
Abstract
By applying the highly sensitive cytochemical Gautron's technique, we were able to reveal AChE activity in rat pancreatic acinar cells, particularly at the level of a complex membrane-bound network formed by tubules with varicosities located around the nuclei and close to the basolateral membrane. The Golgi apparatus was devoid of cytochemical reaction beside the trans-Golgi network cisternae, which showed a positive reaction. The RER of some acinar cells also presented a signal, demonstrating their capability of synthesizing AChE. Immunogold using a specific anti-AChE antibody yielded similar results. Double-labeling experiments corroborated the presence of enzyme cytochemical and immunocytochemical signals in the same lysosomal tubular network. Biochemical sedimentation assays confirmed the presence of AChE in acinar cells, which exists as two globular molecular forms, G1 and G4. These results were obtained with pancreatic tissue in situ as well as with isolated acinar cells maintained in culture and devoid of neural elements. The existence of a continuous tubular lysosomal network containing AChE is in agreement with previous reports on acinar and other cell types, and supports a more general hypothesis on dynamic continuities among cell structures. Whether AChE is being secreted by the acinar cells or internalized through this endo-lysosomal system was not defined. However, the capability of the acinar cells to synthesize AChE and to channel it through a tubular system is a good indication that the cells can modulate their cholinergic stimulation for optimal secretion of digestive enzymes.
A
Acetylcholinesterase (AChE) is an essential constituent of this autonomic innervation needed to prevent desensitization of cholinergic receptors and thereby preserve the efficiency of cholinergic regulation of pancreatic secretion. AChE exists in multiple molecular forms, a polymorphism that enables the cells to position AChE at different sites where it may fulfill specific roles (for review see Massoulié et al. 1993). For example, the tetrameric form of AChE, G4, is either membrane-bound or secreted as a soluble enzyme, whereas the complex asymmetric form A12 is specific for the junctional synaptic structures (Massoulié et al. 1993; Legay 2000). In the pancreas, AChE, as revealed by early cytochemical studies, is mainly concentrated within the neural network that provides pancreatic innervation (Coupland 1958; Hiramatsu et al. 1993; Ushiki and Watanabe 1997). However, endocrine cells do exhibit intracellular AChE cytochemical reaction (Ogneva et al. 1989), and acinar cells also may contain this enzyme because several esterases have been found in acinar cell lysosomal structures (Oliver 1983).
Lysosomes are polymorphic structures generated by the Golgi apparatus and belonging to the lysosomal–endosomal system of the cell. Pancreatic acinar cells have been described as displaying two distinct populations of secondary lysosomes. One is represented by the classical Golgi-associated globular, acid phosphatase-positive structures, and the other, located in the perinuclear and basolateral regions, has been described as a tubular network (Oliver 1983; Oliver and Yuasa 1987; Oliver et al. 1989). Studies performed in various cell types, including the acinar cells, have confirmed the existence of lysosomal tubular structures (Beaudoin et al. 1984, 1985; Araki et al. 1989; Robinson and Karnovsky 1991; Kobayashi and Robinson 1991). The existence of such a tubular network supports a more general hypothesis that postulates dynamic continuity between membrane-bound structures that may comprise the endosomal–lysosomal compartment participating in both endocytic and secretory cell processes (Marsh et al. 1986; Swanson et al. 1987; Kristié 1988; Heuser 1989; Hopkins et al. 1990; Tooze and Hollinshead 1991; Sandborn and Bendayan 1996; Stoorvoge et al. 1996; Bendayan and Rasio 1997). Whether both the globular and the tubular lysosomal structures described in pancreatic acinar cells are independent systems or are, in fact, part of the same network remains to be established and constitutes part of the present investigation.
Whereas most studies dealing with AChE in the pancreas were performed to understand the innervation of the organ and the neuronal control of pancreatic secretion, in the present study we took advantage of a highly sensitive cytochemical technique (Gautron 1982) and the immunocytochemical approach (Bendayan 1995) to examine the subcellular distribution of AChE in rat pancreatic acinar cells. Furthermore, biochemical determinations were performed to identify the AChE molecular forms present in the pancreas and, more specifically, those in acinar cells. These combined studies have indicated that acinar cells are capable of synthesizing two globular forms of AChE and that the enzyme is channeled through a lysosomal system formed by a complex network of tubular membrane-bound structures with globular varicosities.
Materials and Methods
Cytochemistry
For the cytochemical detection of AChE, the protocol developed by Gautron (1982) was followed, with some adjustments.
For electron microscopy, pancreatic tissue from adult male Sprague-Dawley rats (100 g) was fixed in situ with 1% glutaraldehyde in 0.15 M sodium cacodylate buffer, pH 7.3. Tissue was then removed from the abdominal cavity and small samples were immersed in the same fixative for a total fixation time of 30 min. On thorough washing, the free aldehyde groups were quenched by adding 0.5 ml of 1 M NaOH to the cacodylate buffer (50 ml). The tissue samples were rinsed and immersed in 0.2 M Tris-maleate-NaOH, pH 6.0, overnight. Incubation was carried out with a substrate medium composed of 0.05 ml acetyl sulfide (Aldrich; Milwaukee, WI) dissolved in 1 ml propylene glycol and diluted in 100 ml 0.2 M Tris-maleate-NaOH, pH 6.0, complemented with 1 ml lead nitrate 3% (w/v) in aqueous solution and 50 mg of acetylthiocholine perchlorate or bromide. Two ml of 10–3 M aqueous solution of iso-OMPA (tetraisopropyl-pyrophosphoramide), a selective inhibitor of butyrylcholinesterase, was added to a final concentration of 2 × 10–5 M. The solution was filtered before adding the acetylthiocholine to remove any precipitate. For the incubation, the tissue samples were immersed in the substrate medium for 10 min at 4C and allowed to warm to room temperature. Incubation times were between 30 and 60 min. After rinsing in the Tris-maleate-NaOH buffer, the tissue was transferred to the cacodylate buffer and postfixed for 60 min with 1% osmium tetroxide. The tissue samples were dehydrated and embedded in Epon according to standard techniques. Ultrathin sections were counterstained with uranyl acetate and lead citrate and examined with a Philips 410SL electron microscope. Thick sections (1 μm) were mounted on grids and examined, without any counterstaining, with a Philips CM30 at 300kV. In this case, most images were electronically registered on CD-ROM.
For light microscopy, rat pancreatic tissue was sampled and cryosections were made according to standard techniques. Five-μm-thick sections were processed for the cytochemical staining as described above. Prefixation with 1% glutaraldehyde was found to yield better results than non-fixed tissues.
Control assays were performed to assess the specificity of the cytochemical reaction. The participation of non-choline esterases to the final reaction product was estimated by inhibiting cholinesterases with eserine sulfate (5 × 10–4 M). The effect of omitting the substrate, acetylthiocholine, on the final reaction product was also tested.
Two types of double-labeling experiments were performed at the electron microscope level. In the first one, we combined pre-embedding cytochemical detection of AChE with postembedding immunocytochemical detection of a pancreatic secretory protein (amylase). The postfixation of the tissue with osmium tetroxide was omitted. For the postembedding immunocytochemical detection of amylase, the protein A–gold technique (Bendayan 1995) was applied in combination with a specific anti-amylase antibody, as described previously (Bendayan 1984). In the second experiment, we combined the pre-embedding cytochemical detection of AChE with the immunogold postembedding detection of AChE as described below.
Immunocytochemistry
For the postembedding immunocytochemical detection of AChE, a rabbit polyclonal antibody (a gift from J. Massoulié) raised against rat brain hydrophobic G4 AChE (Marsh et al. 1984) was used at 1:10 dilution. This antibody was found to be highly specific, recognizing hydro- and detergent-soluble forms of AChE (Marsh et al. 1984). Sections from rat pancreatic tissue fixed in Bouin's fluid and embedded in paraffin, or fixed in 1% glutaraldehyde and embedded in Lowicryl at −20C were used for light and electron microscopy, respectively. For light microscopy, the paraffin sections were rehydrated and incubated with the anti-AChE antibody overnight at 4C. This was followed by a 1-hr incubation with a goat anti-rabbit immunoglobulin antibody coupled to FITC. For electron microscopy, the Lowicryl ultrathin sections were incubated with the anti-AChE antibody overnight and revealed by a 1-hr incubation with the protein A-gold complex (10 nm) (Bendayan 1995). To assess the specificity of the immunocytochemical signals, control experiments were performed, particularly by omitting the primary antibody and by replacing the primary specific antibody with normal rabbit serum.
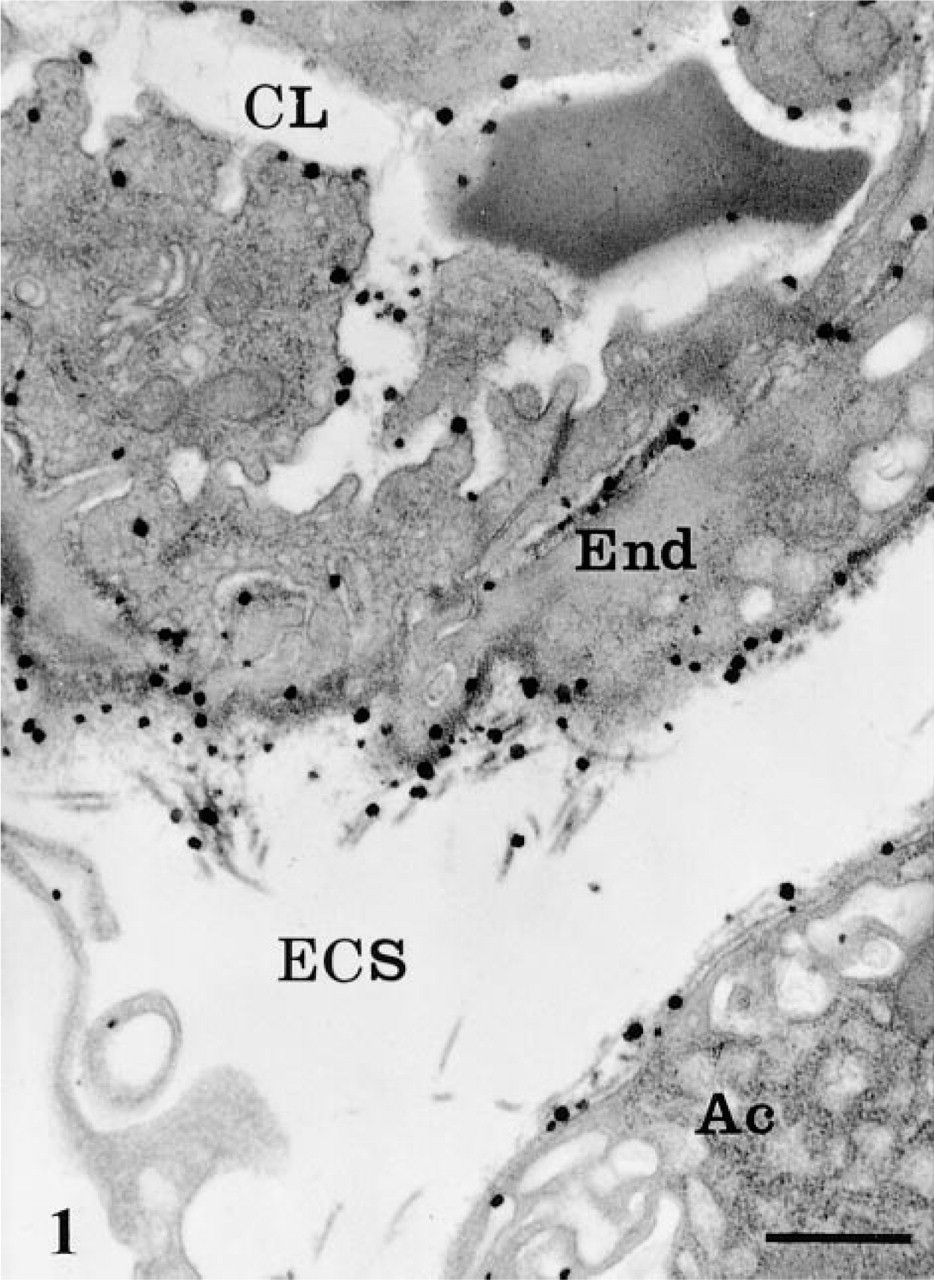
AChE cytochemical reaction in blood capillary vessel and extracellular space in rat pancreatic tissue. The reaction product, as revealed by large electron-dense spots, is present in the capillary lumen (CL) associated with the red blood cell and the endothelial plasmalemmal membranes. In the extracellular space (ECS), it is particularly associated with collagen fibers and endothelial (End) and acinar cell (Ac) basement membranes. Bar = 1 μm.
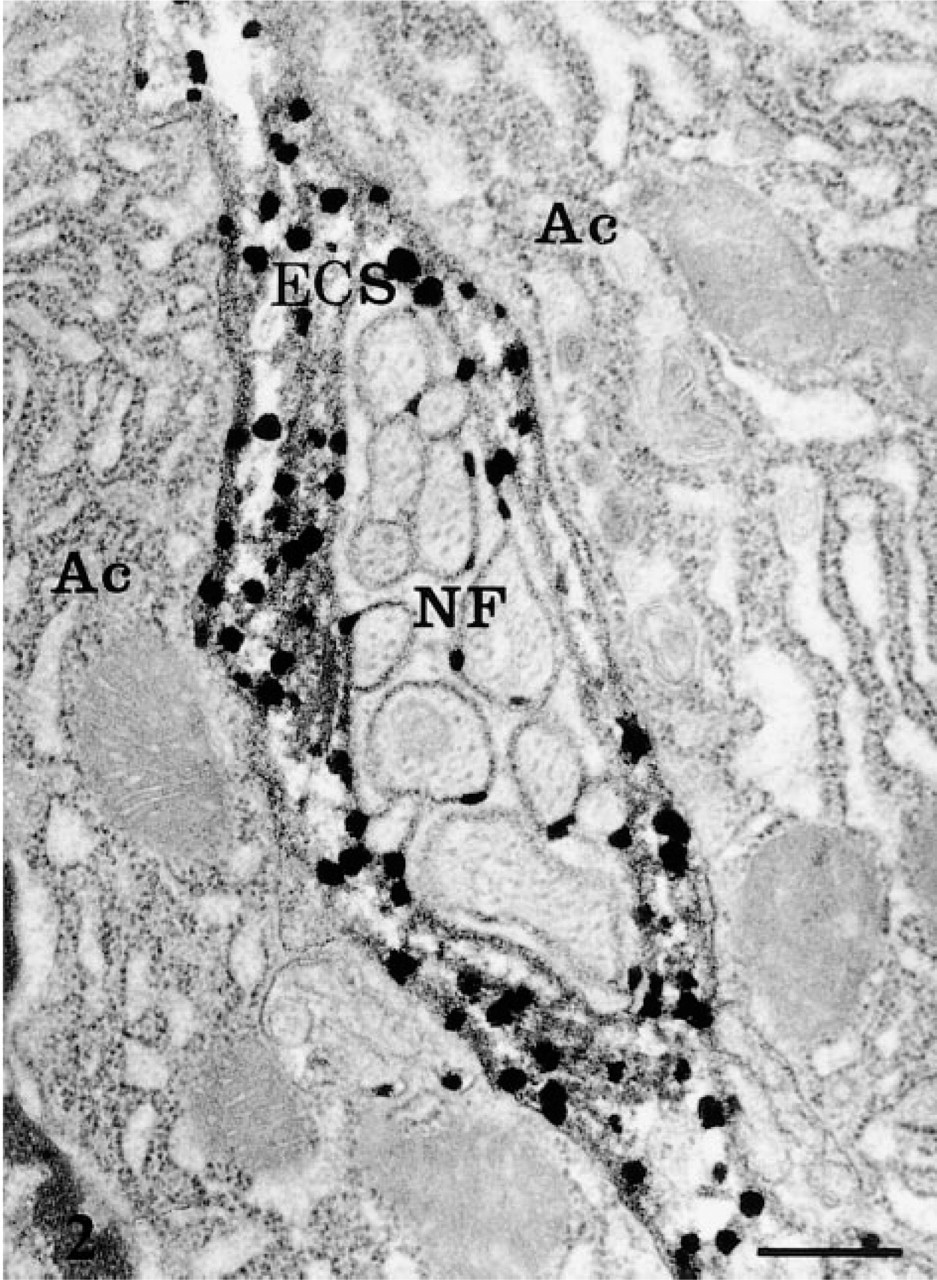
AChE cytochemical reaction product associated with nerve fibers (NF) adjacent to the acinar cells (Ac). Bar = 1 μm.
Acinar Cells in Culture
In addition to the work performed with rat pancreatic tissue fixed in situ, the cytochemical approach was also applied on rat acinar cells maintained in tissue culture. Rat pancreatic acinar cells were isolated and plated on Matrigel-coated Petri dishes as described previously (Bendayan et al. 1986). After 2 days of culture, the cells were fixed in situ with 1% glutaraldehyde and processed as described above for cytochemical demonstration of AChE.
Ganglionic Tissue
As an internal control, we used rat sympathetic superior cervical ganglion (SCG) in parallel with our pancreatic tissue. Neurons of these ganglia are known to contain large amounts of AChE, particularly in their Nissl bodies (Gautron 1974; Gautron and Gisiger 1976; Gisiger et al. 1978; Brzin et al. 1982). Both the cytochemical and immunocy-tochemical protocols were applied on the ganglionic tissue as described above for the pancreas.
AChE Analysis
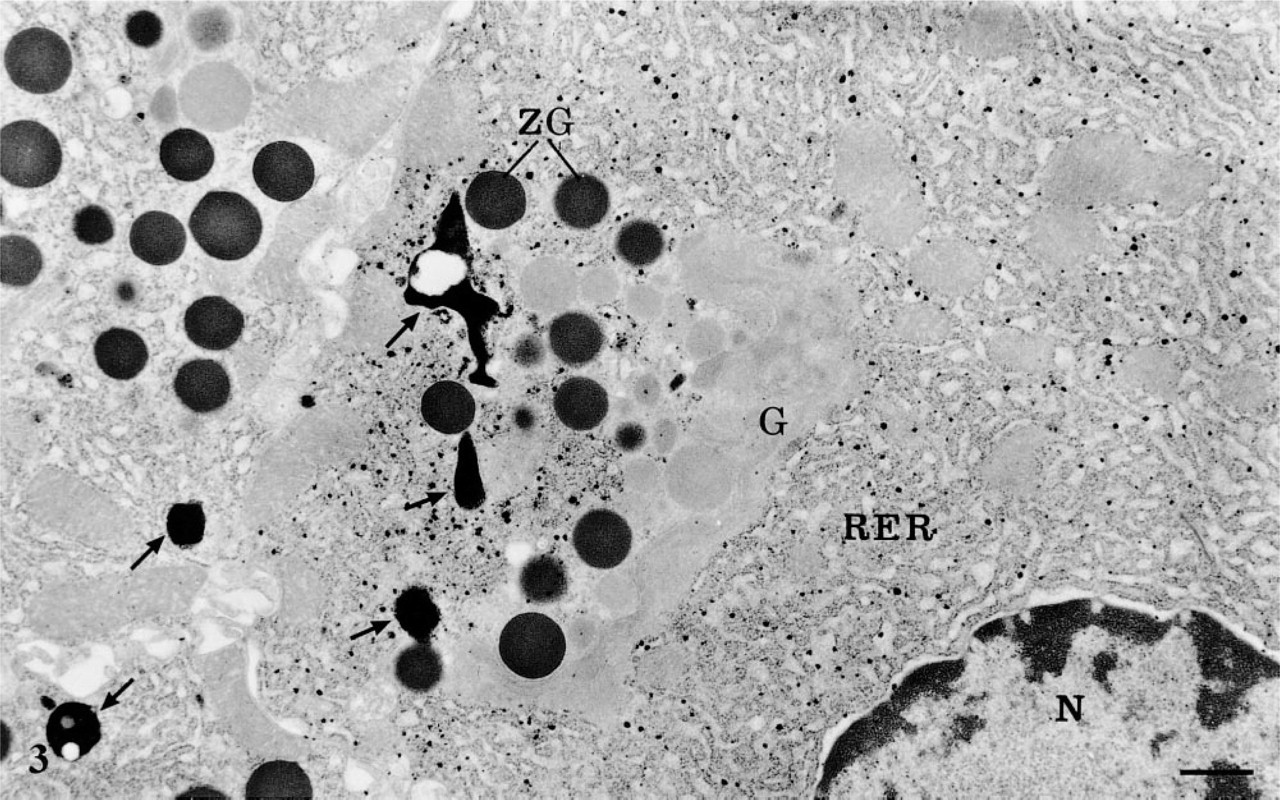
AChE cytochemistry in pancreatic acinar cells. The reaction product is present within the cisternae of the rough endoplasmic reticulum (RER) in one of the cells. The RER of the neighboring cells is devoid of signal. Globular lysosomes (arrows) show strong, dense positive reaction. The Golgi apparatus (G), nucleus (N), zymogen granules (ZG), and mitochondria are free of staining. Bar = 1 μm.
Velocity sedimentation analyses of the AChE molecular forms were performed as described previously (Gisiger and Stephens 1988; Jasmin and Gisiger 1990). Aliquots (50–100 μl) of the supernatants obtained after a low-speed centrifugation (20,000 × g for 15 min) were loaded on 5–20% sucrose gradients and centrifuged in a Beckman SW41 rotor at 40,000 rpm for 19 hr at 3C. Sucrose solutions were made up in a buffer identical to that used for the homogenization, except that the trypsin inhibitor was omitted. Approximately 45 fractions were collected from each gradient and assayed for the AChE activity, using a modified Ellman technique in the presence of iso-OMPA (Gisiger et al. 1978).
Control assays were performed with eserine (5 × 10–4 M) and anticholinesterase BW 284c51 (1,5-bis(4-allyldimethyl-ammoniumphenyl-pentan-3-one dibromide) (2 × 10–5 M).
Results
By light microscopy (results not shown), the AChE cytochemical approach revealed a signal consisting of dark deposits over some acinar cells. These cells were scattered among the acinar parenchyma. Nerve paths were also delineated with a much more intense signal, in a pattern consistent with that reported previously (Coupland 1958; Radke et al. 1985; Luiten et al. 1986).
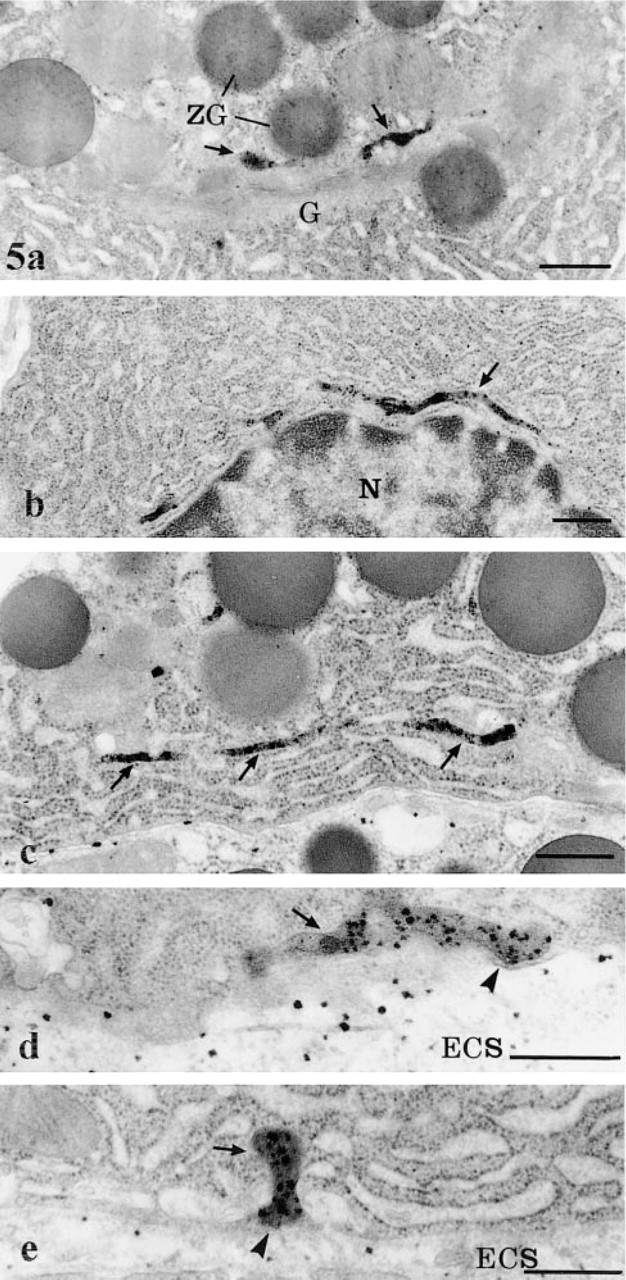
By electron microscopy, the cytochemical reaction product was detected within the pancreatic cells, the neural elements, blood vessels, and in the extracellular space. The reaction product was characterized by small electron-dense spots of about 50 nm (Figures 1 and 2). Within capillaries, the reaction was encountered in the lumen (Figure 1), consistent with the well-established presence of AChE in blood (Koelle 1963). It was also associated with the luminal and abluminal endothelial plasmalemmal membranes (Figure 1). In the interstitial space, the reaction product was located in the subendothelial region around the blood capillaries (Figure 1) and along the basolateral plasma membrane of the acinar cells, particularly in regions adjacent to nerve fibers (Figure 2). The reaction product was often clearly associated with the basement membranes and the collagen fibers (Figures 1 and 2). In the acinar cells, the reaction product was detected in several cellular compartments (Figure 3). The most common location consisted of the lysosomal structures, which were labeled by highly electron-dense homogeneous material (Figure 3). The positive lysosomal structures displayed the classical globular shape and were located close to the Golgi area and along the basolateral membrane. In several instances the globular lysosomes presented elongated protrusions that extended into tubular formations (Figure 4). Both the globular and tubular structures were labeled. In addition, snake-like tubular structures delineated by smooth membranes and displaying positive cytochemical reaction were also revealed (Figure 5). These were located close to the Golgi areas, around the nuclei and neighboring the basolateral membrane of the acinar cells, with very close contacts with the plasma membrane itself (Figures 5d and 5e). The reaction product displayed by these snake-like lysosomal structures consisted of a dense homogeneous material together with a spotty (50 nm) highly dense product resembling that found in the extracellular space (Figures 1, 2, 4, and 5).
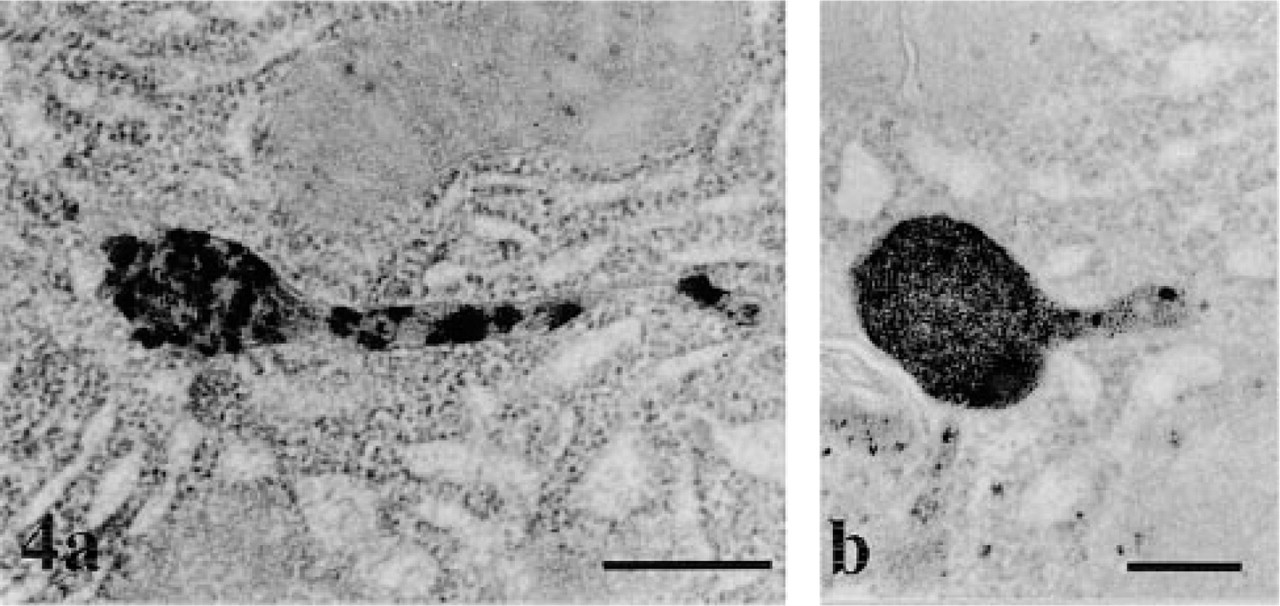
AChE cytochemistry in pancreatic acinar cells. The figure illustrates two globular lysosomal structures displaying strong cytochemical reaction. Both lysosomes present tubular extensions that also display cytochemical reaction product. Bars = 0.5 μm.
In addition to the reaction product found in the lysosomal network, some acinar cells, in an isolated fashion, demonstrated a strong reaction in their rough endoplasmic reticulum (RER) (Figure 3). The reaction product was present within the lumen of the reticulum and displayed only a spotty appearance, resembling, again, that of the extracellular space. This reaction product was restricted to the RER and was not present in the Golgi apparatus and the secretory granules, although in many instances the trans-Golgi network did show a positive signal (Figure 5a). It was also noteworthy that cells with positive reaction in their RER were isolated: neighboring cells, even those of the same acinus, exhibited an RER free of any cytochemical reaction (Figure 3). In fact, the labeling for AChE within the RER displayed a mosaic-like pattern.
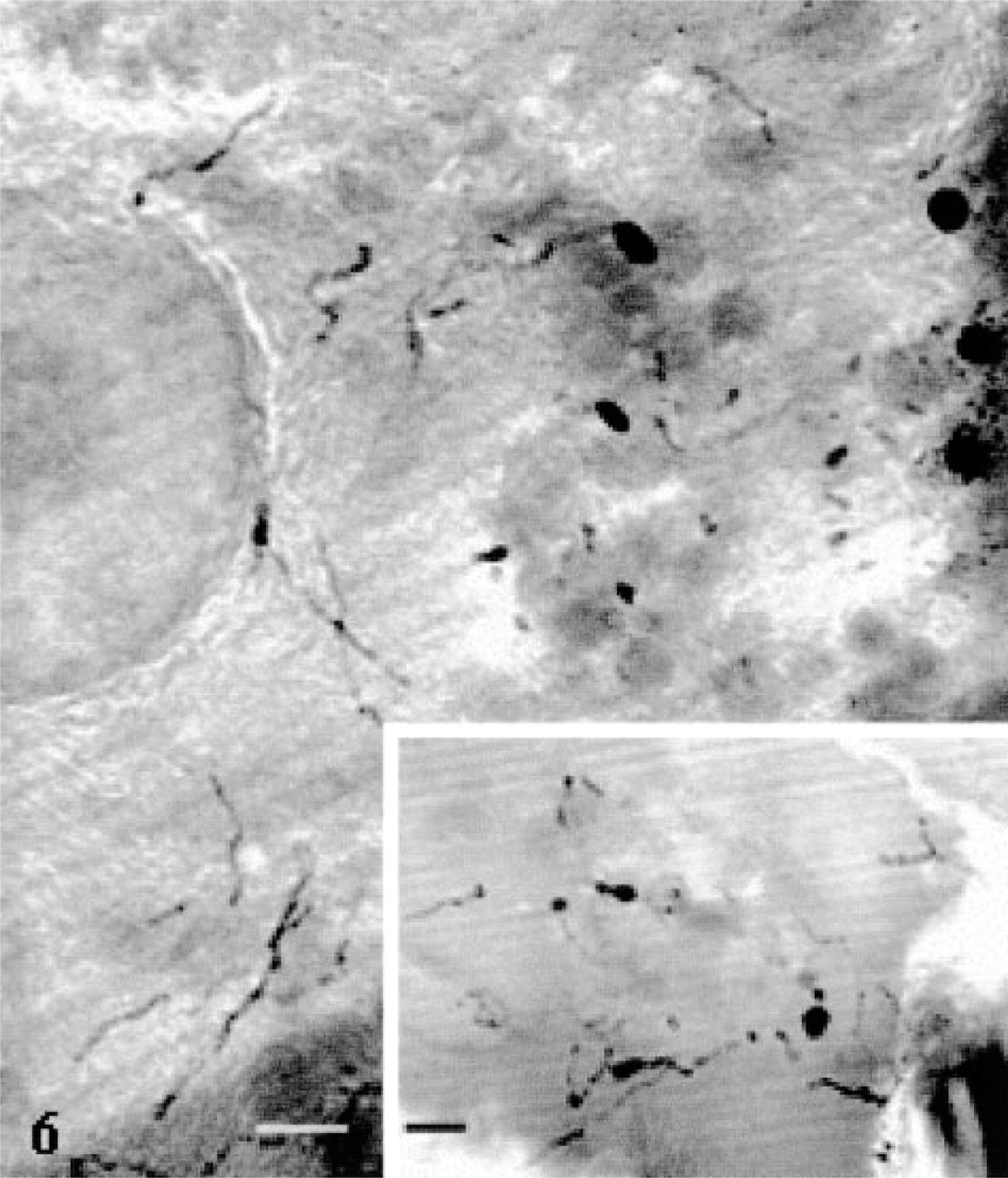
High-voltage electron microscopy of thick sections allowed the delineation of the tubular system in the acinar cells in a more extended fashion (Figure 6). Long tubular membranous structures, usually presenting various varicosities and bifurcations, were conspicuous throughout the cell cytoplasm, particularly in the basolateral and nuclear regions (Figure 6). They formed a complex 3D tubular network. Some of the tubules appeared to come in very close contact with the basolateral membrane (Figure 6, inset). In particularly well-oriented sections passing through the Golgi apparatus, a complex network of tubules with connected vesicles, as well as positively stained dense globular structures, were observed (Figure 7). This network could well represent the positively stained trans-Golgi network (TGN) as seen on thin sections (Figure 5a). It did display the classical TGN profile described in many secretory cells (Rambourg and Clermont 1990).
AChE cytochemistry in pancreatic acinar cells. (
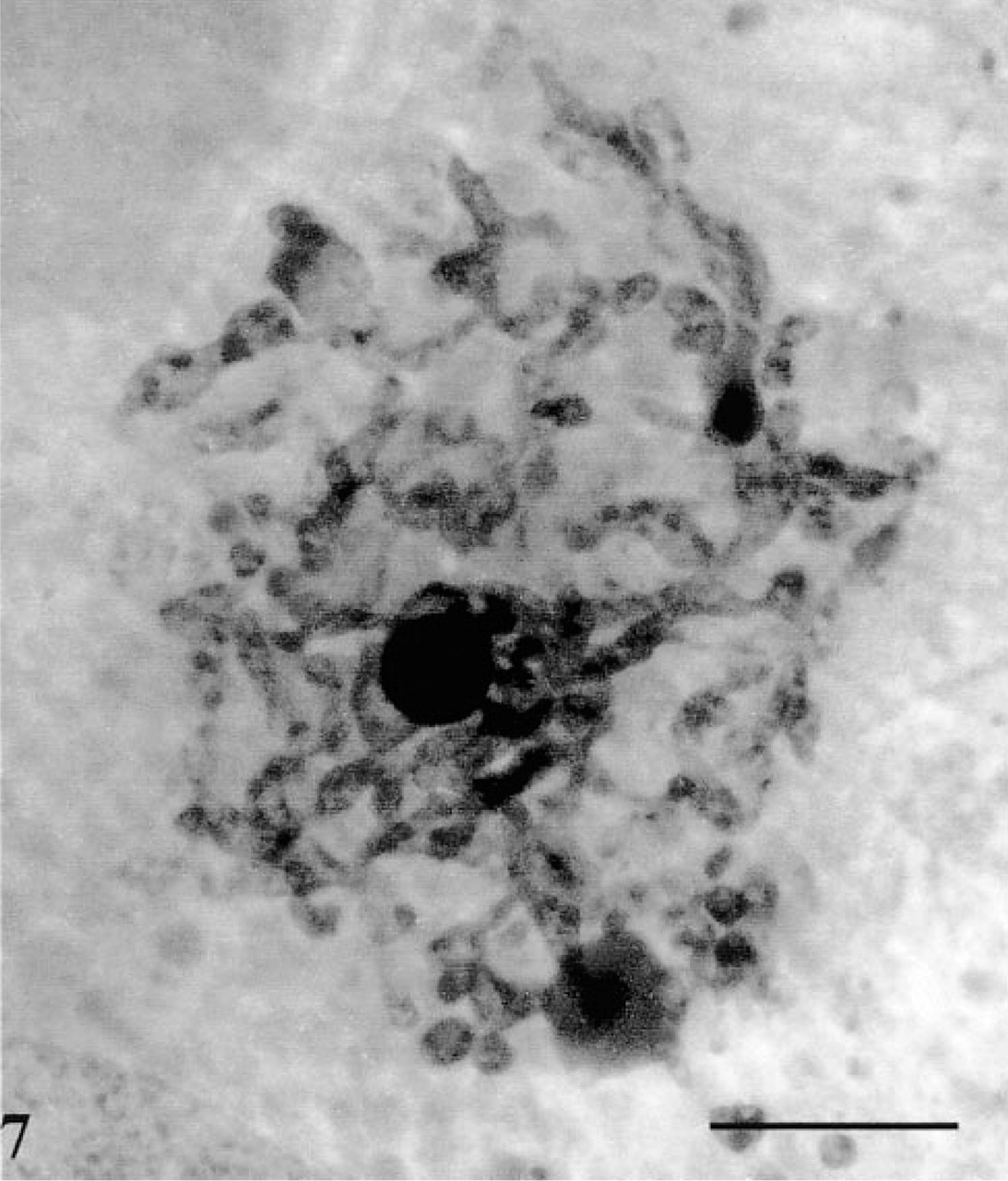
An extended network of positively stained tubules and vesicles together with a few intensely stained globular structures. This network may well represent the trans-Golgi network of the cell. Bar = 0.5 μm.
In addition to the exocrine parenchyma, the endocrine cells in the islets of Langerhans also displayed AChE-positive tubular structures, although the lysosomal network appeared less well developed than in the acinar cells (result not shown).
The double-labeling experiment in which the AChE reaction was combined with the immunocytochemical detection of a secretory protein, i.e., amylase, demonstrated that the AChE-positive compartments, in addition to the RER, are in fact devoid of pancreatic secretory amylase (Figure 8).
Under the control conditions tested, the reaction product was markedly reduced or eliminated. In the presence of eserine, the labeling found in the extracellular space was faint, whereas the globular and tubular lysosomes remained somewhat labeled, although with much reduced intensity. Removal of the acetylthiocholine from the incubation medium, on the other hand, abolished all labeling (not shown).
Overall, very similar results were obtained in acinar cells maintained in culture. The AChE reaction product was present in the globular and tubular lysosomal structures (results not shown).
Application of the immunocytochemical approach for detection of AChE on pancreatic tissue led to labeling at the level of the acinar cells. By light microscopy, the fluorescent staining within acinar cells displayed a punctate pattern. This was mainly located along the basolateral membrane and around the nuclei (not shown). The apical region of the cells, rich in secretory zymogen granules, was devoid of staining. On the other hand, strong signal was present along nerve paths. By electron microscopy, the labeling by gold particles, although of low intensity, was mainly detected in globular lysosomal structures, some of them displaying short tubular extensions, and in the RER (Figure 9a). In the extracellular space, the signal was mainly along the collagen fibers (Figure 9b). Double-labeling techniques combining enzyme cytochemistry and immunogold were able to confirm the location of the AChE (Figure 10). Globular positive structures with tubular extensions, as well as snakelike tubular structures, were simultaneously labeled by the enzyme cytochemical reaction product and the immunogold. However, the enzyme reaction product was much reduced in intensity compared to that obtained with the single-labeling protocol. This is probably due to extraction of the enzyme reaction product, particularly in areas containing small amounts of cytochemical reaction, during immunocytochemical processing. However, in spite of such extraction, it was possible to identify the globular and tubular AChE-positive network simultaneously labeled by enzyme and immunocytochemistry.
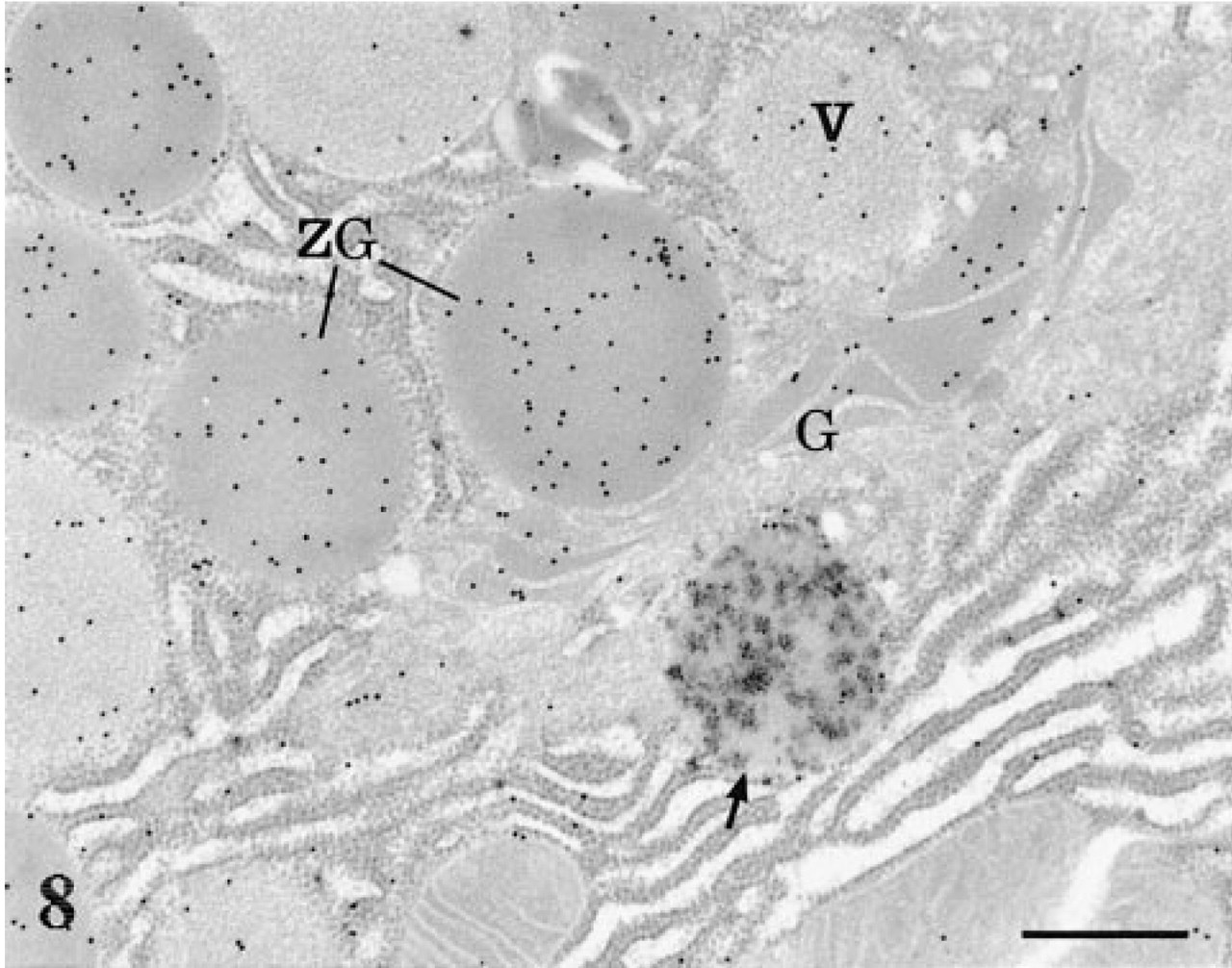
Double labeling. AChE-labeled pancreatic tissue section was processed for immunogold labeling of amylase. The AChE cytochemical reaction is present in a globular lysosome (arrow), while the gold particles revealing amylase antigenic sites are present in the Golgi cisternae (G), the condensing vacuoles (V), and the zymogen granules (ZG). No overlapping of both stainings is observed. Bar = 0.5 μm.
Control experiments confirmed the specificity of the immunocytochemical results, because omitting the primary antibody or using normal rabbit serum led to absence of any labeling.
The parallel experiments performed on SCG supported our cytochemical and immunocytochemical results. The AChE cytochemical reaction was present at the level of the Nissl bodies, the entire Golgi apparatus, and in the extracellular space surrounding the ganglionic cells. These results were identical to those published previously (Gautron 1974; Gautron and Gisiger 1976; Gisiger et al. 1978; Brzin et al. 1982). In the immunocytochemical demonstration of AChE in ganglionic cells, the gold particles revealed the presence of antigenic sites within the cisternae of the RER forming the Nissl bodies (results not shown).
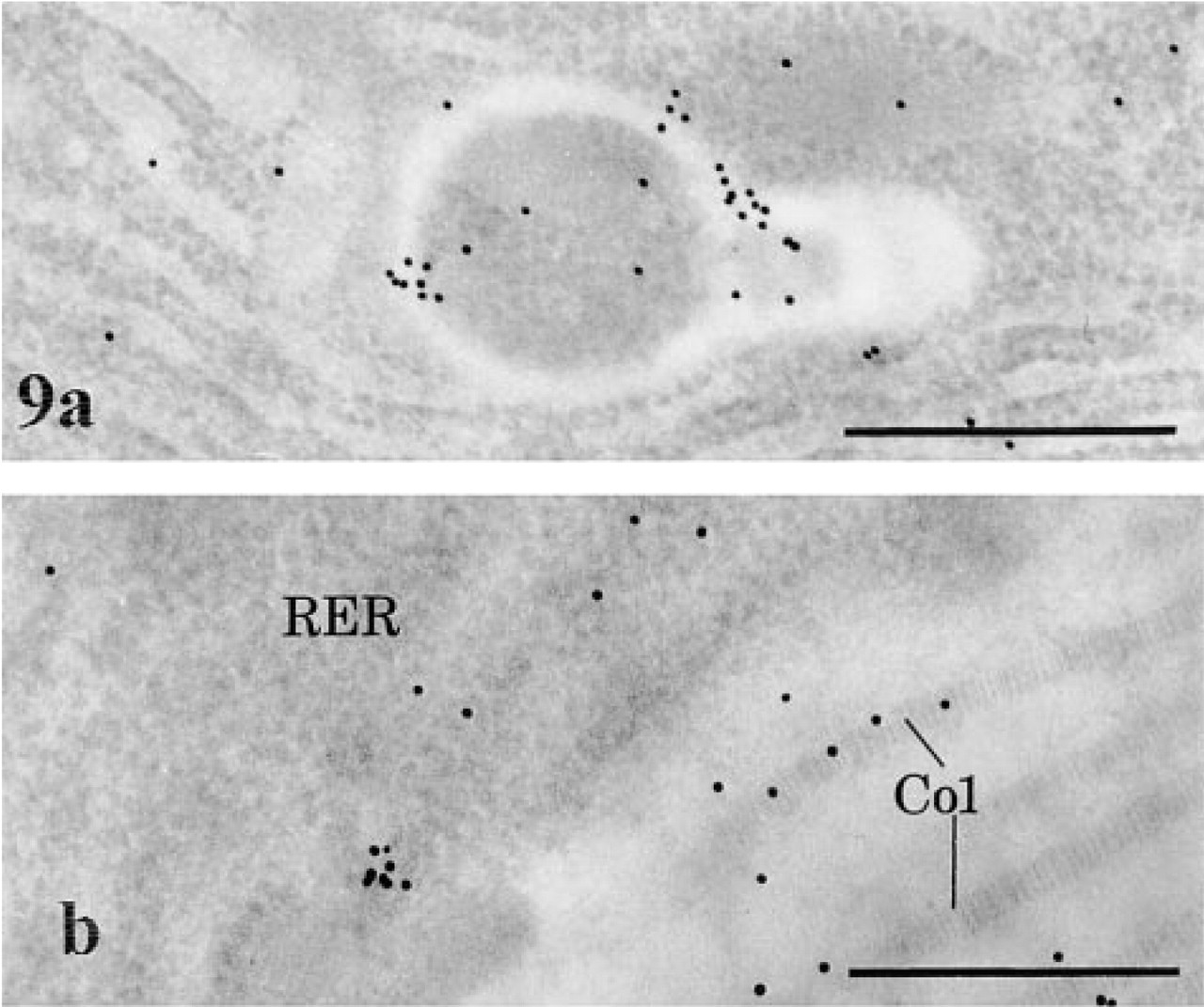
AChE immunocytochemistry in pancreatic acinar cells. (
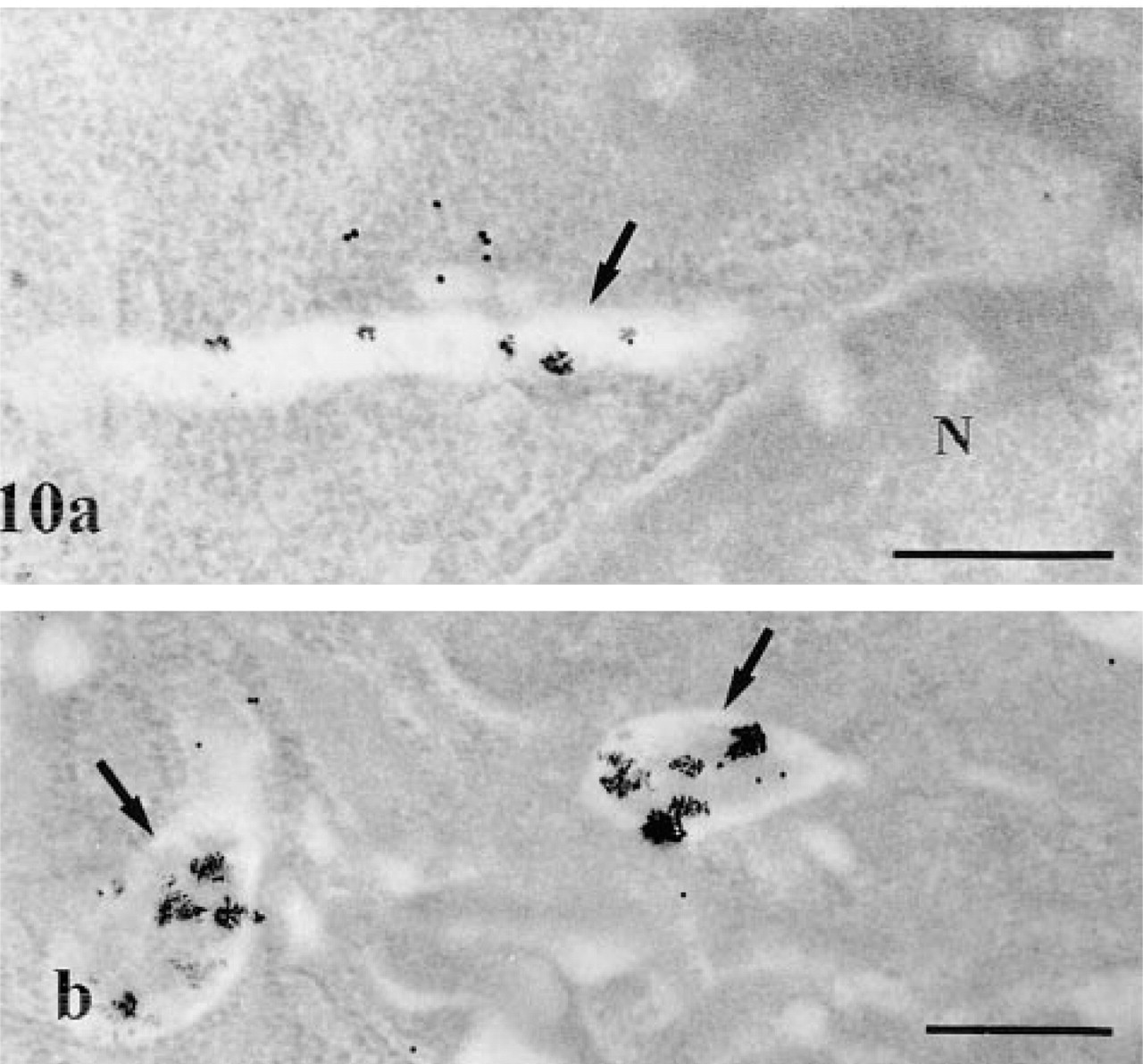
AChE cytochemistry and immunocytochemistry; double-labeling protocol. The cytochemical reaction, although of low intensity, and some gold particles are present in tubular (
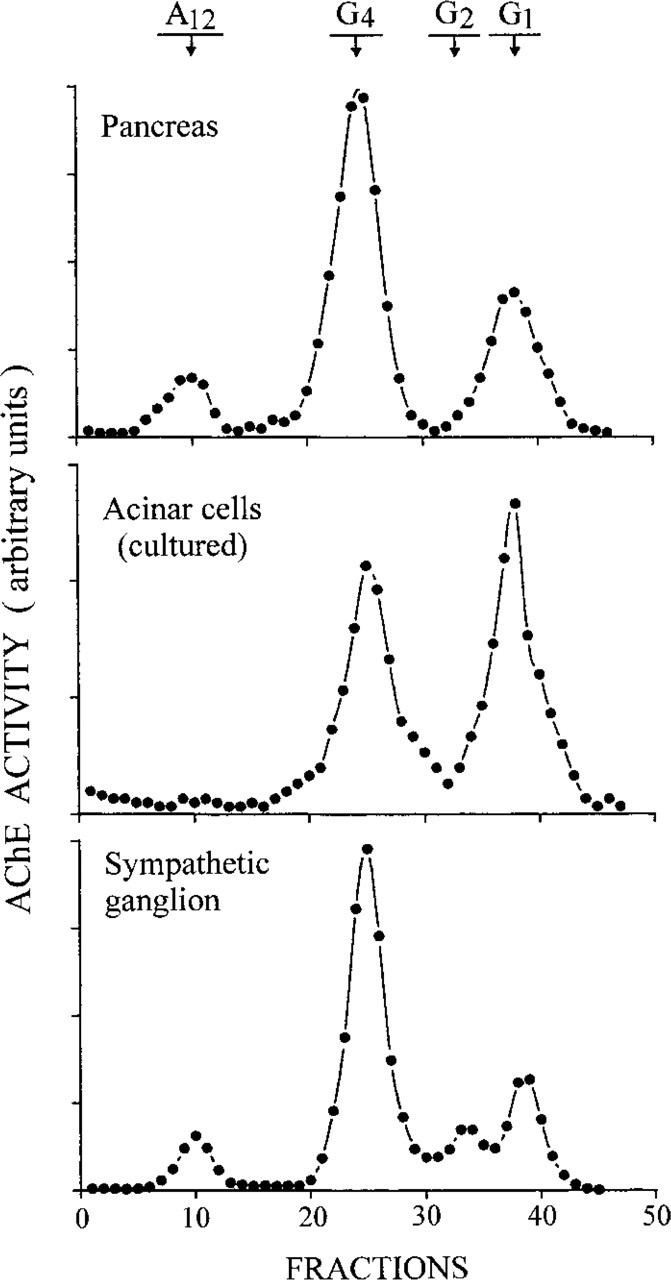
Representative distribution of AChE molecular forms, as established by velocity sedimentation analysis of extracts obtained from pancreas (top), from primary cultures of isolated pancreatic acinar cells (middle), and from SCG (bottom). The AChE profile of the pancreatic extract is similar to that of the SCG. The A12 form is present in the pancreatic and SCG tissues but absent in the isolated culture cells. Acinar culture cells displayed only the G1 and G4 forms.
To complete this study, we performed sedimentation analyses to examine the AChE molecular form content of the pancreas and of isolated acinar cells in comparison to that of the SCG (Figure 11). The sympathetic ganglion yielded its well-established profile of AChE molecular forms characterized by a largely predominant G4 peak and the presence of significant amounts of the asymmetric A12 form (Gisiger et al. 1978; Brimijoin 1983). The pancreatic tissue produced an AChE profile very similar to that of the SCG, except for the absence of the G2 form. This similarity was not unexpected because of the great abundance of ganglionic cells in the pancreas, which bear the bulk of AChE present in the gland. This is consistent with the presence in the pancreatic tissue of significant amounts of the A12 form, which is known to be related to nicotinic synaptic structures (Massoulié et al. 1993). In contrast, the primary cultures of isolated acinar cells contained only two AChE molecular forms, G1 and G4, both present in similar proportions (Figure 11). In the absence of neural elements, the origin of these AChE molecular forms is likely to be the acinar cells themselves. To ascertain that the enzymatic reaction resulted from AChE, control assays were performed with specific inhibitors. Eserine sulfate, which specifically inhibits cholinesterases, abolished all peaks of enzymatic activity. The same was true when BW 284c51, a selective inhibitor of AChE, was added to the Ellman medium, which already contains iso-OMPA, a selective inhibitor of butyrylcholinesterase.
Discussion
This study has revealed the presence of AChE in rat pancreatic tissue and, more specifically, in pancreatic acinar cells. That AChE is present in pancreas has been previously reported through cytochemical studies (Coupland 1958; Hiramatsu et al. 1993; Ushiki and Watanabe 1997). However, these focused mainly on the innervation of the organ and have delineated the AChE-rich neural network wiring the pancreas. Here, using Gautron's highly sensitive cytochemical technique (Gautron 1982) together with the immunogold approach (Bendayan 1995), we were able to demonstrate the presence of AChE not only in the extracellular space, in association with neural elements and connective tissue, but also in the acinar cells. This finding was corroborated by our biochemical approach. In particular, sedimentation analysis revealed that isolated pancreatic acinar cell preparations devoid of neural elements contain the two molecular forms, G1 and G4, of AChE. Moreover, A12, the AChE molecular form specific for the synaptic junction (Massoulié et al. 1993) and present in extracts of the pancreatic organ, was conspicuously absent in the isolated cells. This further strengthens the conclusion that part of the AChE found in the pancreas originates from the acinar cells.
In the acinar cells, the AChE cytochemical reaction was observed in globular lysosomal structures located in the Golgi area and the basolateral region of the cell, as well as in a particular lysosomal compartment with a tubular snake-like appearance delineated by smooth membranes. Images of globular lysosomes extending into tubular projections, both displaying AChE activity, indicate continuity between the globular and tubular lysosomal structures. Examination of thick sections by high-voltage electron microscopy supported the existence of such a continuous complex network of tubular structures displaying varicosities and bifurcations. These snake-like tubular lysosomes form a complex 3D network within the acinar cell, inter twined between the cisternae of the ER, around the nuclei, and in the basolateral region, coming into very close contact with the plasma membrane itself. Similar networks were previously demonstrated in several types of cells, including pancreatic cells, and were shown to contain various lysosomal enzymes, such as adenine dinucleotide phosphatase, trimetaphosphatase, aryl sulfatase B, thiolacetic acid, and several other esterases (Oliver 1983; Beaudoin et al. 1984; Oliver and Yuasa 1987; Robinson and Karnovsky 1991; Alvares 1994; Robinson et al. 1996). However, two populations of lysosomal structures were ascribed to acinar cells (Oliver et al. 1989), one corresponding to the classical acid phosphatase-positive globular structures and a second corresponding to the tubular system and considered as acid phosphatase-negative. However, Araki et al. have shown that tubular and globular lysosomal structures in acinar cells are in fact acid phosphatase-positive and form a single complex network (Araki et al. 1989; Araki and Hatae 1999). Our results support such a view with respect to the AChE-positive network. Our high-resolution cytochemistry on thin sections and high-voltage electron microscopy on thick sections have shown that globular and tubular structures are indeed connected, forming an AChE-positive continuous network.
In some acinar cells, AChE was also present within the cisternae of the RER, indicating that the acinar cells do synthesize this enzyme. Despite their presence in the RER, both the cytochemical and immunocytochemical reactions were absent from the main body of the Golgi apparatus and from the secretory zymogen granules, although the trans-Golgi network did display a positive reaction. The absence of AChE in the Golgi is further supported by the double-labeling protocol revealing AChE and amylase, a pancreatic secretory digestive enzyme, which showed that in addition to the RER where both molecules are present, AChE is absent from the classical regulated secretory pathway. The absence of AChE from the different cisternae of the Golgi could be related to the nature of the AChE molecules present in acinar cells. The A12 asymmetric molecules composed of globular catalytic tetramers and a collagen tail are assembled in the Golgi compartment (Rotundo 1984; Legay et al. 1999). Previous studies (Gautron 1974; Gautron and Gisiger 1976; Gisiger et al. 1978), as well as the present one, have demonstrated that in the SCG, which contains the A12 form, AChE cytochemical reaction is present throughout the entire Golgi apparatus. This, however, was not the case for the pancreatic acinar cells lacking the A12 form, in which only the TGN appeared labeled. The absence of the collagen tail subunit of AChE in acinar cells may well be related to the lack of cytochemical signal in the Golgi of the acinar cells, because no assembling of the A12 form is required in these cells.
That some cells, but not all, showed involvement in the synthesis of AChE may indicate that this cellular activity could be cyclic. The mosaic-like pattern detected for AChE in RER reflects differences in amounts of AChE among cells as related to the sensitivity of the technique and to the threshold levels of detection. A similar mosaic-like pattern of AChE was also reported for neural and muscle tissues (Gautron and Gisiger 1976; Gisiger et al. 1978; Gisiger and Stephens 1988). For the pancreas, differences in amounts of cellular AChE may be related to modulation of the response to cholinergic stimulation and may explain, at least to some extent, the non-homogeneous secretory response of digestive enzymes to stimulation (Phaneuf et al. 1985).
As to the AChE signals obtained by enzyme cytochemistry and immunocytochemistry in the extracellular space, particularly those associated with the basement membranes and the collagen fibers, they are consistent with the literature showing interactions between AChE and various basement membrane components, including perlecan (Vigny et al. 1983; Brandan et al. 1985; Nicolet et al. 1987; Ramirez et al. 1990; Peng et al. 1999).
Recent progress has underlined the essential role of cholinergic receptors located on the acinar cells in the regulation of exocrine pancreatic secretion (Gardner and Jensen 1993; Holst 1993; Singer 1993; Shetzline and Liddle 1999). As described for the central nervous system (Descarries et al. 1997), acetylcholine released by varicosities of the interacinar nerves must be delivered to the cholinergic receptors as concentric waves of transmitters diffusing throughout the glandular parenchyma. One essential role of AChE in the pancreas should be to keep the ambient concentration of acetyl-choline resulting from this diffuse transmission as low as possible, to minimize receptor desensitization and thereby maintain the efficiency of the cholinergic regulation of secretion. Alternatively, the new non-neural localization of AChE reported here, together with the many already known non-neural ones (e.g., erythro-cytes, endothelial cells) provides significant support for non-cholinergic functions of AChE (Massoulié et al. 1993). Our data suggest that two sources of AChE are available to the acinar parenchyma, AChE present in the extracellular space surrounding them, most probably as a result of the well-established AChE secretion by the neurons (Gisiger and Vigny 1977; Brimijoin 1983; Massoulié et al. 1993), and AChE produced and externalized by the acinar cells themselves. In this respect, the AChE present in the tubular lysosomal network of the acinar cells may stem from either or both origins. It has been postulated that the tubular lysosomal network consists of an endosomal–lysosomal system that could be involved in the uptake and degradation of endocytosed material, because it sequesters fluid-phase soluble tracers present in the interstitial space and thus could be involved in receptor-ligand processing (Oliver 1982; Beaudoin et al. 1991; Alvares 1994). In such a case, our tubular network might be more related to the endosomal compartment than to the lysosomal one. On the other hand, studies performed on these acinar cells and on other cell types have, in turn, assigned an exocytotic function to the tubular network (Kobayashi and Robinson 1991; Grondin and Beaudoin 1996). The presence of AChE within the RER cisternae, reflecting synthetic activity, together with the previous indication that the tubular network in acinar cells can be involved in secretion (Grondin and Beaudoin 1996), may indicate that the cell, probably through its tubular system, would be able to secrete this enzyme at its basolateral pole. Work performed on GP2 in acinar cells also supports the possible involvement of the tubular lysosomes in the exocytotic activity of the cell (Beaudoin et al. 1991; Grondin and Beaudoin 1996). In this respect, there could be some similarities with the coexistence of ortho- and retrograde axonal transport of AChE in neurons (Lubinska 1964; Brimijoin 1983). The methods used in the present study were, however, unable to enlighten us on the movement of the AChE in these tubular structures. Either way, we can assume that the presence of such an enzyme in the acinar cells is a good indication that the cells can regulate and modulate the cholinergic stimulation to which they are submitted.
Footnotes
Acknowledgments
Supported by a grant from the Medical Research Council of Canada.
We would like to thank Jean Gautron for advice and direction concerning the cytochemical technique, Jean Massoulié for kindly providing the specific antibody against AChE, and Bernard Jasmin for constructive discussion. We are also grateful to Gilles Lesperance for letting us use his high-voltage electron microscope facility. The technical assistance of Diane Gingras is gratefully acknowledged.