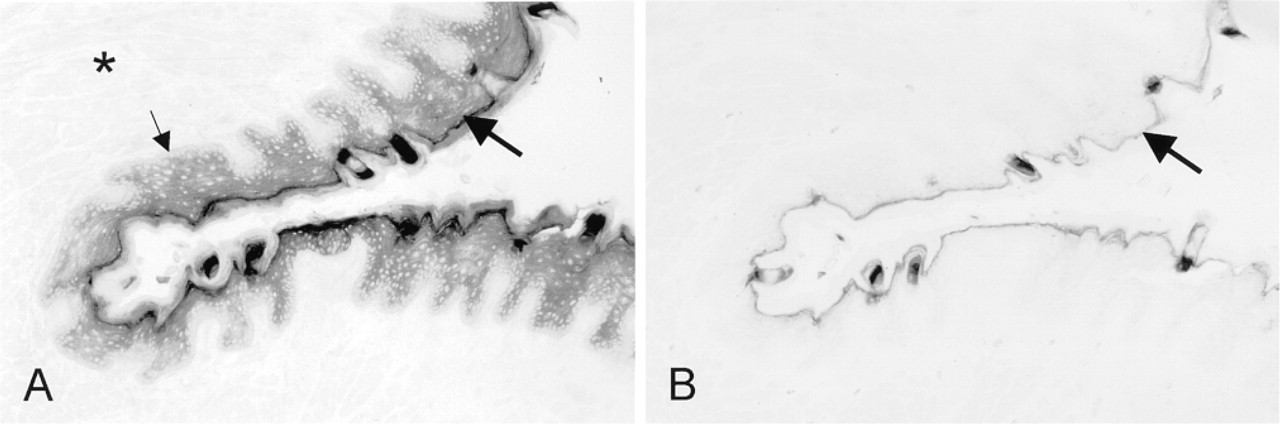
Distribution patterns of G6PD activity after incubation in the presence (test reaction;
The following distribution patterns of G6PD activity were observed in epithelium. Basal cell layers contained low activity, whereas activity became higher towards upper cell layers (Figure 1A and 2A). Activity was not detectable in nuclei of epithelial cells. Control incubations did not result in any significant precipitate in these layers (Figure 1B). Highest amounts of final reaction product were found in keratinized superficial layers (Figure 1A), but high amounts were also found after the control reaction and can therefore be considered to represent nonspecific staining (Figure 1B). Formation of final reaction product in the keratinized layer was prevented by incubating sections in the presence of 10 mM N-ethylmaleimide. This finding suggests that SH groups in keratin were responsible for nonspecific staining. Addition of 10 mM epiandrosterone, a specific inhibitor of G6PD, to the incubation medium containing substrate and co-enzyme resulted in similar staining patterns and intensities as those in the control reactions, which confirms that SH groups in keratin caused nonspecific staining in superficial layers but that staining was specific in basal and upper cell layers.
Similar G6PD activity distribution patterns were observed in tongue of rabbit. The overall activity was lower than that found in rat but the distribution pattern was similar, with the basal cell layers containing lower activity than the upper cell layers.
Image analysis revealed that rat tongue epithelium contained twofold higher activity than that of rabbit, whereas activity in the upper epithelial cell layers was almost threefold higher than that in basal cell layers in both rat and rabbit (Table 1).
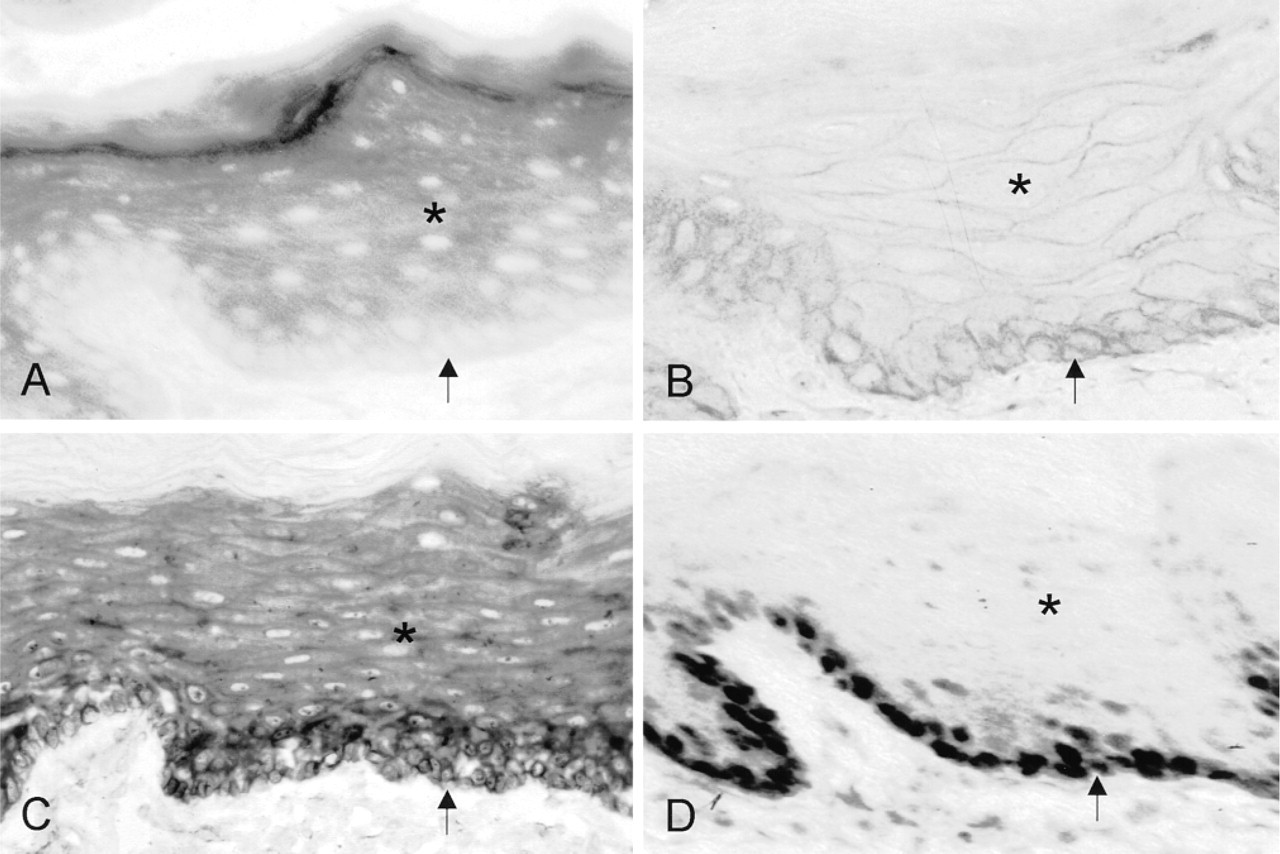
Distribution patterns of G6PD activity (
Immunohistochemical Localization of G6PD Protein
G6PD protein distribution patterns in epithelium of rat tongue are shown in Figure 2B. Highest amounts were observed in basal cell layers and lower levels in the upper cell layers. Protein was not present in keratinized layers. Nuclei of epithelial cells were negative (Figure 2B). Control incubations resulted in the absence of staining except for the external (luminal) surface of tongue epithelium, which must be due to food residues because G6PD activity was not present here (Figure 1A). Table 1 shows that the amounts of protein in basal cell layers of both species were at least twofold higher than those in the upper cell layers.
In Situ Hybridization of G6PD mRNA
Figure 2C shows the localization of G6PD mRNA in the anterior part of rat tongue. Similar results were obtained with both mRNA probes, G6PD1 and G6PD3. The distribution patterns of G6PD mRNA showed that mRNA levels were highest in basal cell layers of the epithelium and decreased towards upper cell layers. In general, mRNA levels were about 40% higher in basal cell layers than in upper cell layers (Table 1). mRNA was absent in the keratinized superficial layers. RNase treatment completely abolished staining.
Immunohistochemical Localization of PCNA
Proliferating cells were localized immunohistochemically by detection of PCNA in rat tongue. Figure 2D shows that nuclei positive for PCNA were present only in basal epithelial cell layers. Control incubations resulted in the absence of staining.
Discussion
In this study, we have shown that G6PD activity, protein, and mRNA have distinct distribution patterns in tongue epithelium of rat and rabbit. The activity of the enzyme is extremely high in tongue epithelium. For example, G6PD activity is here 50-fold higher than in rat liver (Kanno et al. 1989).
G6PD activity, protein, and mRNA in basal and upper cell layers of tongue epithelium expressed as absorbance of test minus controla
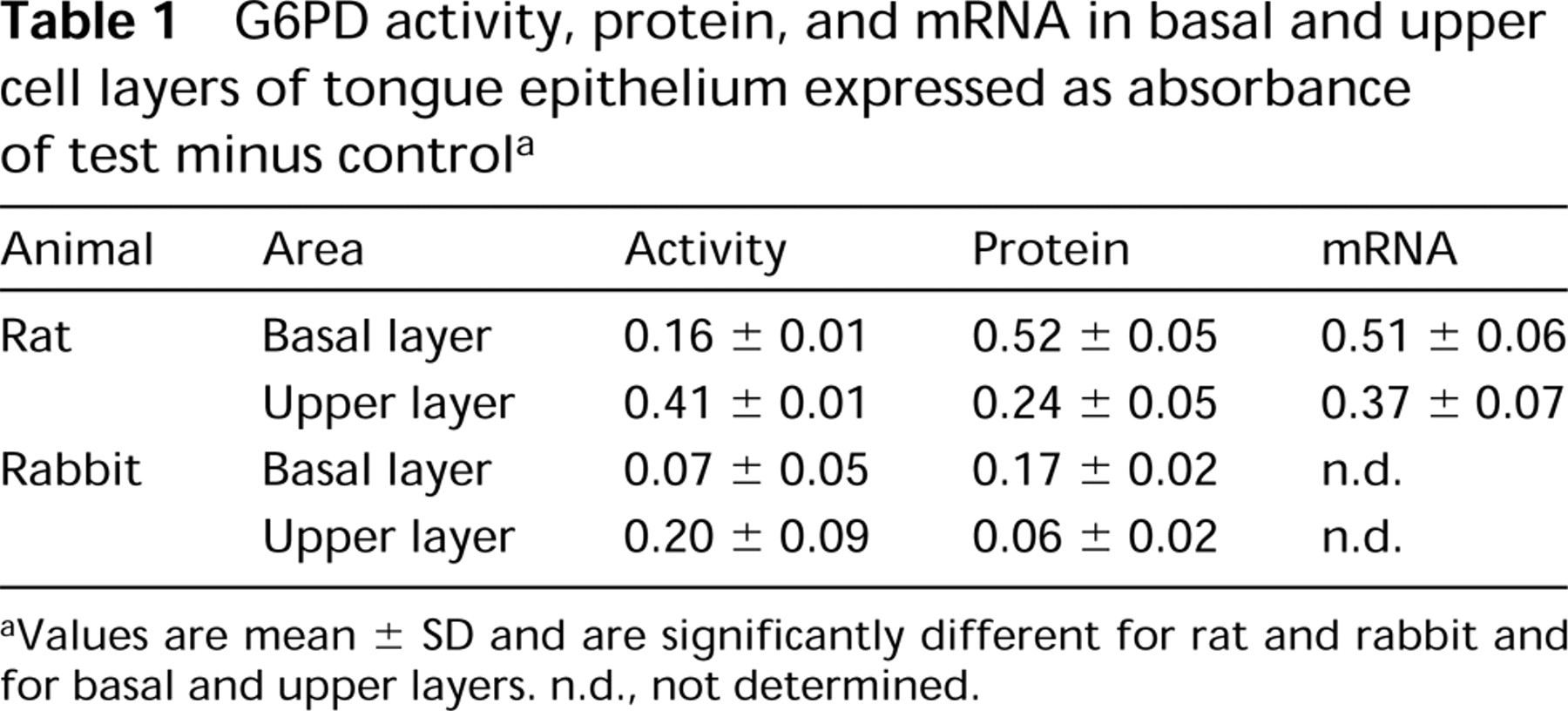
aValues are mean ± SD and are significantly different for rat and rabbit and for basal and upper layers. n.d., not determined.
The histochemical reaction to demonstrate G6PD activity is specific. The specific reaction is defined as the test minus control reaction (Butcher and Van Noorden 1985), and final reaction product generated in this specific reaction was completely abolished by addition of 10 mM epiandrosterone, a specific inhibitor of G6PD. This means that 6-phosphogluconate dehydrogenase, a following enzyme in the hexose monophosphate shunt that converts the product of G6PD with concomitant production of NADPH, was not involved in production of staining. This has been shown previously in rat liver (Jonges and Van Noorden 1989). The high amounts of final reaction product generated in the test and control reactions in cornified projections of filiform papillae were not affected by epiandrosterone but were largely abolished by NEM, a blocker of SH groups (Van Noorden and Frederiks 1992), suggesting that staining in this region was due to SH groups in proteins and not to G6PD activity using endogenous substrates. Keratin is the most likely candidate, because soft and hard keratins contain large numbers of SH groups both in rat and rabbit tongue epithelium (Farbman 1970).
Image analysis showed that G6PD activity in tongue epithelium is twofold higher in rat than in rabbit. Both animals showed a non-homogeneous distribution of G6PD activity over the epithelial layers. Highest activity was found in the upper cell layers of the epithelium, whereas the basal cell layers showed lower activity. The ratios of the activities in the upper and basal cell layers were similar for both animals. The different activities in the two species may be explained by their diets: rats are omnivorous, whereas rabbits are herbivorous. Previous studies have shown that differences in feeding habits, mastication, and mechanical stress can modify characteristics of epithelial cells of the tongue (Kullaa–Mikkonen et al. 1987).
The study of distribution patterns of G6PD protein in serial sections was performed to correlate amounts of protein and activity. Protein content was highest in cells of the basal layers in both rat and rabbit. This means that areas containing relatively high amounts of protein express low activity, whereas the areas with relatively low amounts of protein show high activity. Because G6PD mRNA showed similar distribution patterns as the protein, we conclude that the enzyme is mainly synthesized in cells of the basal layers, and becomes activated posttranslationally in upper cell layers. There are indications that the enzyme is synthetized in a glycosylated form, which is less active, and that the enzyme becomes activated by removal of oligosaccharide groups (Ozols 1993). In accordance with this, the study of Ursini et al. (1997) suggests that G6PD molecules are synthesized in inactive form and can be activated according to cellular needs. This may well be due to binding to structural elements, as has been suggested by Swezey and Epel (1986) and by Stanton et al. (1991). However, recent studies have demonstrated that other factors also affect G6PD expression and activation. Oxidative stress can induce increased G6PD mRNA levels in human hepatoma cells, indicating regulation at the transcriptional level (Ursini et al. 1997), whereas small heat shock proteins are responsible for increased G6PD activity in human and murine hepatoma cell lines indicating posttranscriptional regulation (Preville et al. 1999).
PCNA localization revealed that the basal cell layers of tongue epithelium are the only cell layers that contain proliferating cells. This means that, in these cells, low levels of G6PD activity sustain proliferation, as was shown in previous studies (Ninfali et al. 1987, 1991; Tian et al. 1998). However, the main role of G6PD in the upper cell layers of tongue epithelium may well be generation of NADPH needed for the defense against free radicals generated by food intake (Kletzien et al. 1994).
In summary, our findings suggest that in tongue epithelium of rat and rabbit, G6PD is mainly synthesized in the basal cell layers, where it is not very active. It is activated in upper cell layers, indicating that its main function in tongue epithelium is NADPH production for protection against oxidative stress. Our quantitative localization study of G6PD activity, protein, and mRNA clearly shows the functional importance of posttranslational regulation of activity of G6PD.
Footnotes
Acknowledgements
Acknowledgments
We wish to thank Dr Frank van den Berg and Dr Eleonora Aronica for their help in performing the in situ hybridization of G6PD, Mr Jan Peeterse for photographic contributions, and Ms Trees Pierik for preparing the manuscript.