
Abstract
Angiotensin II (Ang II) regulates water and sodium transport in renal tubules and gastrointestinal tract. Two types of Ang II receptors have been cloned, but their distributions have not been determined in human colon. In addition, tissue renin-angiotensin systems (RAS) are believed to exist and to regulate local actions in human colon. We studied by reverse transcription-polymerase chain reaction (RT-PCR) and immunohistochemistry (IHC) the presence and localization of Ang II receptors Type 1 (AT1), Type 2 (AT2), and RAS components [angiotensinogen, renin, and angiotensin-converting enzyme (ACE)] in normal human colon. AT1 receptors were localized in vessel walls, myofibroblasts, macrophages, and surface epithelium. AT2 receptors were found in mesenchymal cells and weakly in parts of surface epithelium. Renin and ACE were distributed in vessel walls, mesenchymal cells, and in parts of surface epithelium. Angiotensinogen was also detected by RT-PCR. These findings demonstrated that Ang II receptors and RAS components were present in human colon, suggesting the possibility of its local regulation.
ANGIOTENSIN II (Ang II) has many different actions, most of which are related, either directly or indirectly, to the regulation of blood pressure and of fluid and electrolyte homeostasis. Ang II modulates the transfer of sodium and water across epithelial tissues of the proximal renal tubules and gastrointestinal tract (Bolton et al. 1975; Levens 1985; Cox et al. 1987). Low concentrations of Ang II stimulate sodium and water absorption, whereas high concentrations of Ang II inhibit absorption and/or stimulate secretion (Levens et al. 1981). Two types of Ang II receptors, Type 1 (AT1) and Type 2 (AT2), exist and have been cloned. Recently, Xio-Hong et al. (1999) demonstrated that low concentrations of Ang II stimulate water and sodium absorption via AT2 receptors and high concentrations of Ang II inhibit sodium absorption and/or stimulate sodium secretion via AT1 receptors at the serosal side of the rat jejunum. However, the functions and localizations of these receptors in human colon have not been clearly determined.
In addition to the classical pathway of Ang II synthesis, tissue renin-angiotensin systems (RAS) have been identified in a number of organs, suggesting that various tissues have the ability to synthesize Ang II independently of the circulating RAS (Paul et al. 1993; Phillips et al. 1993). Tissue-specific RAS may be present in the kidney, brain, vasculature, adrenal, heart (Campbell and Habener 1986), and possibly the colon.
To the best of our knowledge, no report is available concerning the localization of AT1 and AT2 receptors and the distribution of RAS components in human colon. We therefore investigate the presence and localization at the mRNA and protein levels of Ang II receptors and RAS components in human colon mucosa.
Materials and Methods
Samples
Two pieces of biopsy specimens were obtained of normal-appearing colon mucosa from patients positive for fecal occult blood. The subjects were 40- to 60-year-old men without hypertension, heart failure, renal disease, sarcoidosis, diabetes mellitus, or inflammatory bowel disease. The research protocol was approved by the Research and Development Committee of the Hamamatsu University School of Medicine.
RT-PCR
Total RNA was purified from biopsy samples by the acid guanidine-phenol-chloroform method (Chomczynski and Sacchi 1987) and was quantified by measurement of absorbance at 260 nm in a spectrophotometer. Total RNA (1.0–5.0 μg) was subjected to first-strand cDNA synthesis using random hexamer primers (Takara Shuzo; Kyoto, Japan) and Superscript II RT (Gibco BRL; Gaithersburg, MD) in a final volume of 20 μl. After incubation at 42C for 50 min, the reaction mixture was treated with RNase H before proceeding to PCR. PCR primers specific for AT1, AT2, angiotensinogen, renin, and ACE cDNA were designed to span at least one intron (Table 1). PCR was performed in a 10 × PCR buffer with 2.5 mmol/liter concentration of each deoxynu-cleoside triphosphate (Takara Shuzo), 0.5 μmol/liter of each sense and antisense primer, and 2.5 U Taq polymerase (Takara Shuzo) in a final volume of 100 μl, using a Zymoreactor AB-1820 (ATTO; Tokyo, Japan). PCR was performed with the hot-start technique to reduce nonspecific priming and thereby enhance the specificity, sensitivity, and reproducibility of the amplification reaction. The PCR cycle consisted of 94C for 1 min, 55C for 1 min (AT1 and renin 55C; AT2 and ACE 53C, and angiotensinogen 57C) and 72C for 1 min; this was repeated for 35 cycles. Control samples for detection of possible PCR contamination were included in all RT-PCR experiments and were routinely negative. The PCR products were separated on 3% (w/v) agarose gels containing ethidium bromide and were visualized with UV light. Some PCR products were ligated into the Thermo sequence II dyterminator cycle sequencing kit (Amersham Pharmacia Biotech; Little Chalfont, UK) and sequenced using an ABI automated DNA sequencing system.
Immunohistochemical Procedures
Other biopsy samples were immediately washed in cold distilled water and fixed for 24 hr by immersion in 4% paraformaldehyde in PBS 0.1 M (pH 7.4) at 4C, and subsequently replaced with 0.1 M PBS (pH 7.4) for 24 hr. Paraformaldehyde-fixed biopsy specimens were embedded in paraffin and cut at 5 μm thickness. Rabbit anti-AT1 receptor polyclonal antibody (sc-1173) and goat anti-AT2 receptor polyclonal antibody (sc-7420) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Mouse anti-ACE monoclonal antibody (CD-143) was purchased from Chemicon International (Temecula, CA). The selectivity of these antibodies has been confirmed in Western blotting analysis as previously described (Nicholas et al. 1995; Miyata et al. 1999; Matsumoto et al. 2000), in which only a single band was detected by each antibody, and there was no crossreactivity of the antibodies against AT1 and AT2 receptors (the manufacturer's information). Rabbit anti-renin antibody was a kind gift of Dr. Fukamizu (Mukai et al. 1984; Fukamizu et al. 1991). A standard biotin-streptavidin-peroxidase method was used. Immunostaining was performed according to routine methods as suggested by the manufacturer (Nichirei; Tokyo, Japan). In brief, sections were deparaffinized and taken to water. The deparaffinated sections were subjected to antigen retrieval in 0.01 M citrate buffer (pH 6.0) by microwaving. Endogenous peroxidase was blocked with 0.3% hydrogen peroxide in methanol for 30 min at room temperature (RT) before washing with PBS, pH 7.4, using a wash-bottle. A blocking reagent, in the form of an appropriate non-immune serum, was applied for 10 min, after which excess reagent was drained from the section. Incubation was performed with the primary antibody overnight at 4C (anti-AT1 and AT2 receptor antibodies were used at a dilution of 1:400; anti-ACE antibody was used at a dilution of 1:200; and anti-renin antibody was used at a dilution of 1:2000 in PBS), and with the secondary biotinylated antibody for 30 min at RT, followed by streptavidin-biotin-peroxidase complex reagent for 10 min at RT. The reaction products were visualized using a color solution that contained 3,3′-diaminobenzidine in a Tris (hydroxymethyl) aminomethane buffer (DAB) (Histofine kit; Nichirei). Sections were lightly counterstained with hematoxylin, dehydrated, and mounted with coverslips. Each step was followed by rinsing of the slide with PBS and draining excess fluid away from the sections. For the immunostaining experiment on AT1 and AT2 receptors, we used preabsorbed sera as a negative control instead of the primary specific antibodies. The antibodies were preabsorbed with a 10-fold excess of the immunizing peptides (sc-1173 p for AT1 antibody and sc-7420 p for AT2 antibody, the concentrations of specific peptides were 5 μg/ml, respectively; Santa Cruz Biotechnology) in PBS, and thereafter sections were incubated overnight at 4C. Sections treated with normal rabbit serum or normal mouse serum in place of the primary specific antibodies were used as a negative control for immunostaining of renin and ACE.
Oligonucleotide sequences of PCR primers and expected length of products
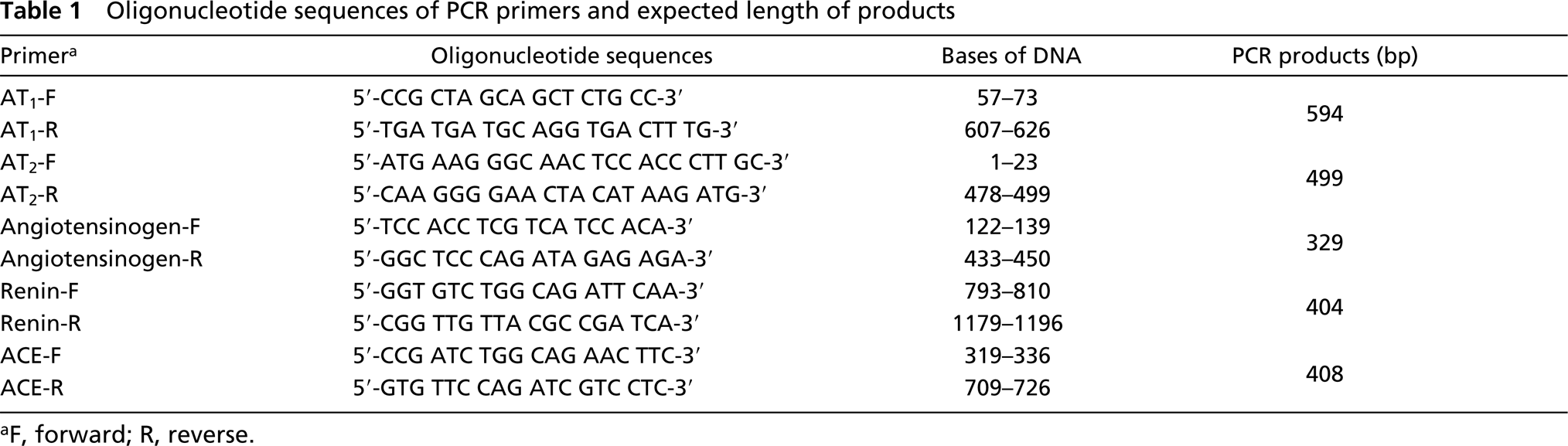
F, forward; R, reverse.
Double Immunofluorescence Studies
To confirm types of mesenchymal cells that showed immunoreactivity for AT1 receptors, we used a double-immuno-fluorescence method. Because, in kidney and heart, myofi-broblasts and macrophages are known to express AT1 receptors on their cell surfaces (Leonard et al. 1997; Sassan et al. 1998), we performed double staining of the following pairs: anti-human AT1 receptor antibody plus anti-human smooth muscle actin antibody (clone 1A4; Dako, Glostrup, Denmark) (1:200) and anti-human AT1 receptor antibody plus anti-human macrophage antibody (clone PG-M1; Dako) (1:100). Sections pretreated with preimmune goat sera were incubated in a moist chamber at 4C overnight with two of the antibodies in combination. After washing with PBS, fluorescence labeling of each of the pairs of primary antibodies was accomplished through a second incubation at RT for 1 hr in the dark with two IgG-specific fluorophore fluorescein-conjugated affinity-purified goat antibodies against rabbit IgG (heavy plus light chains; Chemicon) (1:200), and rhodamine-conjugated affinity purified goat antibodies against mouse IgG (Chemicon) (1:25). After another washing with PBS, the sections were mounted in Vectashield Mounting Medium (Vector Labs; Burlingame, CA) for analysis by fluorescence microscopy. A Zeiss Axiophot epifluorescence microscope equipped with an HBO 100-W/z mercury lamp was used. For visualization of fluo-rescein labeling, we employed two different filters to detect rhodamine and fluorescein fluorescence.
Results
RT-PCR for AT1 and AT2
AT1 receptor mRNA was found in the samples of sigmoid colon and ascending colon (Figure 1A). AT2 receptor mRNA transcripts were also detected in sigmoid and ascending colon (Figure 1B).
RT-PCR for the Components of RAS
All RAS component (angiotensinogen, renin, and ACE) mRNA transcripts were found in both sigmoid colon and ascending colon (Figure 1C). Without RT (for negative control), no mRNA was found. The nucleotide sequences of the above PCR products were identical to the known human genes.
Immunohistochemistry for Ang II Receptor
On IHC examination of human sigmoid colon, immunoreactivity for AT1 receptors appeared to be localized in vessel walls, surface epithelium, crypt bases, and in mesenchymal cells in the lamina propria (Figures 2B and 2C). To confirm the types of mesenchymal cells, we performed double immunofluorescence staining. As with IHC staining, AT1 receptor-positive cells were seen in surface epithelium, crypt bases, and in mesenchymal cells in the lamina propria (Figures 3D and 3G). Most of the mesenchymal cells below the surface epithelium were macrophages (Figure 3E), some of which showed immunoreactivity for AT1 receptors (Figure 3F). α-Smooth muscle actin was seen in crypt bases and weakly in some mesenchymal cells (Figure 3H). Double immunofluorescence staining with AT1 receptors and α-smooth muscle actin antibodies showed that some of myofibroblasts also expressed AT1 receptors (Figure 3I). Immunoreactivity for AT2 receptors was observed in parts of mesenchymal cells and weakly in parts of surface epithelium in sigmoid colon (Figures 2E and 2F). A small number of crypt cells were also immunoreactive for AT2 receptors. The ascending colon exhibited the same pattern of immunoreactivity for AT1 and AT2 receptors (data not shown). No immunostaining was found in control sections incubated with preabsorbed sera instead of primary antibodies (Figures 2A and 2D).
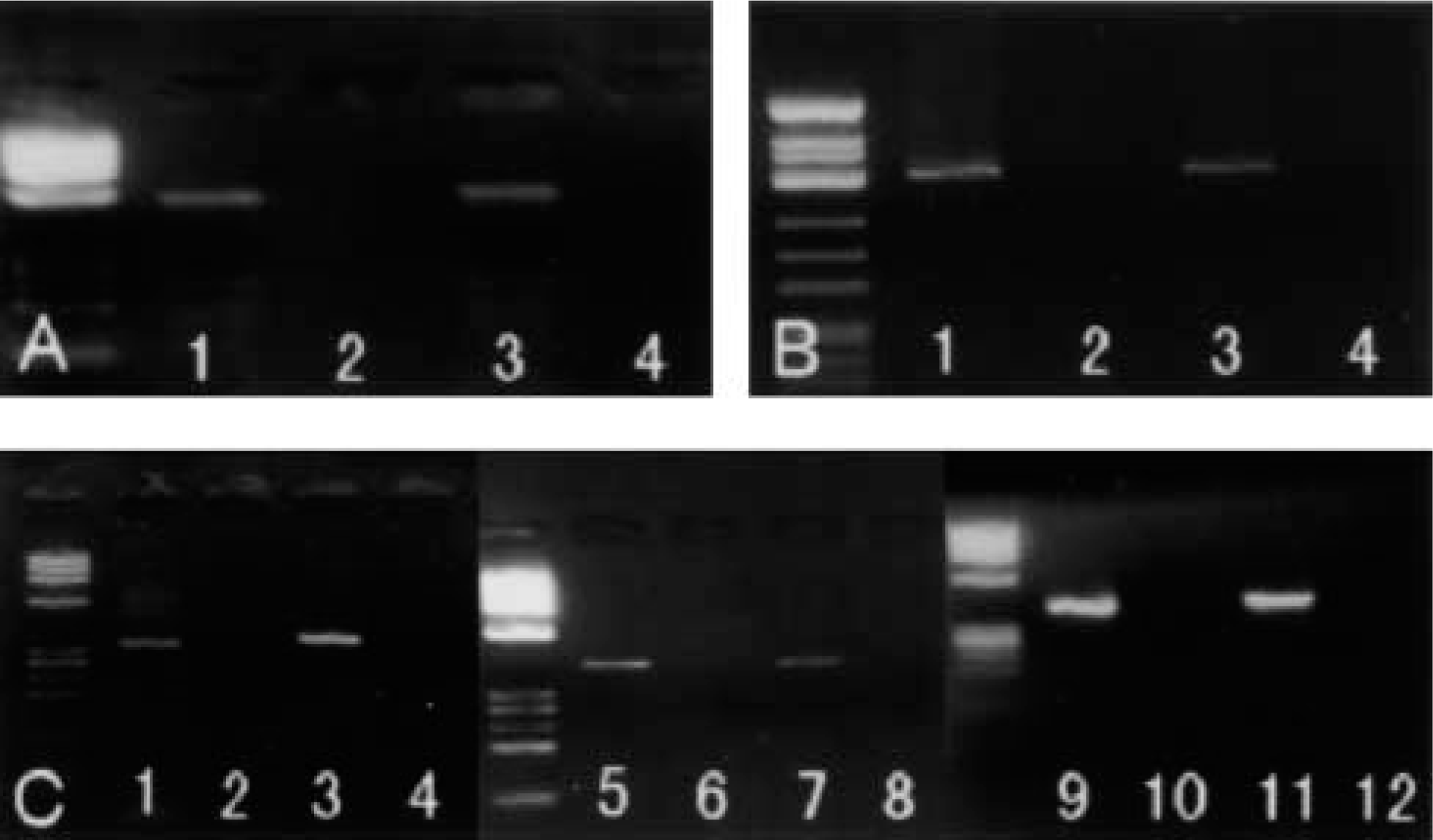
Detection of Ang II receptors and RAS components mRNA. (A) mRNA of AT1 receptor present in samples of sigmoid colon and ascending colon (Lane 1, sigmoid colon with RT; Lane 2, sigmoid colon without RT; Lane 3, ascending colon with RT; Lane 4, ascending colon without RT). (B) AT2 receptor mRNA transcripts were also detected in samples of human colon (Lanes 1 and 2, sigmoid colon; Lanes 3 and 4, ascending colon, with or without RT). (C) All RAS component mRNA transcripts were observed in both sigmoid colon (Lanes 1, 5, and 9) and ascending colon (Lanes 3, 7, and 11) [angiotensinogen (Lanes 1–4), renin (Lanes 5–8), and ACE (Lanes 9–12)]. However, without RT (for negative control) no mRNA could be found (Lanes 2, 4, 6, 8, 10, and 12).
Immunohistochemistry for Components of RAS
On IHC examination of human colon, immunoreactivity for renin appeared to be localized in the surface epithelium, vessel walls, muscularis mucosae, and in mesenchymal cells in the lamina propria (Figures 4B and 4C). Immunoreactivity for ACE appeared to be shown in vessel walls and mesenchymal cells in lamina propria and submucosa and weakly in parts of the surface epithelium in ascending colon (Figures 4D and 4E). Heterogeneity in immunoreactivity between sigmoid colon and ascending colon could not be found in renin and ACE (data not shown). No immunostaining was found in control sections incubated with preimmune sera instead of primary antibodies (Figure 4A).
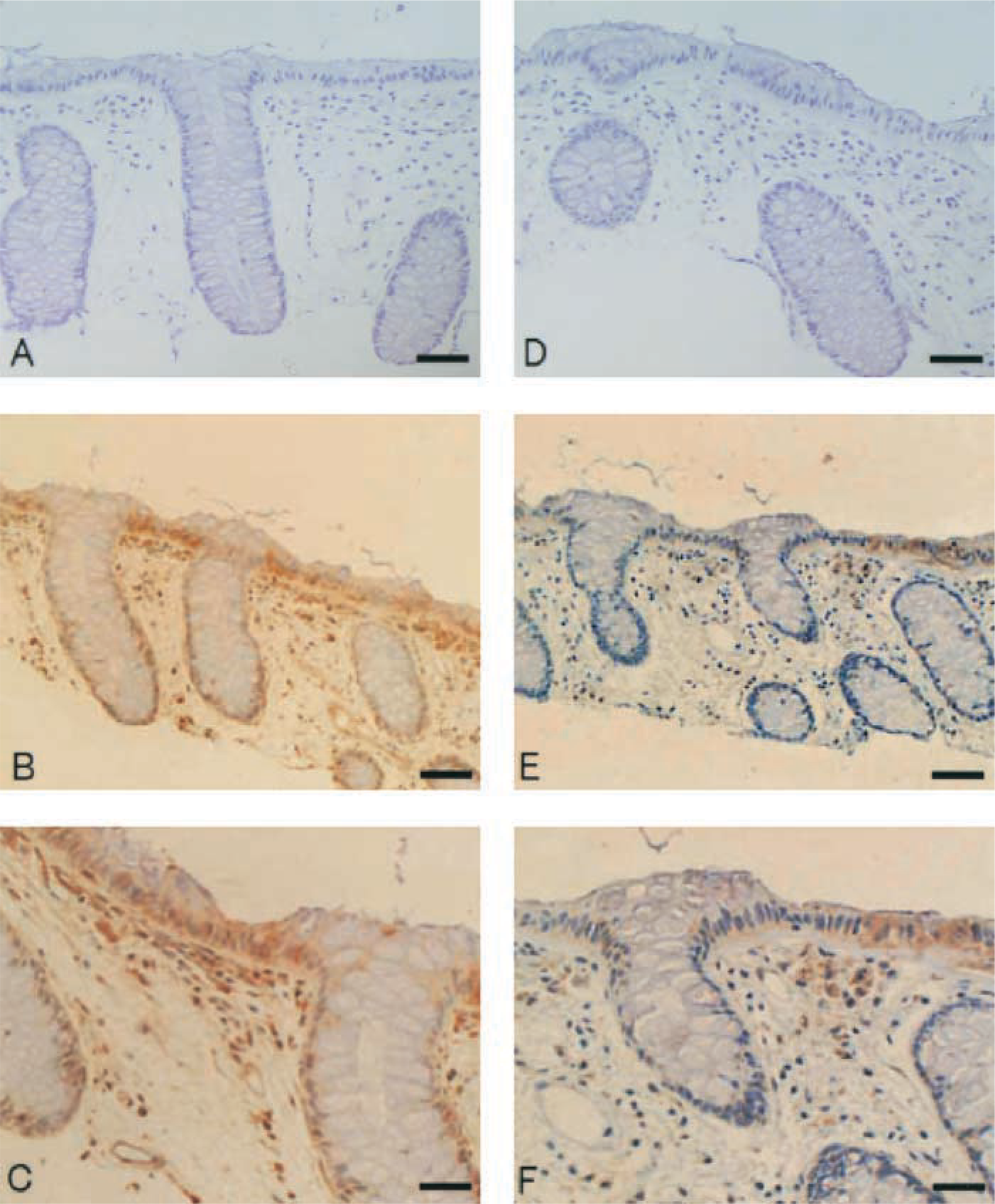
Immunohistochemical examination of human sigmoid colon revealed that immunoreactivity for AT1 receptors appeared to be localized in blood vessels, crypt base, surface epithelium, and in mesenchymal cells in lamina propria (B,C). Immunoreactivity for AT2 receptors appeared to be localized in parts of mesenchymal cells in lamina propria and weakly in parts of surface epithelium in human sigmoid colon. A small number of crypt cells also have weak immunoreactivity for AT2 (
Discussion
To the best of our knowledge, no report is available concerning localization of AT1 and AT2 receptors in human colon. Leonard et al. (1993) demonstrated that the predominant Ang II receptor in all intestinal segments is AT1, but a small population of AT2 receptors also appears to be present in rat intestine by autoradiography. In their study of the colon, Ang II binding was significantly more abundant in the muscularis than in the mucosa. In guinea pig, immunoreactivity for AT1 was seen in neural somata of the submucosa and myenteric nerve plexuses (Hosoda et al. 2000). Although surface epithelial cells also showed immunoreactivity, it was less intense than that of the nerve cells. Furthermore, crypt cells lacked significant immunoreactivity for AT1. Another study reported immunolocalization of Ang II receptors in icefish (Marsigiliante et al. 1997), showing immunoreactivity for Ang II in absorptive cells and smooth muscle cells in icefish intestine. The present study demonstrated that AT1 receptors were localized in vessel walls, myofibroblasts, and macrophages in lamina propria, crypt bases, and surface epithelium in human colon. Moreover, we showed that AT2 receptors were also present in human colon, localized in parts of mesenchymal cells, crypt cells, and weakly in surface epithelium.
Ang II has many physiological functions in vivo. Most of the functions of Ang II, including, among others, vascular constriction, cell proliferation, and release of aldosterone, are known to mediate via AT1 receptors. AT1 receptors in vessel walls in human colon mucosa may regulate vascular constriction (Dudley et al. 1990; Wong et al. 1990). Concerning electrolyte transport, Ang II stimulates water and sodium absorption across the human colon epithelium (Binder and Rawlins 1973; Shultz 1981). Low concentrations of Ang II stimulate sodium and water absorption, while high concentrations of Ang II inhibit absorption and/or stimulate secretion (Levens et al. 1981). Recently, Xio-Hong et al. (1999) demonstrated that low concentrations of Ang II stimulate water and sodium absorption via AT2 receptors and high concentrations of Ang II inhibit sodium absorption and/or stimulate sodium secretion via AT1 receptors at the serosal side of the rat jejunum. We found abundant AT1 receptors in surface epithelium, whereas AT2 receptors were weakly localized in parts of epithelial cells. Although the physiological function of Ang II receptors is still not defined, AT1 and AT2 receptors in surface epithelium may regulate water and sodium transport in each other.
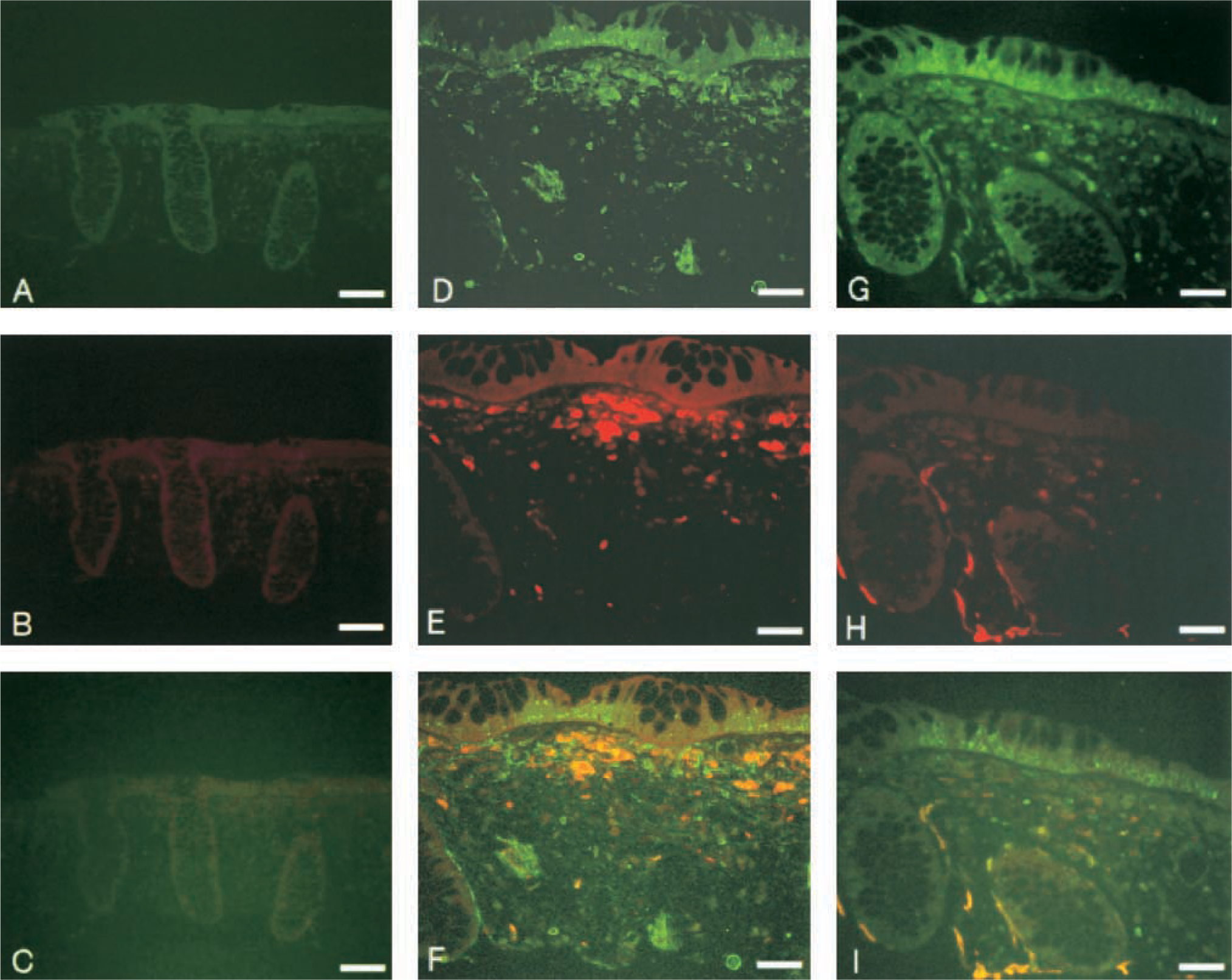
Immunofluorescence staining showed AT1 receptor-positive cells (green) in surface epithelium, crypt bases, and in mesenchymal cells in lamina propria (
Intestinal subepithelial myofibroblasts (ISEMFs) are located in the lamina propria under the epithelial cells. They exist as a syncytium and have important roles in releasing chemical mediators and extracellular matrix proteins (Powell et al. 1999). With the release of chemokines, myofibroblasts regulate epithelial growth and differentiation, tissue repair and fibrosis, immunological response, and intestinal absorption and secretion. Some studies have revealed AT1 and AT2 receptors exist in both fibroblasts and leukocytes, affecting cell proliferation and differentiation (Sadoshima and Izumo 1993; Tsutsumi et al. 1998). In T84 colonocytes, stimulation of Cl− secretion is confirmed as a result of cAMP increase induced by prostaglandins (principally PGE2) that are released from myofibroblasts (Barrett 1993). Naftalin and colleagues (1999) showed that the ISEMF sheath represents a diffusion barrier to Na+ transport that creates a significantly hypertonic middle compartment between epithelial tight junctions and the vascular system (Naftalin and Pedley 1999; Naftalin et al. 1999). Furthermore, they demonstrated proliferation of the ISEMF sheath in response to dietary NA+ depletion, which would account for the increase in Na+ and water absorption in high Ang II-high aldosterone states. In our study, the mesenchymal cells that showed immunoreactivity for AT1 receptors were identified as myofibroblasts and macrophages. Although we cannot clarify the function of receptors expressed in ISEMFs, it is possible that the Ang II receptors have similar roles locally, such as cell proliferation, water and electrolyte absorption, and secretion.
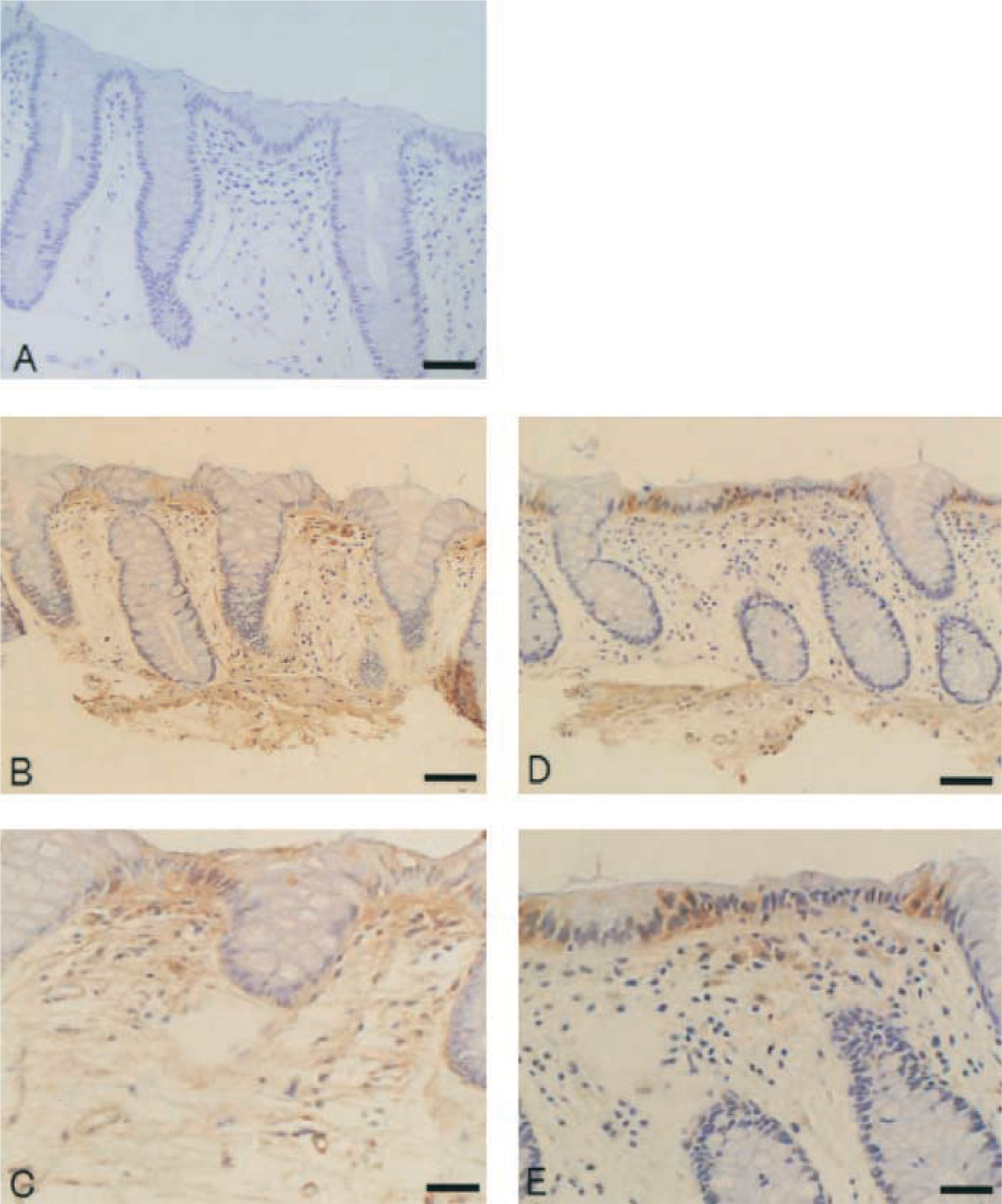
Immunohistochemical examination of the human sigmoid colon revealed that immunoreactivity for renin appeared to be localized in surface epithelium, vessel walls, muscularis mucosae, and in mesenchymal cells in lamina propria (B,C). Immunohistochemical examination of the human ascending colon revealed that immunoreactivity for ACE appeared to be localized in vessel walls and in mesenchymal cells in lamina propria (
Classically, angiotensin has been considered as a plasma hormone. The discovery of components of the RAS in many tissues has led to the hypothesis that, in addition to being a plasma hormone, Ang II may be formed locally and may therefore possess an additional paracrine and/or autocrine action. There are currently inconsistent issues concerning the origin of tissue RAS components in the cardiovascular system. It has not been clearly shown whether or not local RAS components are derived from plasma or local production in tissues, or from both (Victor and Richard 1994). A perfused rat heart model showed there are two compartments for local production of angiotensin: an intravascular compartment and an interstitial fluid compartment (Larissa et al. 1997). At the vascular surface, angiotensinogen is converted to Ang I by endothelium-bound renin (intravascular compartment). On the other hand, some circulating renin and angiotensinogen is delivered to the interstitial fluid, thereby producing Ang I in the interstitium (interstitial fluid compartment). Recently, the concept of local RAS appears to indicate that local generation of Ang II by RAS components is more important than the origin of RAS components. For the gastrointestinal tract, the findings related to the existence of a local RAS are insufficient. We detected renin and ACE in surface epithelium, vascular walls, and mesenchymal cells, and expression of angiotensinogen mRNA in human colon mucosa. Although we cannot determine their origin, the present study suggests the possible existence of local RAS in human colon.
In conclusion, we obtained the following findings. (a) AT1 receptors were localized in vessel walls, macrophages, and myofibroblasts in the lamina propria and in the surface epithelium of human colon. (b) AT2 receptors were localized in parts of mesenchymal cells and weakly in the surface epithelium. (c) Renin and ACE were distributed in parts of the surface epithelium, vessel walls, and mesenchymal cells. These findings suggest the possibility that RAS exists in human colon mucosa. However, the definition of existence of local RAS is required to demonstrate generation of Ang II in colon, in addition to the existence of its components. Further studies will be needed.
Footnotes
Acknowledgements
We thank Dr Akiyoshi Fukamizu for his kind gift of antihuman renin antibody. The help of Ms Satoko Takebayashi, Ms Yoko Takei, Ms Hiroko Murakami, Ms Kazuko Tsukioka, and Ms Kyoko Nishitani, the nurses in the Division of Endoscopy, is greatly appreciated.