
Abstract
Immunodetection of renin–angiotensin system (RAS) components indicates that there is a local RAS in anterior pituitary cells, particularly in lactotropes. We have attempted to determine if RAS molecules are secreted by lactotropes and the secretory pathways and intracellular sites of maturation. We investigated the secretory activity of individual lactotropes, using the reverse hemolytic plaque assay (RHPA), with GH3B6 tumor cells and normal male rat pituitary cells. We also determined the subcellular distributions of RAS components in these cells. Both tumor and normal cells secreted angiotensinogen, prorenin, renin, angiotensin I, angiotensin-converting enzyme, and angiotensin II, although at different levels. The percentage of secretory cells was generally higher in tumor lactotropes than in normal cells. The subcellular distribution of RAS components obtained by immunoperoxidase was very similar in both cell types, although the intensities of immunoreactivity differed. Cleaved and uncleaved components were found in rough endoplasmic reticulum (RER), Golgi saccules, and secretory granules, all compartments of the secretory pathway. The cleaved components in the RER suggest the existence of early maturation, whereas the presence of uncleaved products in the secretory granules of normal lactotropes might indicate late maturation sites.
Keywords
T
We first investigated the secretion of RAS components by individual cells. The capacity of pituitary cells to secrete AGT or A II has been investigated in hemipituitaries and in cell cultures or aggregates by direct or indirect measurements of these components in perfusion or culture media (Deschepper 1991; Deschepper et al. 1986; Robberecht et al. 1992; Sernia et al. 1997). However, these measurements did not provide any data on the secretion by individual specialized pituitary cells.
The reverse hemolytic plaque assay (RHPA) has been used with pituitary cells (Neill and Frawley 1983; Smith et al. 1986; Neill et al. 1987; Vila–Porcile and Barret 1996) to demonstrate and quantify the secretory activity of individual cells for a given component and from a given cell type when coupled to immunocytochemistry. The secretion of AGT by gonadotropes was detected by this method (Sernia et al. 1992).
Because our previous study on the distribution of AGT, prorenin, and renin in the secretory granules of pituitary cells showed that most of these components were in lactotropes (Vila–Porcile and Corvol 1998), we have focused on this cell type. We measured the secretion of AGT, prorenin, renin, AI, ACE, and A II by individual lactotropes and determined the subcellular distributions of these components.
We used two models of lactotropes: a tumor rat cell line, GH3B6 cells, that produce mainly prolactin (Gourdji et al. 1972), and normal cells dispersed from male rat pituitaries and briefly cultured. The lactotropes in the pituitary preparations were identified immunocytochemically after each assay, to determine the percentage of secretors in a given cell population.
Both types of cells appeared to secrete all the components of the RAS. The tumor cell line had the highest percentage of secretors for most components. The RAS components were found in the same subcellular compartments of the secretory pathways in tumor and normal lactotropes: the cisternae of the rough endoplasmic reticulum (RER), Golgi apparatus, and secretory granules, in which all the components were detected. The intensities of the immunoreactions varied with the component in both tumor and normal cells. These findings are consistent with the results obtained by RHPA on the capacity of lactotropes to secrete RAS components. However, they do not indicate the precise intracellular steps of maturation of the active forms.
Materials and Methods
Antisera
The sources and specificities of the antibodies used in this study are summarized in Table 1. The specificities of these antibodies were previously established by the corresponding authors.
Antibodies used in study
aDilutions in parentheses were used for RHPA; dilutions not in parentheses were used for immunocytochemistry.
Light microscopic pictures of living GH3B6 cells after a 4-hr RHPA. (
Clonal Pituitary GH3B6 Cells
This rat tumor pituitary cell line, which secretes large amounts of prolactin (Gourdji et al. 1972), is currently cultured in our laboratory and was used because these cells behave in many ways as normal lactotropes.
RHPA
This technique was applied to pituitary cells as early as 1983 by Neill and Frawley. A detailed description of our methodology is given elsewhere (Vila–Porcile 1994; Vila–Porcile et al. 1994; Vila–Porcile and Barret 1996). The technique was slightly modified to study the secretion of RAS components. RHPA requires the use of reactive but non-cytolytic antibodies (Neill et al. 1987; Vila–Porcile 1994).
GH3B6 cells were seeded in 25-cm2 plastic flasks (Falcon; 2.106 cells/flask) in Ham's F12 medium (Gibco, Grand Island, NY) supplemented with 15% horse serum, 2.5% fetal calf serum, 1 nM E2, 10 nM EGF, 180 nM insulin, and antibiotics, and were grown for 7 days. They were harvested by a short (2-min) trypsinization, rinsed three times with Ham's F12 medium supplemented with 0.1% bovine serum albumin (henceforth referred to as F12/BSA), and mixed with an equal volume of ovine red blood cells (RBCs; Sanofi-Pasteur Diagnostics, Marnes-la-Coquette, France). The RBCs had been previously coated with protein A (Sigma; St Louis, MO) by the method of Gronowicz et al. (1976), modified by Neill and Frawley (1983). The mixture of cells (5.105 cells/ml) and RBCs (20% in F12/BSA) was infused into Cunningham chambers constructed on polylysinecoated glass slides (Cunningham and Szenberg 1968; Neill et al. 1987; Smith et al. 1986). The cell mixtures were first incubated in F12/BSA for 1 hr in a humidified incubator (37C, 5% CO2) for attachment of cells and RBCs; unattached cells and RBCs were removed by washing with F12/BSA. The cell mixture was then incubated with specific antibodies against RAS components (see Table 1 for specificities and dilutions) for 4 hr at 37C. The resulting plaques were developed with undiluted guinea pig complement (Sanofi-Pasteur Diagnostics) for 2 hr at 37C. Longer incubations (15–22 hr) increased the apparent number of plaques but led to diffuse hemolysis that made counting difficult. GH3B6 cells were tested for the secretion of AGT, prorenin, renin, A I, ACE, and A II. Two or three chambers were examined for each of these components.
Controls were obtained by (a) omitting the specific antibodies, (b) omitting the complement, or (c) incubation with rabbit pre-immune serum instead of the specific antibody. In all three cases, no plaques were detected.
A positive control of prolactin secretion was included in each experiment to check the cell conditions and reactivity to the assay.
Plaques were systematically photographed under a phase-contrast microscope and then fixed with 2% glutaraldehyde in cacodylate buffer, supplemented with 100 μl toluidine blue/ml to enhance the contrast of non-plaque-forming cells. Plaques were counted and measured on fixed cells. At least five microscopic fields were counted, giving an average of 460 cells per chamber checked for plaque formation. The number of plaques was corrected for the number of cells in the chamber. Plaque diameters were measured with an ocular micrometer to calculate the plaque areas.
Data from six different experiments were analysed statistically by ANOVA, followed by the Fischer's and Scheffé's tests; p >0.01 was considered significant.
Immunoelectron Microscopy. GH3B6 cells were seeded in 35-mm plastic Petri dishes (Falcon, Becton Dickinson Europe, Heidelberg, Germany; 1.105 cells/dish) and cultured for 7 days in Ham's F12 medium supplemented as indicated above.
Immunoelectron microscopic detections of RAS components were performed directly in culture dishes that had been rinsed with F12 alone to eliminate traces of serum. The peroxidase pre-embedding technique was used (Tougard et al. 1980; Tougard and Picart 1986). Briefly, cells were fixed with a periodate-lysine-paraformaldehyde mixture (McLean and Nakane 1974). Specific antibodies against RAS components (see Table 1) were incubated with the cells at RT for 90 min. Cells were not pretreated to remove endogenous peroxidase, because we found no change in the results after using the method proposed by Straus (1971). Cells were also incubated with peroxidase-conjugated complementary antibodies at RT for 90 min. Cells were embedded in situ in Epon (Picart and Tixier–Vidal 1974; Vila–Porcile et al. 1992). Ultrathin sections from selected areas were cut parallel to the embedded cell monolayer (Picart and Tixier–Vidal 1974). Sections were examined under a LEO 906 electron microscope (Oberkochen, Germany) without any further staining.
Normal Pituitary Cells
The pituitaries were removed from normal adult male Wistar rats (180–200 g; Iffa-Credo, L'Arbresle, France), under aseptic conditions and placed in a sterile vessel containing Ham's F12 medium. They were immediately dissected and the neurointermediate lobes were discarded.
Pituitary cells destined for RHPA and primary cultures were dispersed enzymatically and mechanically (Tixier–Vidal et al. 1973). Normal cells destined for RHPA were cultured overnight in large dishes for bacteriology (Falcon; 2.106 cells/dish) in F12 medium supplemented with serum (10% horse serum, 2.5% fetal calf serum) to allow the cells to recover after trypsinization. Primary cultures were prepared by plating the dispersed cells (4.105 cells/dish) in 35-mm plastic Petri dishes in the same medium.
Some whole dissected pituitaries were fixed for 1 hr with 4% paraformaldehyde in SÖrensen buffer, minced into small pieces, fixed for a further hour in fresh fixative, and embedded in LR White (LRW; Ladd Research Industries, Burlington, VT) (Ozawa et al. 1994; Vila-Porcile and Corvol 1998).
RHPA results
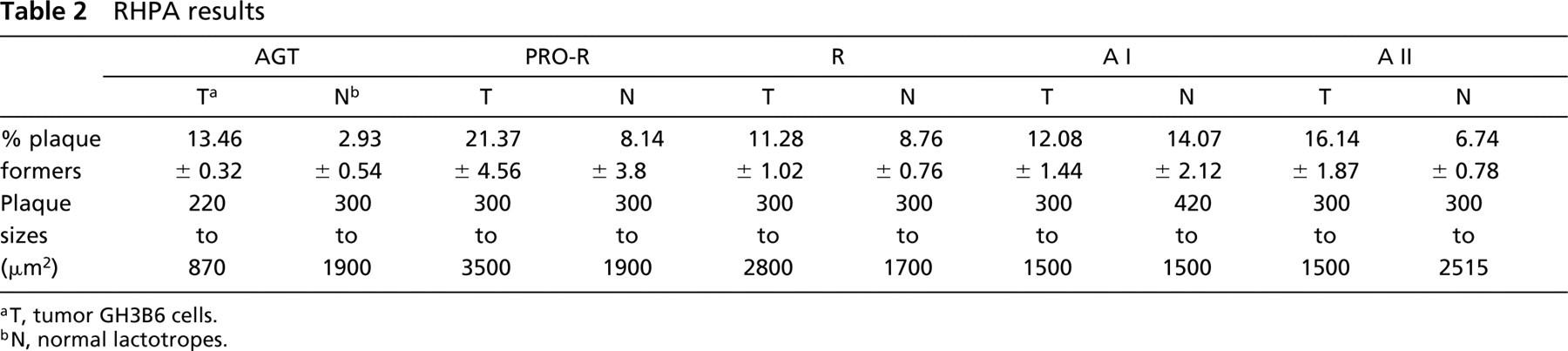
aT, tumor GH3B6 cells.
bN, normal lactotropes.
(
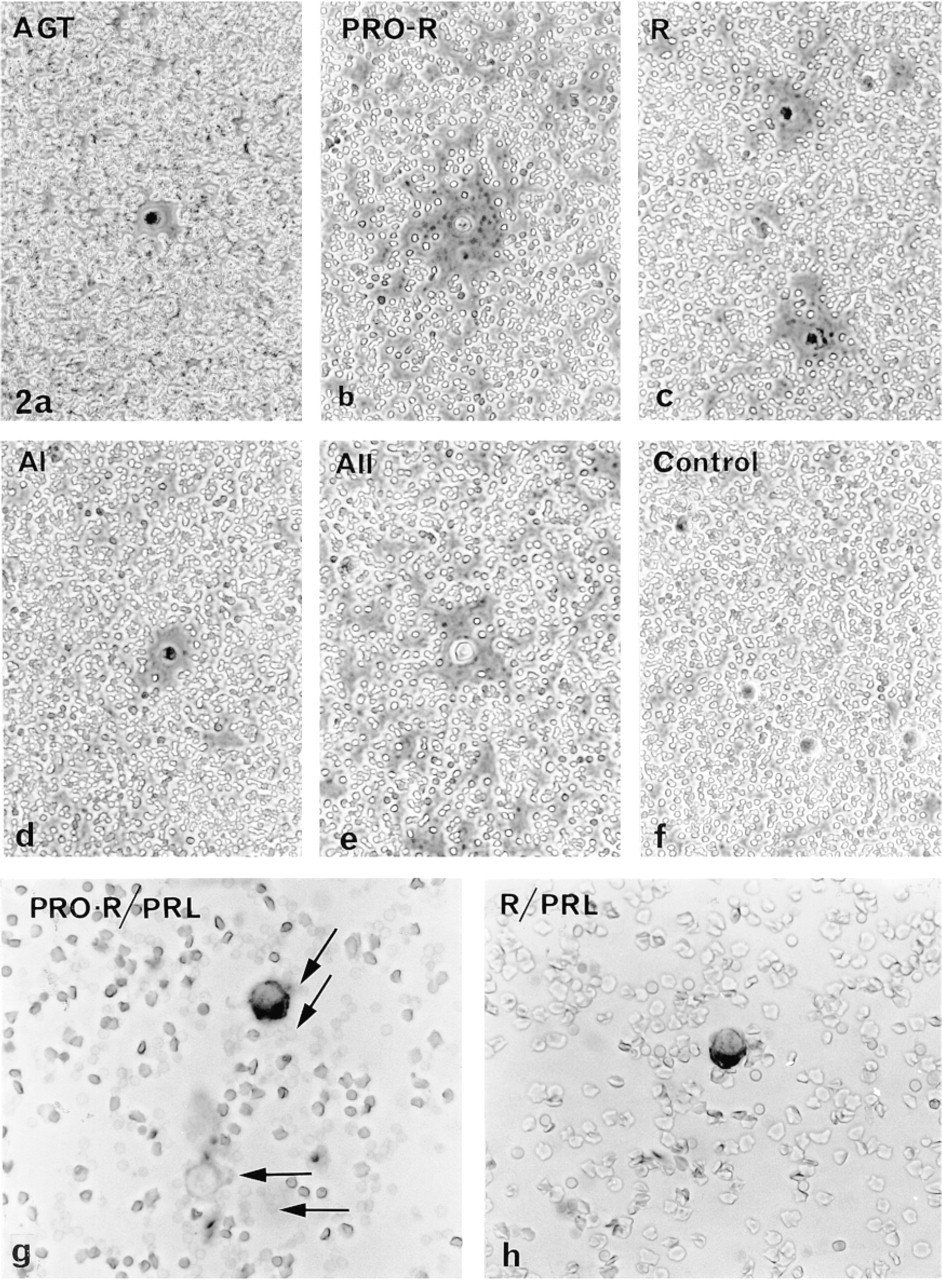
RHPA. Precultured cells were harvested by gentle pipetting, rinsed twice with F12/BSA, adjusted to 106 cells/ml with F12/BSA, mixed with protein A-coated RBCs, and processed for RHPA. At the end of the plaque development (2 hr), plaques were photographed under the phase-contrast microscope. The chambers were then infused with 0.2 M sodium acetate/0.5 M NaCl for 10 min at 4C to elute antibodies bound to the RBCs (Smith et al. 1986). The plaques were fixed overnight at 4C with a slightly modified B5 fixative [(0.6 g HgCl2, 0.125 g sodium acetate, 9 ml distilled water, and 1 ml 20% formaldehyde (Ladd Research Industries) added extemporaneously)]. The RAS components studied were AGT, prorenin, renin, ACE, A I, and A II.
Lactotropes were identified immunocytochemically. Chambers were successively infused with ethanol 95% (1 min), Lugol (1 min), distilled water (1 min), 2% sodium thiosulfate in 0.05 M Tris-HCl buffer, pH 7.4 (1 min), buffer (1 min), 3% H2O2 in distilled water (2 min), buffer (three times), 3% BSA in buffer (30 min), and buffer supplemented with 1% BSA plus 0.3% saponin (permeabilizing agent; 30 min). This detergent-supplemented buffer was used thereafter to dilute the antibodies and to wash the chambers. The cells were then incubated with anti-prolactin (PRL) antibody (diluted 1:100–1:200) for 90 min at RT, rinsed three times for 10 min, and incubated with peroxidase-conjugated complementary antibody (Sanofi-Pasteur Diagnostics; diluted 1:100) for 1 hr at RT. The cells were rinsed with buffer (10 min) and immunoreactions were revealed with DAB: three rinses with 0.5 M Tris-maleate buffer, pH 7.6, infusion of DAB (0.4 mg/ml) plus 0.03% H2O2 as an oxygen donor in Tris-maleate buffer (5 min). The stained cells were rinsed once with buffer and twice with distilled water and the chambers cautiously infused with Mowiol (Hoechst).
Hemolytic plaques were generally small (300–2500 μm2), and consequently they were counted and measured under the light microscope using a ×40 objective, to detect even a single row of RBC ghosts around secreting cells (Childs and Burke 1987). Two ratios were calculated from these counts. The number of plaques formed by cells identified as lactotropes was divided by the total number of labeled cells to obtain the percentage of secreting lactotropes. The total number of plaques was divided by the total number of cells (labeled plus unlabeled elements) to obtain a profile of the overall secretion of each component explored. Five to six fields and 200 cells at least (average 460 cells/chamber) were counted in each chamber (six experiments performed, with two to three chambers per component). Statistics were managed as for GH3B6 cells.
Immunoelectron Microscopic Study of Pituitary Primary Cultures. Dispersed pituitary cells were cultured for 7 days in Ham's F12 medium supplemented with 10% horse serum, 2.5% fetal calf serum, and antibiotics. They were thoroughly washed before pre-embedding immunocytochemical detection of RAS components and embedding. The same components (AGT, prorenin, renin, A I, ACE, A II) were revealed, sometimes using several specific antibodies for a given component. A supplementary step was required to identify lactotropes because of the cellular heterogeneity of pituitary cell cultures. Immunogold postembedding was therefore used for ultrathin sections from selected areas of the Epon-embedded cultures. Because Epon embedding generally hinders immunocytochemical reactions, incubations with the specific anti-PRL antibody were extended to 15 hr at 4C (diluted 1:100–1:200) and to 2 hr at RT for the conjugated protein A–colloidal gold (Biocell, Cardiff, UK; diluted 1:20). Sections were examined without further staining. A total of 20 to 40 cells were examined for each component.
Immunoelectron Microscopic Study of Intact Pituitary Tissue. Three components, A I, ACE, and A II were localized in lactotropes by postembedding immunogold on ultrathin sections of pituitary tissue embedded in LRW, by double labeling (Bendayan 1982) as previously reported (Vila–Porcile and Corvol 1998) for other RAS components. The distributions of antibodies against these RAS components (15–24 hr at 4C) were revealed with large-sized gold particles. Cells were incubated with an anti-PRL antibody for 1–2 hr at RT, which was revealed with small-sized immunogold particles.
Results
Secretion of RAS Components: RHPA Results
The RHPA study of the secretion of several RAS components by individual pituitary cells demonstrated that both GH3B6 cells and normal cells, particularly lactotropes, released these components.
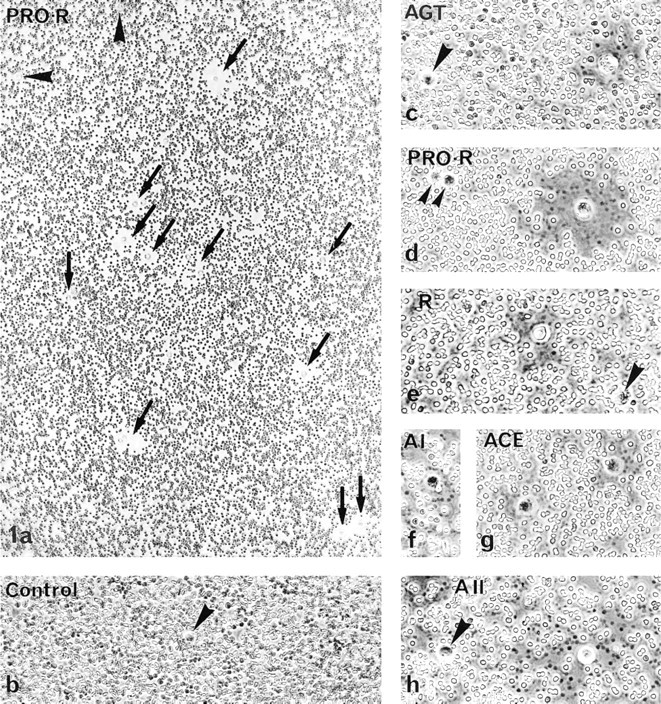
GH3B6 cells actively secreted all of the constituents tested: the precursor (AGT), the two inactive products, prorenin and A I, the cleavage enzyme ACE, and the two active components, renin and A II (Figure 1). The diameter of the plaques varied with the component and with the cell sample for a given component (see Table 2). As a consequence of this variability, the differences between the mean areas were not statistically significant. Because only some GH3B6 cells were active, we determined the percentage of secreting cells in fixed preparations. We obtained significant differences between mean values, thus permitting comparisons between RAS components and between tumor and normal cells (see below; Figure 4). More than 20% of the GH3B6 formed hemolytic plaques for prorenin, and only 12–16% did so for AGT, AI or AII (see below; Figure 4). The percentage of plaque formers could not be determined for ACE in these experiments owing to poor plaque fixation after the assay with the specific Y4 anti-ACE antibody.
Normal pituitary cells, taken as a whole population, released all the components studied (AGT, prorenin, renin, A I, ACE, and A II). Well-formed but rather small hemolytic plaques were seen around living cells under the phase-contrast microscope (Figure 2). As in GH3B6 cells, the plaque diameters varied with the component and the cell sampling, precluding any statistical comparison of the plaque areas. We therefore focused on the percentages of secretors (see Table 2). About 4% of the cells secreted A I and 8% of them released prorenin. The greatest percentages were for prorenin and A II (Figure 3).
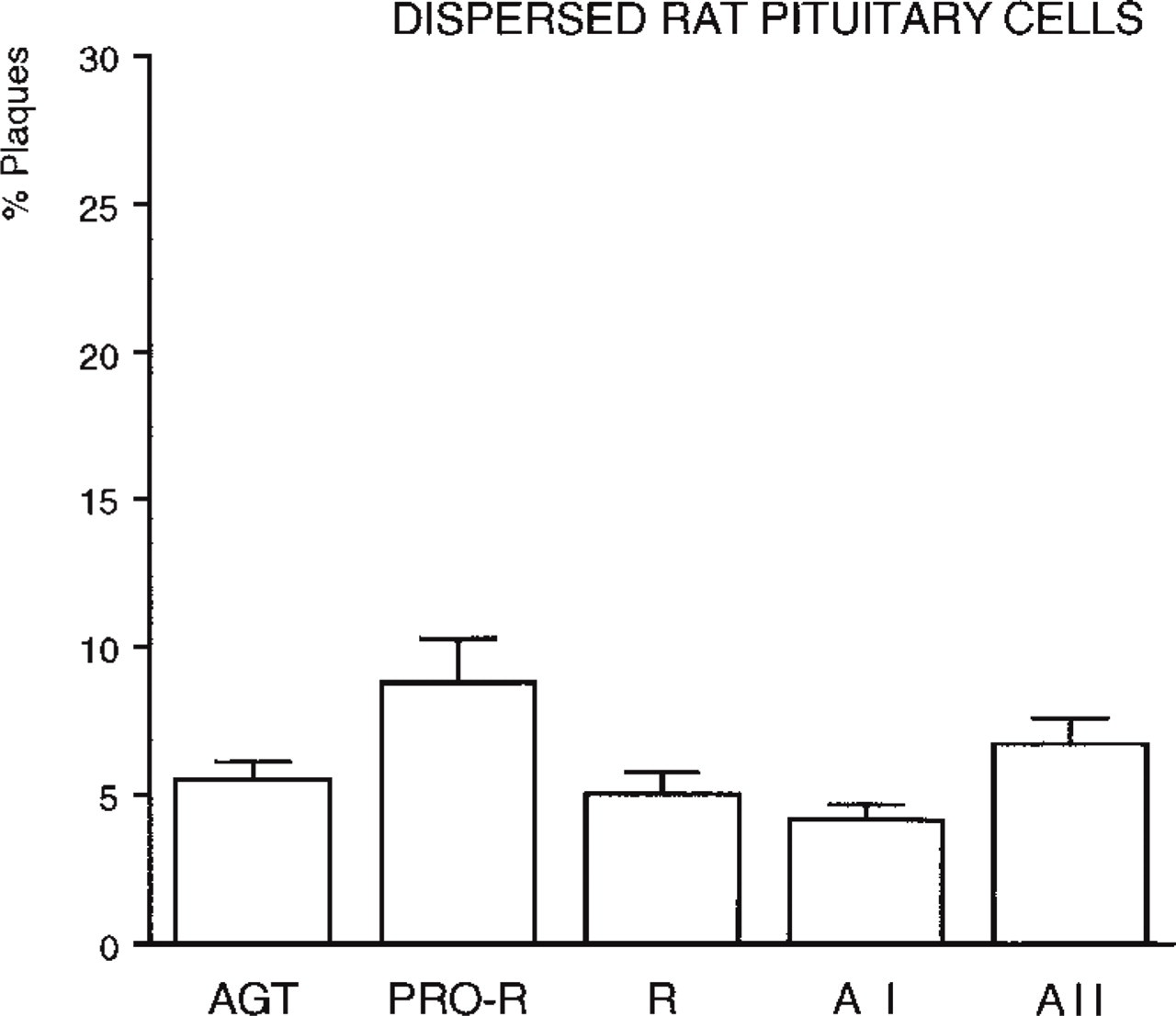
Percentages of hemolytic plaques formed by the total population of normal pituitary cells cultured for 15 hr, after a 4-hr RHPA performed with different antibodies raised against RAS components: AGT (polyclonal HCL antibody); PRO-R, prorenin (polyclonal 657-2 antibody); R, renin (polyclonal BR 1-4 antibody); A I (polyclonal BA I 02 antibody); A II (polyclonal BA II 01 antibody). For each component, the ratio of the total plaque number to the total number of cells counted in the chambers (mean cell count 460 cells/chamber) was established from the accumulated results of six different experiments. Such a graph provides a profile of the global secretory activity of all pituitary cells, and is to be compared to Figure 4 (see text). Values were significantly different (p >0.01) for AGT vs PRO-R and for PRO-R vs R.
We used immunocytochemical identification of lactotropes to compare the number of secretors in this subpopulation to that in the whole pituitary cells (Figure 3 vs Figure 4). The percentages of prorenin and A II secretors were almost similar in the two cases. In contrast, the percentage of AGT-secreting lactotropes was lower and those of renin- and A I-secreting lactotropes were higher. Because immunocytochemically identified lactotropes accounted for approximately 30% of the total dispersed cells, they were the main source of prorenin and A II released by the total population.
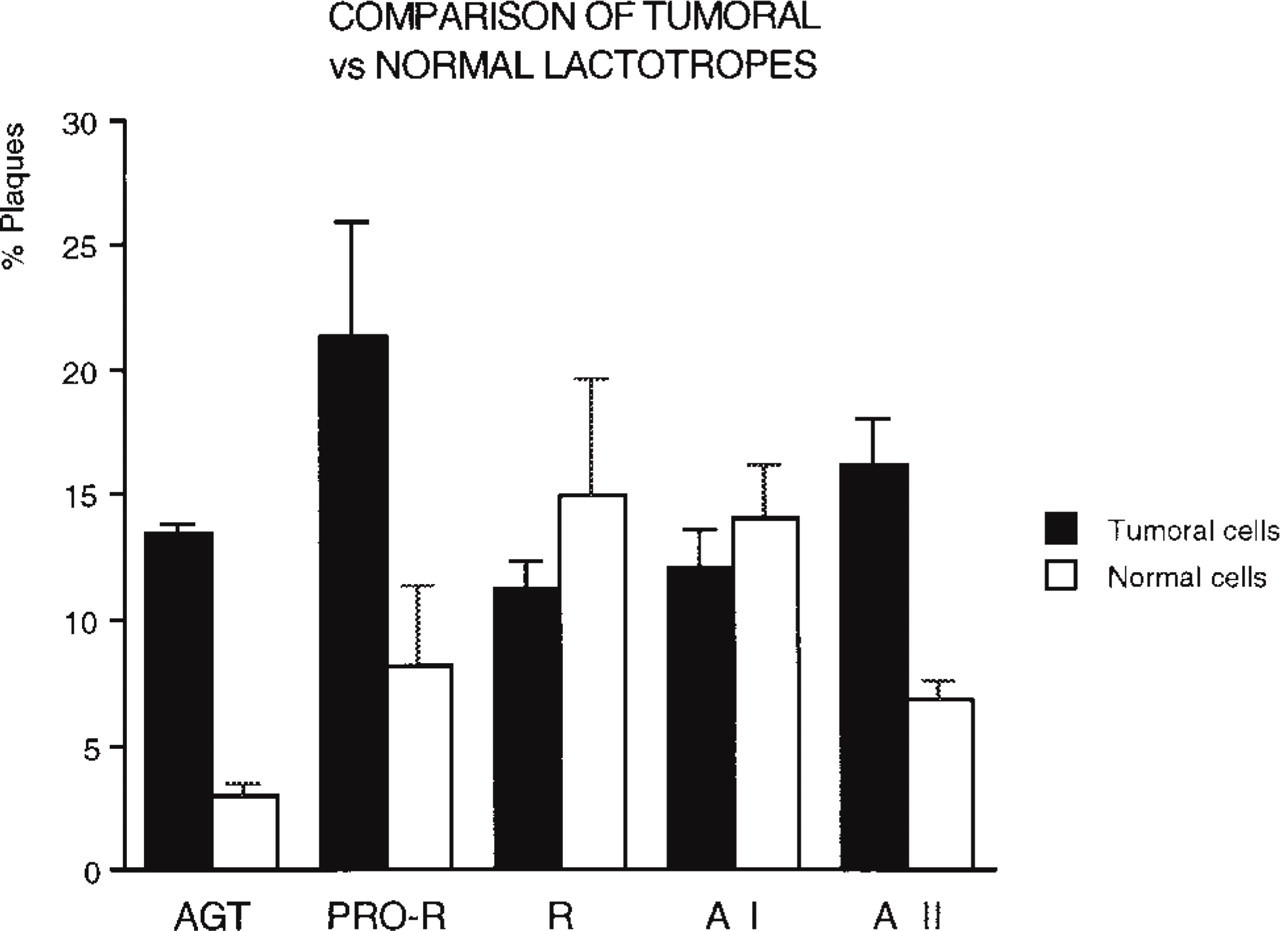
Comparative percentages of hemolytic plaques formed by tumor GH3B6 cells and by normal pituitary lactotropes immunocytochemically identified after a 4-hr RHPA performed on the two cell populations with the same antibodies as in Figure 3 (accumulated results; see text). Values obtained from tumor vs normal cells were significantly different (p >0.01) for AGT, PRO-R, and A II. Among tumor cells, differences are significant (p >0.01) for PRO-R vs R and vs A I, for R vs A II, and for AI vs AII. Among normal cells, significant differences are noted for AGT vs all the other components. At least 200 cells/chamber were checked for plaque formation by normal lactotropes.
The percentages of secretors from GH3B6 cells and from normal lactotropes (Figure 4) indicated that a larger proportion of GH3B6 cells secreted AGT, prorenin, and A II.
Ultrastructural Localization of RAS Components in Tumor and Normal Cultured Cells: Results of Immunoperoxidase Pre-embedding
Because normal and pituitary tumor cells appeared able to secrete both cleaved and uncleaved products, their precise subcellular localizations were investigated with the immunoperoxidase pre-embedding method to obtain information about the maturation pathways of these RAS constituents in normal and tumor lactotropes.
All the RAS components were detected in the sub-cellular compartments of the secretory pathway in all the cells examined: RER, Golgi apparatus, immature and mature secretory granules, and occasionally the lysosomes. However, the intensity of the immunoreaction for each constituent in a given compartment differed greatly (Figures 5 and 6; Table 3).
The most prominent labeling was obtained for prorenin in both GH3B6 cells and normal lactotropes. It was mainly in the Golgi apparatus, on the luminal side of the saccules and vesicles, and on forming, immature (Figure 5b and 6a) and mature secretory granules. Prorenin was also detected in some cisternae of the RER (Figure 5b and 6a). The polyclonal BR 1-4 and monoclonal 4G1 antibodies detected renin mostly in the Golgi (Figure 5c and 6b), in some vesicles, and in some cisternae of the RER (Figure 5c). AGT (using either ORNI or HCl antibodies), ACE, and A II were detected in some RER cisternae of both normal and tumor lactotropes (Figure 5a, 5g, and 6d), suggesting that they coexist in this compartment. Labeling for A I was discrete within the sub-cellular compartments, even in the Golgi apparatus of tumor (Figure 5d) and normal (Figure 6c) cells. There was a strong immunoreaction to ACE in the mature and immature granules of both cells (Figure 5e and 6d) and, as expected, labeling of the plasma membrane (Figure 5d). A II was found in the RER with the monoclonal 4D8 (Figure 5g), while the Golgi saccules and forming secretory granules were discretely labeled with the polyclonal antibody BA II-01 (Figure 5f and 6e).
Ultrastructural Distribution of RAS Components in Intact Pituitary Tissue: Immunogold Results
A I, ACE, and A II were detected on ultrathin sections of LR White-embedded normal male rat pituitary tissue by double postembedding labeling (Figure 7). There were large gold particles at the periphery of the matrix of mature secretory granules in cells identified as lactotropes with smaller particles. Therefore, both the precursor A I, its cleavage enzyme ACE, and the mature product A II were present in the storage compartment, the last site on the secretory pathway, just before release.
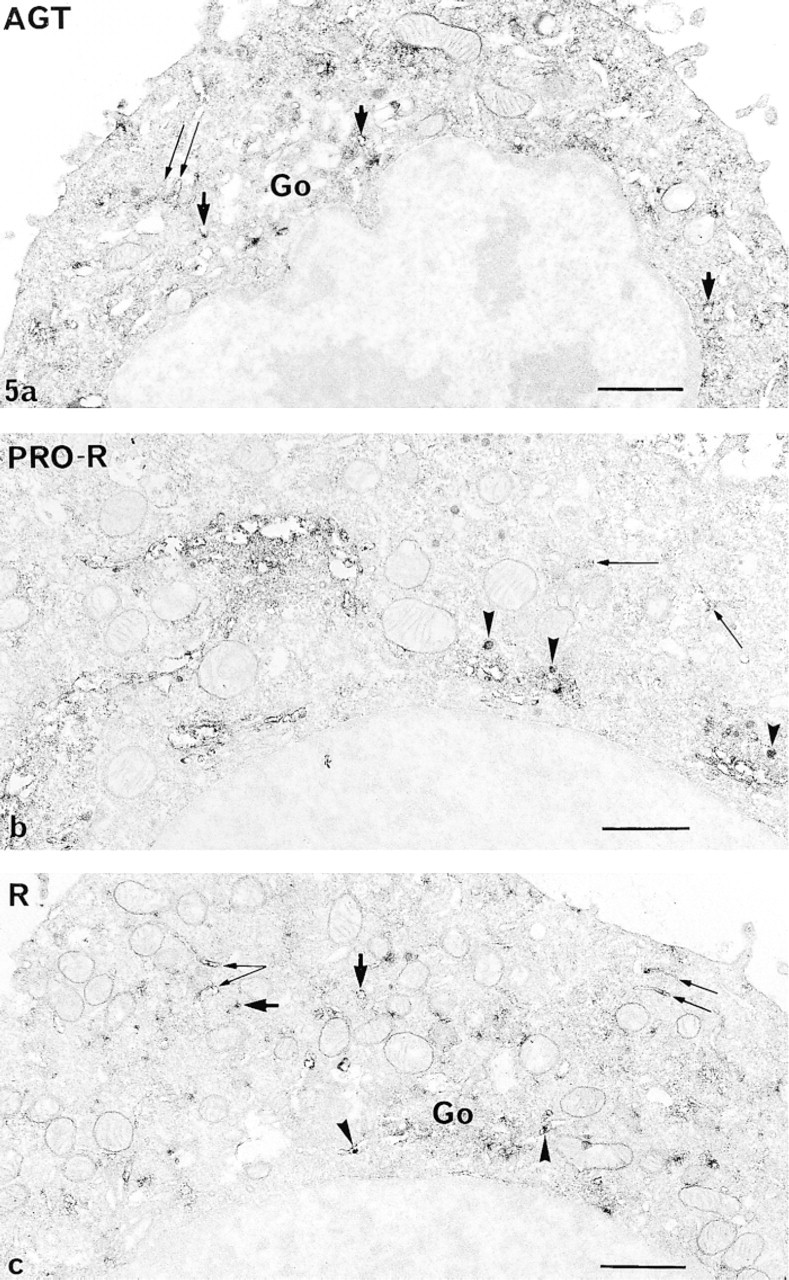
Utrastructura distribution of the immunoreactions to anti-RAS antibodies, as shown in GH3B6 cells by the peroxidase pre-embedding technique. (
Discussion
The present work was done to answer two questions.
(a) Are prolactin cells, which contain RAS components, able to release all or some of these components?
(b) Where do the active forms of the RAS components, renin and A II, mature?
We used RHPA and immunocytochemistry to show that normal rat pituitary cells, particularly lactototropes, and the tumor PRL-producing GH3B6 cells, are able to secrete RAS components, including the cleavage enzymes, as active or inactive forms, although at different levels. All of these components were also detected by immunoelectron microscopy in all the subcellular compartments of the secretory pathway in normal cells and in GH3B6 cells, although with variable labeling intensities.
However, the secretory activities of the normal and tumor cells differ. None of the RAS components studied was ever secreted by all of a cell population. This variable capacity of anterior pituitary cells has been reported for other secretory products (Neill et al. 1987; Vila–Porcile and Barret 1996). We therefore evaluated and compared the percentages of secretors in a given cell population for a given component. We also compared the diameters of the hemolytic plaques, but the most significant differences between the two cell populations were in the percentages of plaque formers.
The percentages of secreting tumor and normal lactotropes varied greatly, depending on the RAS component studied. Comparison of the respective percentages of tumor and normal lactotropes secreting each component (see Figure 4) reveals that a higher percentage of tumor cells secreted AGT and prorenin than normal lactotropes, whereas renin and A I were secreted by about the same percentages of both cells.
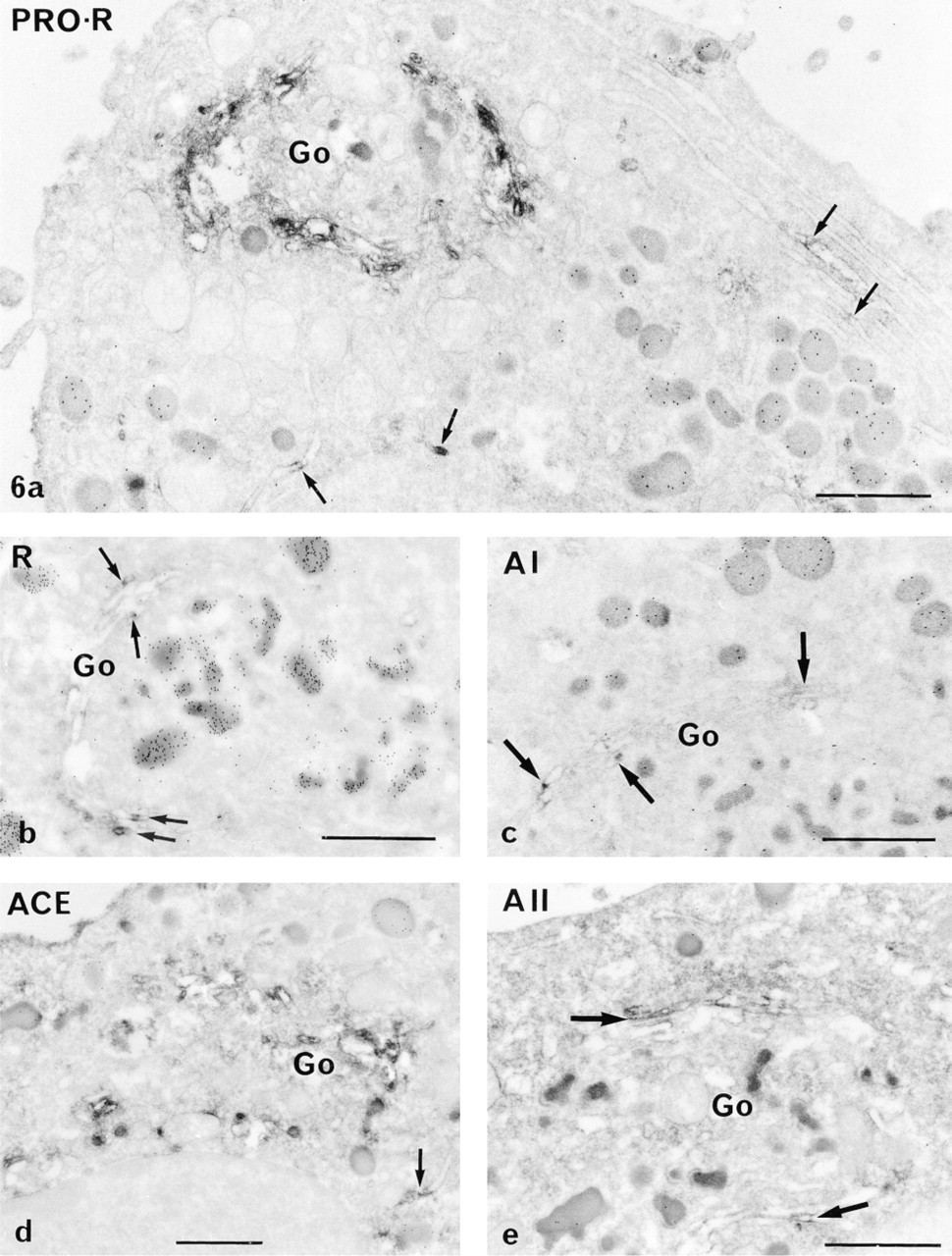
Immunodetection of RAS components in normal pituitary cells, as shown in the Golgi area by the peroxidase pre-embedding technique and the use of specific anti-RAS antibodies. Cells were identified as lactotropes by the presence of gold particles on their secretory granules after immunogold postembedding (see Materials and Methods). (
Semiquantitative evaluation of immunoreactions to RAS different components in tumor and normal lactotropes
aAGT, angiotensinogen; PRO-R, prorenin; R, renin; A I, angiotensin I; ACE, angiotensin-converting enzyme; A II, angiotensin II.
bAll the antibodies (see Table 1) were used for both immunocytochemistry and RHPA, except 4D8.
AGT was secreted by 13.5% of the GH3B6 cells but by only 3.4% of normal lactotropes. AGT secretion has been previously investigated using the RHPA approach, but with different anti-AGT antibodies, on dispersed pituitary cells (Sernia et al. 1992; Sernia 1995). The authors concluded that AGT was secreted by gonadotropes and by an undetermined population of small perisinusoidal cells, but not by lactotropes (Sernia et al. 1997). However, we find that normal lactotropes do secrete AGT, although in small numbers. They appear to contribute only a part of the AGT secreted by the total pituitary cell population (see Figure 3). In contrast, normal lactotropes appear to be particularly involved in the secretion of renin and A I (see Figures 4 and 3). The anti-A I antibody used in this work did not crossreact with A II, making it certain that the secretory activity of normal and tumor cells as well as the positive immunoreactions concerned only A I.
The secretion of ACE is less certain. Hemolytic plaques were formed by the two cell populations and were observed around living cells under the phase-contrast microscope, making it certain that the enzyme was released. However, it was difficult to count the plaques because our anti-ACE antibody caused a general hemolysis of the RBCs at fixation. No reliable quantitative results were therefore obtained.
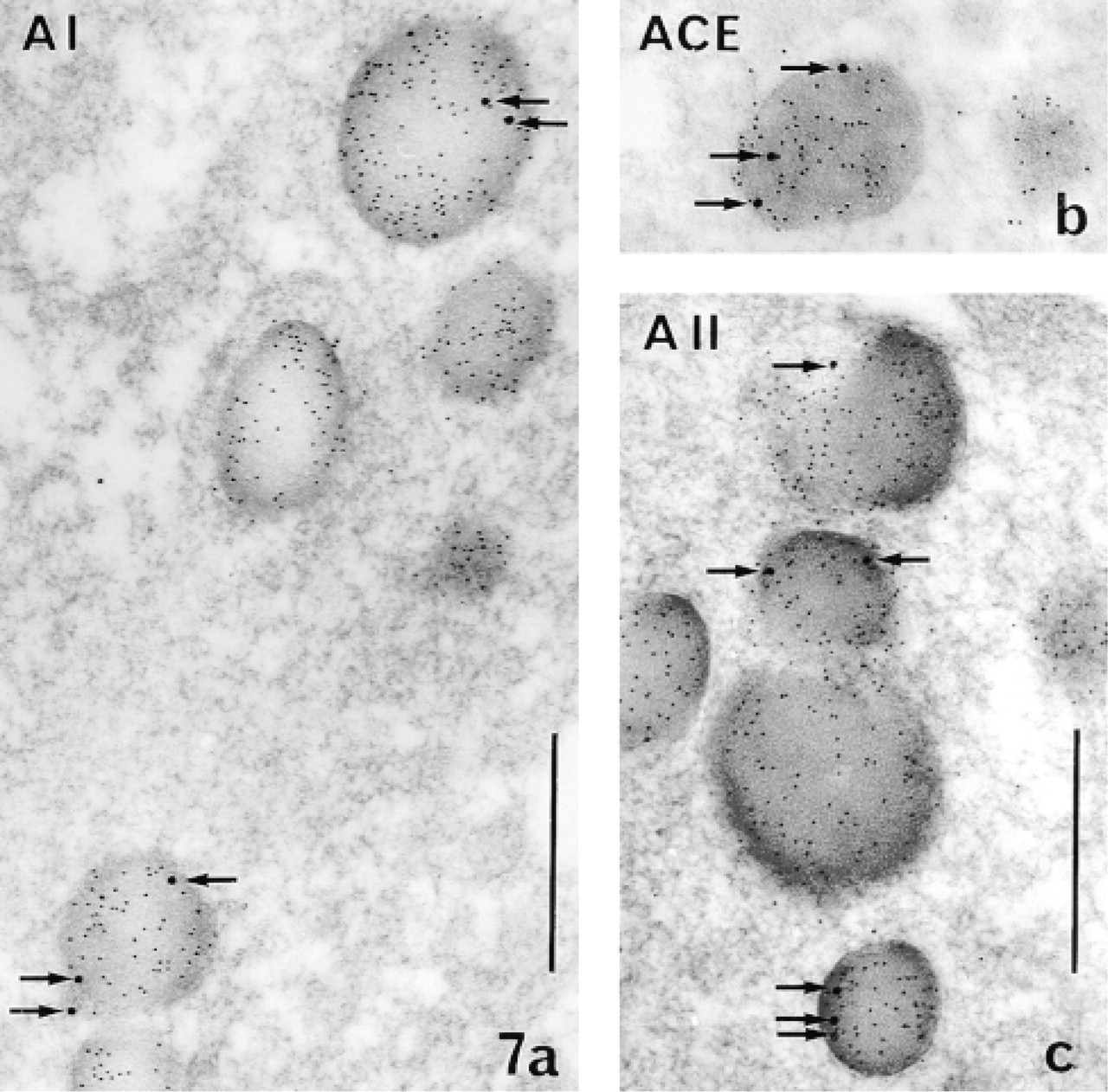
Electron micoscopic immunodetection of A I, ACE, and A II in the secretory granules (storage compartment) of pituitary cells in vivo (double postembedding immunogold technique applied on ultrathin sections of LR White-embedded pituitary tissue). Large gold particles (15 nm, arrows) indicate the presence of the different RAS components. The preferential location of the three components at the periphery of the granules is to be noted. Cells were identified as lactotropes by detection of prolactin with small (5-nm) gold particles, which appear dispersed over the entire granular matrix. (
In contrast to the large differences in their secretory ability revealed by RHPA, the two lactotrope models had many common features in the distribution of the RAS components by immunoelectron microscopy.
The main common feature of GH3B6 cells and normal lactotropes was the very strong labeling for prorenin in the Golgi. Not only the stacks of saccules but also forming and immature secretory granules were densely stained in the Golgi apparatus, thus suggesting either a blockade at this level or oversynthesis of the proenzyme. The fact that many cells that secreted prorenin had intensely labeled Golgi apparatus under the light microscope (not shown) argues in favor of the later.
Both normal and tumor cells had discrete labeling of A I and A II only in RER cisternae, Golgi area, and secretory granules, unlike prorenin, AGT, and renin, although RHPA indicated considerable release of these components, except AGT, by normal lactotropes.
ACE was sometimes found in the Golgi zone of a small number of cells. This does not support further speculation, but the labeling of ACE in secretory granules suggests late maturation.
All of the tested RAS components therefore appear to be secreted by normal and tumor lactotropes, and their subcellular distributions have many common features. However, the GH3B6 cells had more plaque formers than the normal lactotropes for most of these components. This may be because of the poor development of the storage compartment in these cells. GH3B6 cells contained only few small secretory granules even when cultured in a medium supplemented with EGF and hormones (see Materials and Methods) (Figure 5). Such a small storage compartment has been linked to a high basal release of their major hormone product, prolactin, and to a more rapid intracellular transport of the newly synthesized hormone (Morin et al. 1984; Tougard and Tixier–Vidal 1994). This feature of GH3B6 cells may explain the high percentage of cells releasing RAS precursors.
Our findings for the subcellular sites of maturation of RAS components are compatible with both late and early maturation. The immunocytochemical localization of the RAS components in secretory granules of normal lactotropes suggests that these components follow at least part of the so-called regulated pathway and suggests the existence of late maturation. This is supported by the detection with immunogold of A I, A II, and the cleavage enzyme ACE in the same secretory granules of normal lactotropes in vivo, which were shown to also contain AGT (Vila–Porcile and Corvol 1998). These data are consistent with the hypothesis of a channeling of AGT through the regulated secretory pathway in lactotropes. However, there may also be a constitutive route for RAS components. The faint labeling for AGT obtained with immunoperoxidase in both tumor and normal lactotropes might be due to rapid cleavage of the precursor, because immunocytochemistry revealed renin, ACE, and AII in RER cisternae and in some Golgi saccules. This suggests early maturation, at least for some of the AGT molecules, with a bypass of the storage or even the Golgi compartments.
The intracellular RAS components we detected might have been recaptured as precursors by pituitary cells, where they could follow the endocytic pathway and complete their maturation. This uptake was debated for AGT (Sernia et al. 1992, 1997). Whatever the importance of this phenomenon, the detection of AGT mRNA in GH3 cells by RT-PCR (Sernia 1995) strongly supports the synthesis of AGT by these cells and is consistent with our present hypotheses.
These results raise the question of the functional significance of RAS components secreted by lactotropes. The most likely function is autocrine regulation of PRL-secreting cells because of the important role played by the final product of the RAS cascade, A II, in the release of prolactin (Steele et al. 1981; see reviews in Ganong 1993, 1994, 1995). Paracrine interactions with other cell types in the intact pituitary and in primary cultures (Steele and Myers 1990; Vankelecom and Denef 1997), as well as the actual role of AII in these interactions, have been questioned (Houben and Denef 1991; Robberecht et al. 1992). Nevertheless, the presence of A II receptors in lactotropes, demonstrated by A II binding (Aguilera et al. 1982) and by RT-PCR (Moreau et al. 1997) on partially purified cells, is a potent argument for an autocrine action. These receptors were also detected by autoradiography in lactotropes morphologically identified by light and electron microscopy (Paglin et al. 1984) and, more recently, they were shown to belong to the AT1B subtype by in situ hybridization on immunocytochemically identified lactotropes in intact pituitary (Lenkei et al. 1999). The responses of lactotropes to A II stimulation was reported to be heterogeneous (Allaerts 1989), owing to the existence of subtypes of lactotropes and also to an unequal distribution of A II receptors and the varying sensitivities of these subtypes to A II.
To our knowledge, this is the first demonstration of the release of other RAS components, particularly the inactive products, by individual pituitary cells, except for reports of AGT secretion by normal gonadotropes (Sernia et al. 1992) and by dispersed pituitary cells under various physiological and experimental conditions (Sernia et al. 1997).
Both tumor and normal lactotropes can release regulatory peptides from the renin–angiotensin system in addition to their normal products, but whether these peptides act to regulate the cells themselves remains to be determined.
Footnotes
Acknowledgements
We thank Dr J. Sealey (657-2, BR 1-4, BA I-02, BA II-01), Dr D. Simon (4G1, 2D12, 4D8), Dr F. Pinet (ORNI), Dr J.M. Gasc (HCL, Y4), and Dr D. Grouselle (anti-PRL) for their generous gifts of antibodies.
We also thank Dr A. Tixier–Vidal for critical and constructive reading of the manuscript and E. Etienne for expert photographic work. The English text was edited by Dr O. Parkes.