
Abstract
Keywords
T
We report here a simple method for detection and localization of transglutaminase activities in tissue homogenates and histological sections. The method uses, as a substrate, a derivative of green fluorescent protein (GFP), His6-Xpress-GFP, which can be easily obtained by recombinant DNA technology, and monitors its size changes or covalent anchoring to tissue sections. On histological examination, a surprisingly wide variety of tissues gave strong positive signals, especially in their extracellular matrices, suggesting an important role of transglutaminase in the maintenance of the integrity of the extracellular matrix (ECM).
Materials and Methods
Materials
Guinea pigs (male, 8 weeks old) were obtained from Tokyo Laboratory Animal Science (Tokyo, Japan). Guinea pig liver transglutaminase (TGase) and restriction enzymes were purchased from Takara (Kyoto, Japan). pGFPuv vector and rGFPuv were from Clontech (Palo Alto, CA). Ni-NTA agarose resin was from Qiagen, (Chatsworth, CA). pTrcHis A was from Invitrogen (San Diego, CA). Lysyl endopeptidase was from Wako (Tokyo, Japan). μ Bondasphere 5 μ C18–100 Å 3.9 × 150 mm was from Waters (Milford, MA). Tissue-Tek OCT compound was from Miles (Elkhart, IN). Immobilon PSQ membrane was from Millipore (Bedford, MA). 5-(biotinamido)pentylamine was from Pierce (Rockford, IL). Vectabond and horseradish peroxidase-avidin D were from Vector Laboratories (Burlingame, CA). Hydrogenated Triton X-100 and 2,2′-azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) diammonium salt (ABTS) were from Sigma (St Louis, MO). Blood coagulation factor XIIIa (FXIIIa) was a gift from Dr. Yuji Saito (Ueki et al. 1996).
Construction of His6-Xpress-GFP Expression Vector
A 0.75-kb PstI-EcoRI fragment of pGFPuv, which encodes the entire open reading frame of GFPuv (abbreviated here simply as GFP), was inserted into the PstI-EcoRI cloning sites of pTrcHis A vector and named pHis6-Xpress-GFP.
Expression and Purification of His6-Xpress-GFP
Escherichia coli JM109 was transformed with pHis6-Xpress-GFP and grown in 2 × Luria Bertani (LB) medium containing 50 μg/ml ampicillin at 37C with aeration until the cells reached an optical density of 0.6 at 600 nm. The expression of His6-Xpress-GFP was induced by addition of 1 mM isopropyl-β-
Assay by SDS-PAGE of Transglutaminase Activity Using His6-Xpress-GFP as a Substrate
One microgram of tissue transglutaminase was incubated with 0.1 μg of His6-Xpress-GFP or rGFPuv for 1 hr in 20 μl of transglutaminase assay buffer (50 mM Tris-HCl, pH 7.5, 1 mM dithiothreitol, and 5 mM CaCl2) in the presence and absence of 50 mM EDTA or 10 mM putrescine, a transglutaminase inhibitor. The reaction was stopped by adding 5 μl of 5 × Laemlli sample buffer and samples were then separated by SDS-PAGE on a 12.5% gel. After electrophoresis, fluorescent bands in the gel were directly visualized with a Bio-image Analyzer FLA-2000 (Fuji Film; Tokyo, Japan).
Normalization of Activities of Tissue Transglutaminase and FXIIIa and Determination of Their Affinities for His6-Xpress-GFP
For comparison of the affinities of tissue transglutaminase and FXIIIa for the substrate His6-Xpress-GFP, the enzyme levels should be adjusted using a combination of control substrates. This was done by a modified method of Slaughter et al. (1992) using fibrinogen and 5-(biotinamido)pentylamine as substrates. The standard solution of tissue transglutaminase contained 0.31 mg/ml in 10 mM Tris-HCl, pH 7.5, 1 mM EDTA, 1 mM dithiothreitol, and 50% glycerol. Wells of 96-well microtiter plates were coated with 100 μl of 20 μg/μl fibrinogen that serves as a substrate (acyl donor) and were stored at room temperature (RT) for 2 hr. The wells were blocked with 0.5% bovine serum albumin (BSA) in Tris-buffered saline (TBS: 10 mM Tris-HCl, pH 7.4, containing 0.15 M NaCl) for 1 hr and washed twice with TBS. Transglutaminase reaction was initiated by adding 5 mM CaCl2, 10 mM dithiothreitol, 0.5 mM 5-(biotinamido)pentylamine, which serves as acyl acceptor, and a dilution series of tissue transglutaminase or FXIIIa. After a 30-min incubation at 37C, the reaction was stopped by 100 mM EDTA. The plate was washed with 0.01% Triton X-100 in TBS, followed by two washes with TBS. The horseradish peroxidase–avidin D was diluted 1:3000 with 0.5% BSA in TBS, added to the wells, and then incubated at RT for 1 hr. The plate was washed once with 0.01% Triton X-100 in TBS, followed by four washes with TBS, and once with C-P buffer (22.75 mM citric acid and 54.5 mM Na2HPO4, pH 5.3), and 100 μl of peroxidase substrate (50 μl of 0.3% H2O2 in MilliQ water and 50 μl of 2 mg/ml ABTS in C-P buffer) was added to each well. Tissue transglutaminase and FXIIIa activities were determined by measuring absorbance at 405 nm (A405). The enzyme activity that catalyzes an increase of A405 = 0.81 for 10 min at 37C was defined as 100 units. On the basis of this normalization, the substrate specificities of the enzymes against His6-Xpress-GFP were determined according to the above-mentioned SDS-PAGE fluorometric method, using the same units of the enzymes.
Isolation, Digestion, and Amino Acid Sequencing of Crosslinked His6-Xpress-GFP
Five hundred pmol of His6-Xpress-GFP and 1 μg of transglutaminase were incubated in 200 μl of transglutaminase assay buffer at 37C for 1 hr. The reaction was stopped by adding 50 μl of 5 × SDS sample buffer. The sample was heat-denatured at 100C for 2 min, electrophoresed in 12.5% SDS-polyacrylamide gels, and then electrotransferred onto an Immobilon-PSQ membrane. After transfer, the membrane was stained with Coomassie Brilliant Blue. The crosslinked band that was seen in the presence of transglutaminase was excised and sequenced either directly for the N-terminal sequence or after digestion with lysyl endopeptidase in 100 mM Tris-HCl, pH 9.5, 10% acetonitrile, and 1% hydrogenated Triton X-100 for C-terminal analysis. The digestion was performed for 24 hr at 37C. Peptide fragments released were purified by reverse-phase HPLC on a Waters C18 column (3.9 × 150 mm) using a 0–60% gradient of acetonitrile in aqueous trifluoroacetic acid (0.8%). Amino acid sequencing was performed with a peptide sequencer (Shimazu PSQ-1100, Kyoto, Japan) using approximately 100 pmol of peptides.
Activity Staining of Transglutaminase in Tissue Sections with His6-Xpress-GFP
Guinea pigs (n = 10) were sacrificed by guillotine or lethal IP injection of sodium pentobarbital. Tissues were immediately removed, frozen in Tissue-Tek OCT compound on dry ice, and stored at –80C. Frozen sections (7 μm) were cut in a cryostat at –16C, mounted on gelatin-coated slides, and air-dried at RT for 30 min. The use of Vectabond for coating slides sometimes gave nonspecific staining (bright dots). Sections were incubated with His6-Xpress-GFP by overlaying 50–200 μl of its solution (0.1 mg/ml) in transglutaminase assay buffer (50 mM Tris-HCl, pH 7.5, 1 mM dithiothreitol, and 5 mM CaCl2) with or without transglutaminase inhibitors (e.g., 50 mM EDTA or 10 mM putrescine) at RT for 1 hr in a moist chamber. The sections were washed three times in excess PBS and covered with coverslips. The samples were analyzed using either a normal fluorescent microscope with mercury lamp and filters for fluorescein isothiocyanate (FITC) fluorescence (Figures 5A–5A) or a laser scanning confocal microscope (TCS4D, Leica Laser Teknik) based on a Zeiss Axiovert microscope (Carl Zeiss; Oberkochen, Germany) interfaced with a mixed gas argon–krypton laser. Fluorescence acquisitions were performed with the 488-nm laser line to excite GFP (Figures 5D–5D). The fluorescence was very stable. The stained specimens could be viewed repeatedly for at least 3 weeks if stored in a humidified chamber at 4C.
Results
Construction and Purification of GFP Derivatives
The E. coli expression vector pTrcHis used in this study is designed for high-level expression of His-tagged recombinant proteins. The proteins produced have a polyhistidine (6 × His) tag on the N-terminus for rapid purification, which is followed by the Xpress epitope and the enterokinase cleavage site for removal of the fused tags. We noted the presence of two Gln and one Lys in the short stretch of the Xpress epitope and enterokinase cleavage site (Figure 1) and considered the possibility that if GFP is produced as a His6-Xpress-GFP fusion protein using the pTrcHis vector, it may serve as a useful fluorescent substrate of transglutaminase. We therefore produced the His6-Xpress-GFP fusion protein in E. coli and purified it by Ni-chelate chromatography as described in Materials and Methods. The yield was about 3 mg per liter of bacterial culture. The purified protein gave a single fluorescent band of 41 kD on SDS-PAGE (Figure 2, Lanes 4 and 10). Like the wild-type GFP, the N-terminally extended GFP was resistant to the reducing agent 2-mercaptoethanol but sensitive to heat denaturation in the presence of SDS (Figure 2, lane 11). The heat-denatured nonfluorescent His6-Xpress-GFP migrated as a band of 34 kD, a value close to the calculated molecular weight (32,776).

Nucleotide and amino acid sequences of His6-Xpress-GFP. The nucleotide sequence derived from the pTrcHis vector is denoted by capital letters and that from pGFPuv is indicated by lower-case letters. The pTrcHis vector encodes the N-terminal extension: histidine (His) tag and Xpress sequence. Gray boxes indicate partial N-terminal sequences contained in the N-terminal and C-terminal fragments generated from His6-Xpress-GFP after its incubation with transglutaminase (Figure 2, Lane 3; lower band of 36 kD) and digestion with lysyl endopeptidase. Glutamine residues (Q) in the N-terminal extension (Xpress) that can serve as acyl donors are boxed and lysine residues (K) that can act as acyl acceptors are indicated by white on black background.
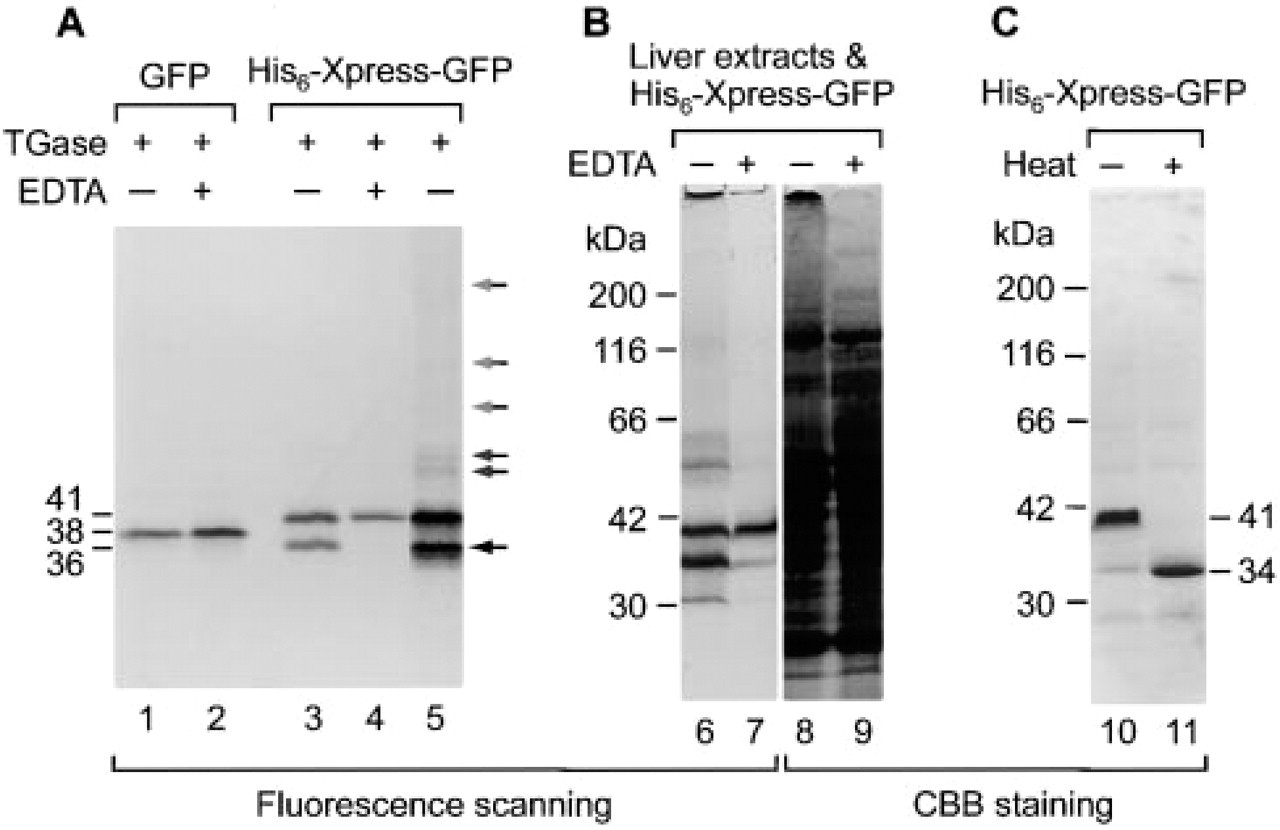
Detection of transglutaminase activity using a fluorescent substrate, His6-Xpress-GFP. (
Electrophoretic Mobility Shift of Crosslinked GFP Derivatives
When His6-Xpress-GFP was incubated with transglutaminase and analyzed by SDS-PAGE, significant changes were observed in its electrophoretic mobility (Figure 2, Lane 3). The wild-type GFP showed no such changes (Figure 2, Lane 1). EDTA, an inhibitor of transglutaminase, completely inhibited the mobility changes (Figure 2, Lane 4). Similar inhibitory effects were also seen with putrescine, a competitive inhibitor of transglutaminase (data not shown). Depending on the incubation time, the transglutaminase-treated His6-Xpress-GFP behaved as compact molecules at the initial stage (Figure 2, Lane 3, lower band of 36 kD) and as much larger molecules at later stages of the treatment or at higher concentrations of the substrate (Figure 2, Lane 5). Figure 3 shows the time course of the crosslinking reaction monitored by measuring the fluorescence intensity of the 36-kD band (open circle). It appears unlikely that the lower molecular weight species are proteolytic degradation products because (a) the transglutaminase used is a highly purified preparation, (b) the N-terminal amino acid sequence of the lower molecular weight species (RGSHHHHHH; Figure 1, shading) matched perfectly to that of His6-Xpress-GFP, and (c) amino acid sequencing of lysyl endopeptidase-generated peptide fragments of the lower molecular weight species revealed the presence of the C-terminal fragment beginning with the sequence RDHMVLLEFVTA (Figure 1, shading). The initial lowering of the apparent molecular size (from 41 kD to 36 kD) and the later appearance of the higher molecular weight species (>50 kD) may therefore be due to intramolecular and intermolecular crosslinking, respectively, of His6-Xpress-GFP by transglutaminase.
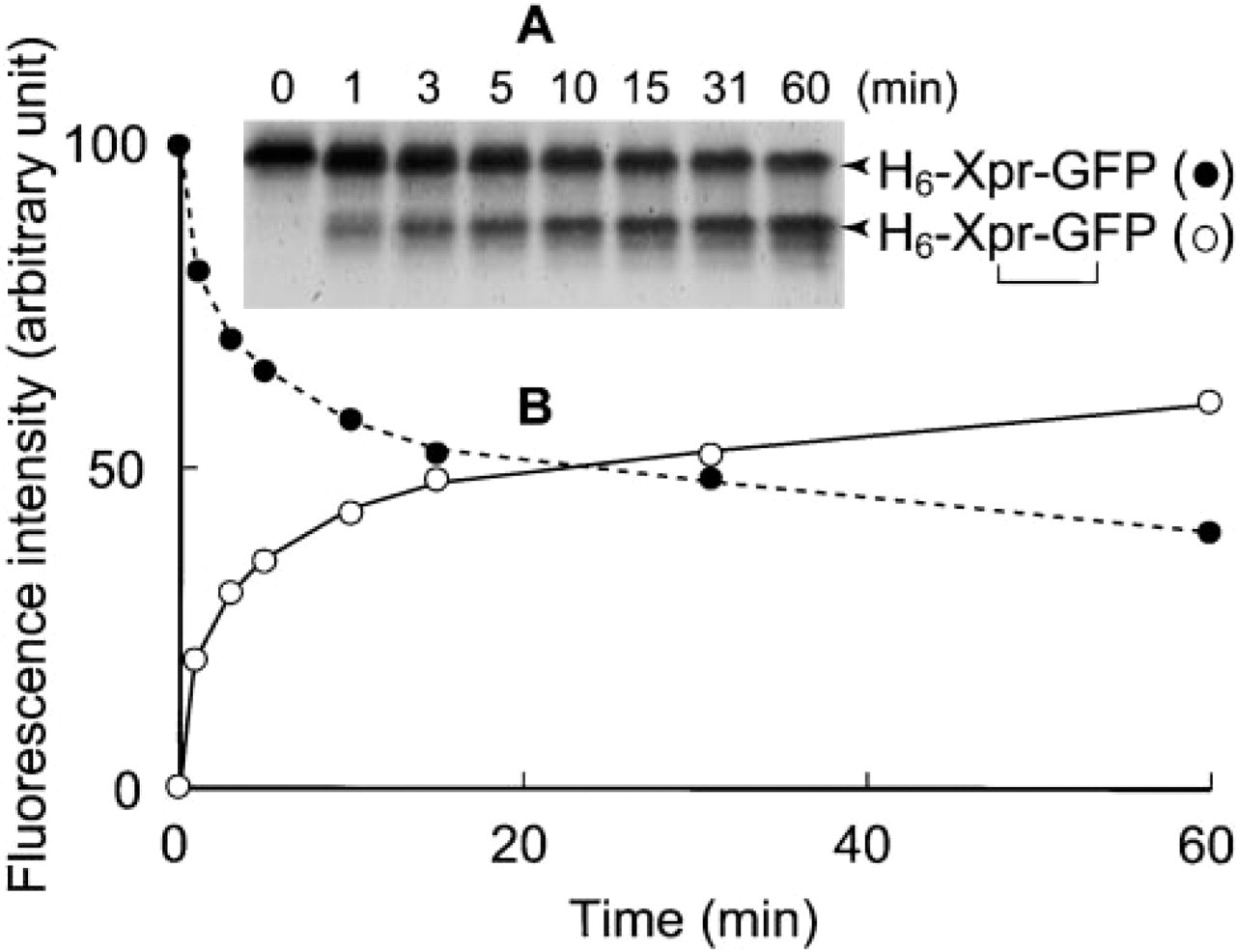
Time course of crosslinking of His6-Xpress-GFP by transglutaminase. (
Higher Affinity for Tissue Transglutaminase
There are several types of transglutaminase that are encoded by separate genes, including keratinocyte transglutaminase, plasma transglutaminase (blood coagulation factor XIIIa, FXIIIa), tissue transglutaminase, and prostate gland transglutaminase. Among these, plasma and tissue transglutaminases are widely distributed in the body. We therefore determined whether the fluorescent substrate His6-Xpress-GFP serves as a substrate for plasma transglutaminase as well as for tissue transglutaminase by first activating the purified preparation of plasma transglutaminase with thrombin and then mixing it with the substrate. His6-Xpress-GFP turned out to be a very poor substrate for plasma transglutaminase (Figure 4). When both enzyme activities were adjusted to the same level by using the synthetic substrate 5-(biotinamido)pentylamine and their crosslinking abilities toward His6-Xpress-GFP were compared, plasma transglutaminase exhibited only weak activity. This weak activity of plasma transglutaminase toward His6-Xpress-GFP is consistent with its narrow substrate specificity; it is highly specific for fibrin.
Detection of Transglutaminase Activity in Tissue Extracts
To evaluate the usefulness of His6-Xpress-GFP as a transglutaminase substrate, we prepared guinea pig liver homogenate, incubated it with His6-Xpress-GFP, and analyzed the reaction products by SDS-PAGE (Figure 2B). Incubation of the fluorescent substrate with the liver homogenate resulted in its intra- and inter-molecular crosslinking (Figure 2, Lanes 6 and 8), indicating that His6-Xpress-GFP is sensitive enough to detect the transglutaminase activity in tissue extracts.
Detection of Transglutaminase Activity on Tissue Sections
Figure 5 shows that His6-Xpress-GFP can also be used for histological purposes, i.e., activity staining of transglutaminase on tissue sections. When incubated with non-fixed frozen sections of a variety of guinea pig tissues shown in Figure 5, His6-Xpress-GFP was crosslinked to certain discrete structures, such as the stratified squamous epithelium of the skin, smooth muscle layers of the blood vessels and ducts, and connective tissues, giving a clear fluorescent pattern of the localization of active transglutaminase. In general, transglutaminase activities appear to be relatively concentrated in the ECM of various tissues.
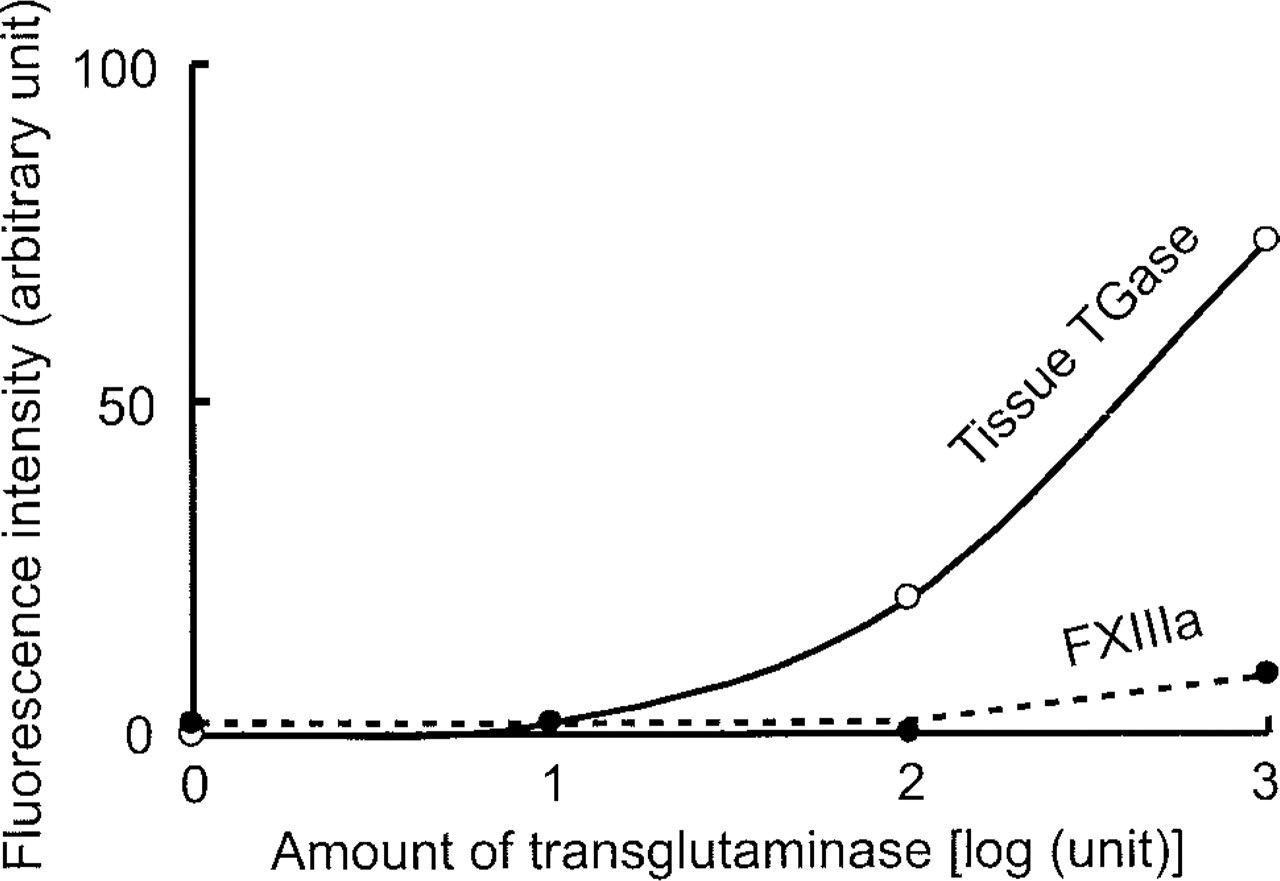
Lower affinity of His6-Xpress-GFP to FXIIIa than to tissue transglutaminase. Variable amounts of purified preparations of FXIIIa (closed circles) and liver transglutaminase (tissue TGase; open circles), whose relative activities were adjusted using 5-(biotinamido)pentylamine as a standard substrate, were incubated with a constant amount of His6-Xpress-GFP at 37C for 1 hr and the reaction products were analyzed as in Figure 3.
In the esophagus, very bright fluorescent staining was seen in the endomysium of the muscularis mucosae (Figure 5A). In the skin, fluorescent labeling was seen in the upper spinous and granular layers of epidermis (Figures 5B and 5R), where membrane-bound keratinocyte transglutaminase has been demonstrated by immunohistochemistry (Michel and Demarchez 1988). In the liver, blood vessels and bile ducts were strongly decorated with His6-Xpress-GFP; the reticular network around sinusoids was also weakly stained (Figure 5C). In the lung, blood vessels, bronchioles, and connective tissue elements of alveoli were stained (Figure 5D), in the testis, the fibroreticular lamina (Figure 5E), and in the pituitary, connective tissues, especially thin layers of connective tissues surrounding endocrine cells in the pars distalis (Figure 5F). In the aorta, elastic lamellae of the tunica media (Figure 5G) were stained, in the spleen, reticular scaffolding of the sinuses (Figure 5H), and in the large intestine, muscularis mucosae and subserosal connective tissues and endomysium of muscularis externa (Figure 5J). In the kidney, connective tissues of the tubules (Figure 5K) were stained, in the seminal vesicle, connective tissues of smooth muscle and mucosa fold (Figure 5L), and in the urinary bladder, endomysium of the smooth muscle (Figure 5M). In the small intestine, dense connective tissues of submucosa, connective tissues of lamina propria and serosa, and endomysium of muscularis externa (Figure 5N) were stained, in the thyroid, connective tissues surrounding the folicular cells (Figure 5O), in the tongue, blood vessels and endomysium of the inferior longitudinal muscle (Figure 5P), and in the heart, endomysium (Figure 5T). These labelings were not present on the control sections processed in parallel in the presence of EDTA (e.g., Figures 5I and 5Q) or putrescine (e.g., Figure 5S). The degree of cellular labeling was greatly reduced when tissue sections were washed extensively with PBS before incubation with His6-Xpress-GFP, whereas the labeling of ECM and connective tissues remained relatively unaltered. As a typical example, the staining patterns of heart sections with and without washing are shown in Figures 5T and 5U. This result suggests that the weak cellular labeling is mainly due to the soluble tissue transglutaminase (transglutaminase 2 or TGC) and that the enzyme responsible for the relatively strong labeling of ECM is tightly associated with their components.
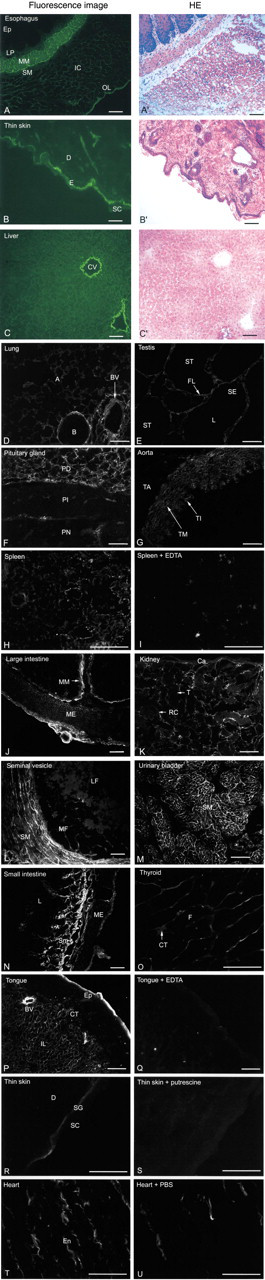
Locations of transglutaminase visualized by activity staining of guinea pig tissue sections. Fresh tissues were frozen, sectioned in a cryostat, and stained with His6-Xpress-GFP (
Discussion
Two types of histological methods have been used for determining cellular localization of transglutaminase: immunohistochemistry and activity staining. In the previous activity staining method, active transglutaminases are detected on tissue sections by incubation with monodansylcadaverine (Aeschlimann and Paulsson 1991) or 5-(biotinamido)pentylamine (Perry et al. 1995) and then by visualizing the incorporated monodansylcadaverine or 5-(biotinamido)pentylamine with anti-dansyl antibody or FITC-labeled avidin. The in situ transglutaminase assay reported here is relatively highly sensitive and therefore does not require amplification with anti-GFP antiserum, making the procedure simple and reliable. Another advantage is that His6-Xpress-GFP has both reactive Gln and Lys residues that serve as an acyl donor and an acceptor, respectively, whereas monodansylcadaverine and 5-(biotinamido) pentylamine can serve only as acyl acceptors. The GFP fluorescence was very stable compared to that of FITC, and practically no fading occurred during observation with a normal fluorescence microscope equipped with FITC optics. Helped by this sensitivity, stability, and simplicity, we could perform a large number of activity stainings of transglutaminase in a series of guinea pig tissues and found a surprisingly wide distribution of the crosslinking enzyme, especially in the ECM. This result demonstrates the previously unrecognized importance of transglutaminase in maintaining the integrity of ECM as well as the well-established roles in maintaining the barrier function of the skin (Hohl 1990; Matsuki et al. 1998; Ishida–Yamamoto and Iizuka 1998; Nemes and Steinert 1999) and in stabilizing fibrin clots (Muszbek et al. 1996; Mosesson 1998). The strong staining of the extracellular matrix mentioned above was somewhat unexpected because (a) tissue transglutaminase lacks the presequence, (b) keratinocyte transglutaminase is membrane-bound (Rice et al. 1992), and (c) the extracellular transglutaminase FXIIIa has very weak affinity for the fluorescent substrate His6-Xpress-GFP. But the result is consistent with the accumulating evidence that, although the mechanism is unknown, a high proportion of transglutaminase is secreted into the extracellular space and remains there (Aeschlimann et al. 1993; Nurminskaya et al. 1998; Gaudry et al. 1999). Identification by Dieterich et al. (1997) of tissue transglutaminase as the endomysial autoantigen recognized by serum samples of patients with celiac disease is also consistent with our staining results. Celiac disease is characterized by small intestinal damage caused by autoantibodies against components of the fibroreticular scaffolding and endomysium, a structure of the smooth muscle connective tissue. The work of Dieterich et al. (1997) therefore strongly suggests that transglutaminase is one of the components of the endomysium.
Our original intention to devise an in situ transglutaminase assay was to demonstrate the presence of active transglutaminase in the brain and in the nucleus of the cell, the locations currently receiving considerable attention in relation to transglutaminase and neuronal development (Hand et al. 1993) and neurodegenerative disorders (Lorand 1996; Igarashi et al. 1998; Kahlem et al. 1998; Chen and Mehta 1999) and regulation of nuclear functions (Shimizu et al. 1996; Lesort et al. 1998; Peng et al. 1999). However, we could not detect significant levels of the activity in these structures, probably because of very low levels of transglutaminase and insufficient sensitivity of the assay for covering such low levels. Alternatively, in these structures the expression of transglutaminase activity may be induced under certain abnormal conditions, such as diseases and apoptosis. In the present study, we demonstrated the usefulness of a GFP derivative, His6-Xpress-GFP, for visualizing active transglutaminase on normal histochemical sections. It would also be useful for determining locations and levels of transglutaminases in clinical specimens. Another potential use of His6-Xpress-GFP would be for preparing GFP-labeled proteins which, in turn, can be used as fluorescent histochemical probes. For example, many antibodies, lectins, and protein ligands are expected to be able to be labeled with His6-Xpress-GFP in the presence of transglutaminase through their Gln and Lys side chains.
Footnotes
Acknowledgments
Supported by Grants-in-Aid for Scientific Research from the Ministry of Education, Science, Sport and Culture of Japan, by a Research Grant for Cardiovascular Diseases (11C-1) from the Ministry of Health and Welfare of Japan, and by an SRF Grant for Biomedical Research. YF is supported by a Research Fellowship for Young Scientists from the Japan Society for the Promotion of Science.
We thank Dr Yuji Saito and Hiroo Takahashi for discussion and help in the FXIIIa assay, and Setsuko Satoh for secretarial assistance.