
Abstract
Bone cells respond to mechanical stimulation by gene expression. The molecular events involved in the translation of mechanical stimulation into cell proliferation and bone formation are not yet well understood. We looked for the expression of early-response genes of the AP-1 transcription factor complex in an in vivo bone regeneration system subjected to mechanical forces because these genes were found to be related to mechanotransduction and important for bone development. Sheep maxillary bone was distracted daily for 15 days. c-Jun and c-Fos were evaluated by Northern blotting analysis and immunohistochemistry in biopsy specimens removed at 8 and 15 days and were compared with post-osteotomy but not distracted repair tissue. Elevated levels of c-Jun and c-Fos mRNA were found after 8 days of distraction. Likewise, mesenchyme-like and fibroblast-like cells composing the 8-day distracted regeneration tissue showed increases in the intensity of immunostaining compared to cells in the corresponding non-distracted fracture repair tissue. After 15 days of distraction, when bone trabeculae start to form distally and proximally in the distracted regeneration tissue, mostly preosteoblasts and osteoblasts retained c-Fos and c-Jun immunoreactivity, similar to bone-associated cells in control non-distracted fracture repair tissue. We propose that the elevated expression of c-Jun and c-Fos is related to mechanical stimulation in this in vivo bone regeneration system.
O
The nuclear protooncogenes c-fos, c-jun, fra-2, and jun-D, the expression of which has been followed in the present study, are members of a family of transcription factors that contain a leucine-rich region and a basic region, which are necessary for dimer formation and DNA binding, respectively. Dimerization is a prerequisite for DNA binding to a consensus response element site (5′-TGAG/CTCA-3′) designated activator protein-1 (AP-1) (Kouzardief and Ziff 1988). AP-1 sites are present in the promotors of many developmentally regulated osteoblast genes, including alkaline phosphatase, collagen type I, collagenase, and osteocalcin. For example, fra-2 and jun-D have been shown to be important for enhanced expression of osteocalcin by mature osteoblasts (Katai et al. 1992; Lian et al. 1998).
In the present study we took advantage of an in vivo membranous bone regeneration system that is under continuous mechanical stimulation, to gain more insight into the translation of mechanical force into gene expression in developing bone (Rachmiel et al. 1998,2002; Nomura and Takano-Yamamoto 2000; Lewinson et al. 2001). We followed the expression of several AP-1 members in distracted sheep maxillary bone and compared them with non-stimulated similarly fractured bone on both the mRNA level and the protein level. We demonstrate an enhanced expression of both c-fos and c-jun in the stretched pre-osteogenic fracture callus relative to non-distracted repair tissue and suggest that the mechanical stimulus applied was translated into stimulation of the expression of early-response genes of the AP-1 family of transcription factors.
Materials and Methods
Seven young adult sheep aged 1 year were operated on in this study. The sheep head was chosen because the sheep is a large mammal and can provide successive biopsies. Surgery was performed under general anesthesia. All the sheep underwent complete maxillary osteotomy from the right side to the left side as described earlier by us (Lewinson et al. 2001; Rachmiel et al. 2002). Great care was taken to preserve the periosteum intact while cutting the bone by subperiosteal tunneling. A distraction device was mounted and fixed by pins to the maxilla and to the infraorbital region. The animals were treated prophylactically with antibiotics (cephamezin 1 g three times daily, IV) for the next 5 days. After a latency period of 5 days, separation of the osteotomized edges of the bone by distraction was started in five sheep by lengthening the device at a rate of 1 mm/day for up to 15 consecutive days. In two sheep the distraction device was left untouched and no separation of the osteotomized edges was applied. These sheep served as non-distracted repair tissue controls. All the sheep provided tissues for the IHC studies. From four sheep (two experimental and two control), removed tissues served for Northern blotting analysis.
Biopsies were removed under general anesthesia 30 min after the ninth and sixteenth distraction manipulations following 8 or 15 days of distraction, respectively, and in parallel from the non-distracted sheep on the corresponding days. The biopsies bridged the whole distracted or the non-distracted regeneration tissue from the first to the second osteotomy line. These two temporal points were chosen because bone trabeculae start to form proximally and distally from the cut edges of the osteotomized bone usually later than 8 days of distraction. No osteogenic process is yet under way after 8 days of distraction and the same is true for the corresponding controls. By 15 days of distraction the osteogenic process is well under way and trabeculae can be observed to grow from both bone edges in the direction of the center of the regeneration tissue. Each temporally different biopsy was removed from one side of the face and the next one from the other side as described in detail earlier by us (Lewinson et al. 2001; Rachmiel et al. 2002). For Northern blotting analyses the biopsies were immediately immersed in RNA (Ambion; Austin, TX) and kept at 4C until the day of isolation. For protein localization by IHC, part of the biopsies were either frozen directly in liquid nitrogen-precooled isopentane and kept at −80C or were immersed in 4% paraformaldehyde for 48 hr, dehydrated, and embedded in paraffin. Eight-μm-thick frozen sections were cut in a Leica cryostat (Jung Frigocut 2800E) and 6-μm-thick paraffin sections were cut with the aid of a Leica microtome (RM 2135). Animals were treated after each biopsy with cephamezin (1 g IV three times daily for 48 hr). After each daily distraction, polydine was applied locally to prevent infection. The Institutional Committee for Animal Care approved all the surgical procedures and animal handling. No feeding problems were encountered.
RNA Extraction and Northern Blotting Analysis
Total RNA was isolated using Tri-Reagent (Molecular Research Center; Cincinnati, OH) according to the manufacturer's protocol. Each sample of 25 μg of total RNA was separated by electrophoresis in 2.2 mol/liter formaldehyde with 1.0% agarose gel containing 10 mg/ml ethidium bromide. After electrophoresis, the gel was examined with UV light to visualize ribosomal RNA bands to ensure quality and equal loading of the samples. RNA was transferred to a nylon membrane (Amersham; Arlington Heights, IL). Membrane was incubated for 3 hr with hybridization buffer [1 M NaH2PO4, 1 M Na2PO4, 7% sodium dodecyl sulfate (SDS), 0.5 M EDTA, 1% bovine serum albumin (BSA), pH 8.0] and then incubated at 65C in the same buffer containing the [α-32p]-d-ATP labeled specific complementary DNA (cDNA) for human c-jun and c-fos (Oncogene Research Products; Cambridge, MA) that was labeled with [α-32p]-ATP. Membrane was washed for the first time with 2 × saline sodium citrate (SSC) (1 × = 1.5 mM sodium citrate, 15 mM NaCl, pH 7) and 0.1% SDS at RT, followed by another wash with 2 × SSC and 0.1% SDS at 65C for 30 min and a third wash with 2 × SSC and 0.1% SDS for 30 min at RT. After a last wash with 2 × SSC for 5 min at RT, the membrane was exposed to autoradiographic film (Kodak; Rochester, NY) for 5 days at −70C.
Immunohistochemistry
The antibodies used to identify cells that express the early-response genes of the AP-1 family were anti-c-fos, anti-c-jun, anti-fra-2, and anti-jun-D (Santa Cruz Biotechnology; Santa Cruz, CA). All were affinity-purified rabbit polyclonal antibodies. According to the manufacturer, anti-c-fos does not crossreact with fos-B, fra-1, or fra-2. Anti c-jun does not crossreact with jun-B or jun-D. Anti-fra-2 does not crossreact with c-fos, fra-1, or fos-B.
The immunohistochemical staining was performed either on paraffin sections or on frozen sections. Ablation of endogenous peroxidase activity by 3% H2O2 in absolute methanol (not performed on frozen sections) was followed by blocking of unspecific binding by 1% BSA in PBS containing 0.01% Tween-20. The sections were then immunostained by the primary antibodies for 1-1.5 hr at RT. This was followed by the biotinylated goat anti-rabbit second antibody, employing the streptavidin-biotin-peroxidase method and AEC as substrate according to the manufacturer's directions (Histostain plus kit; Zymed Laboratories, San Francisco, CA). Replacing the primary antibody by pre-immune rabbit serum served as negative control.
Results
Northern Blotting Analysis
RNA was extracted from biopsies that contained the whole callus that was last distracted 30 min earlier. The biopsies were removed from experimental sheep that were distracted for 8 and 15 days after a prior latency period of 5 days and from the corresponding fractured, but not distracted sheep maxillae that served as controls. Because the yield of mRNA was very poor, we had to pool the extract from the callus of the two experimental animals and from the two control sheep. The Northern blotting analysis for c-fos mRNA showed that the mRNA levels were upregulated in regeneration tissue removed from animals that were distracted for 8 days (receiving the last stimulus of distraction 30 min before removal of the tissue) compared with the mRNA levels in tissues from the corresponding control animals. At this stage of regeneration the distracted tissues consist of mesenchyme and fibroblast-like cells. No overt bone elements are yet present. A representative gel separation of c-fos is shown in Figure 1A. c-fos mRNA level has increased about ninefold over control as calculated from densitometry measurements (Figure 1C). mRNA levels of c-jun were likewise elevated after daily distractions for 8 days in the experimental samples relative to the control samples about fivefold, (Figures 1B and 1C). Northern blotting analysis for c-jun mRNA from 15-day distracted callus did not show any difference between control and experimental samples (not shown). c-fos mRNA levels in 15-day tissues was not analyzed because of the low yields of tissue from the control non-distracted sheep. Control for loading is shown by ethidium bromide staining of 18S and 28S RNAs. We could not use GAPDH for normalization because there is no specific probe for sheep tissues.
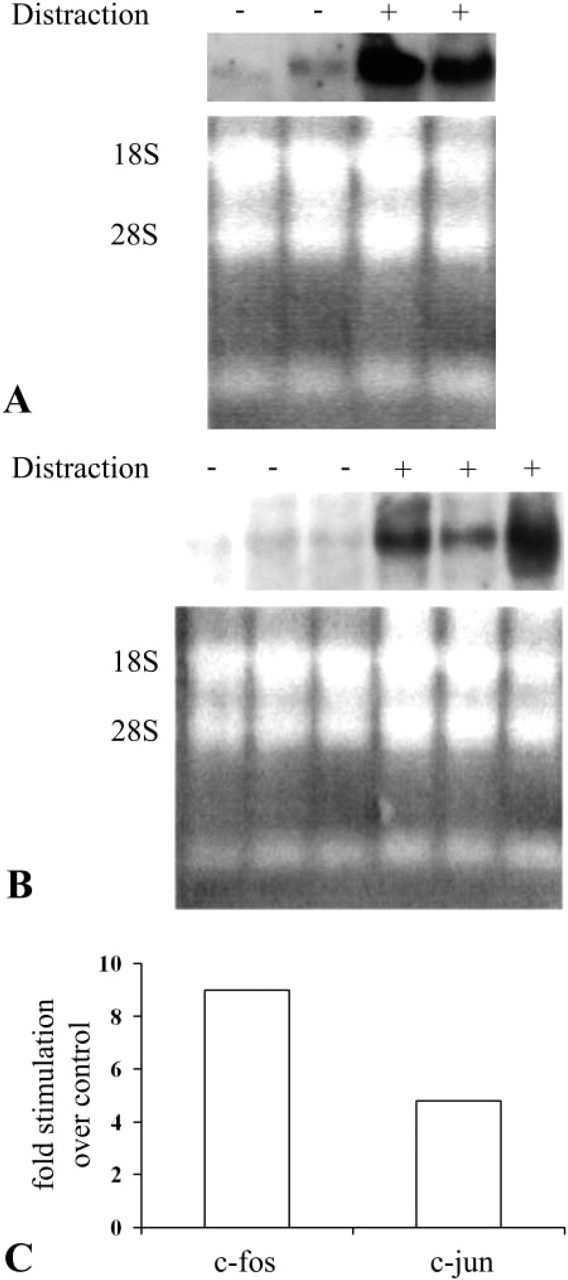
AP-1 Protein Immunohistochemistry
Compatible with the mRNA findings, c-Fos and c-Jun proteins, as exposed by the IHC staining, were revealed in paraffin sections of 8-day distracted tissues in more than 50% of the cells. In comparison, the control non-distracted tissues did not reveal positively stained cells (Figures 2A–2E, Table 1). The staining was mainly confined to the cytoplasmic compartment of the polymorphic mesenchyme-like cells, which are the main components of the distracted callus at this stage. These cells might be regarded as preosteogenic cells because they already express Cbfa-1 (Lewinson et al. unpublished data). Fra-2 protein immunostaining was quite impressive in both 8-day distracted tissues and in the respective non-distracted control regeneration tissue (Figures 2G and 2H). In the 8-day distracted samples, Jun-D protein was expressed in lining cells of the capillaries and in some mesenchyme-like cells (Figure 2F). Control tissues were not checked for Jun-D. Tissues in which the primary antigen was substituted with non-immune rabbit serum were negative (Figure 2I).
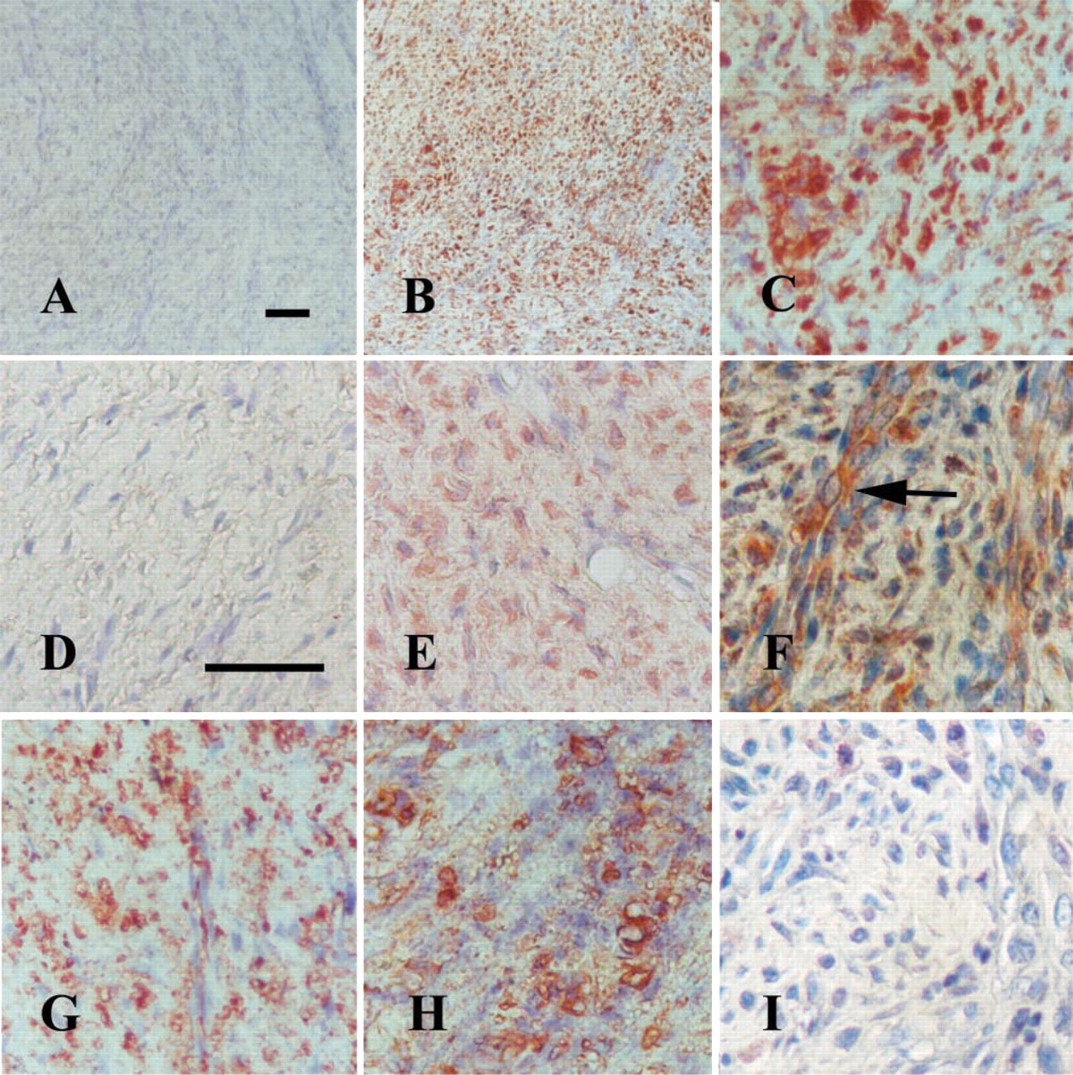
Eight-day distraction: c-Fos (
After 15 days of distraction, the distraction gap consists mainly of a fibrous zone, the cells of which are fibroblast-like secreting a collagen-rich matrix (Rachmiel et al. 2002). These cells also express, in addition to Cbfa-1 mRNA and protein, the message for osteocalcin (Lewinson et al. unpublished data). Therefore, they might be regarded as the progenitors of the preosteoblasts that contribute to the osteogenic front at the trabecular tips. The trabeculae start to grow from both edges of the fractured maxilla directing to the center of the distraction gap and are oriented along the distraction force.
Prominent immunostaining for c-Fos, Jun-D, and Fra-2 proteins was demonstrated in the longer (15-day) distracted calluses in many of the fibroblast-like cells (Figures 3A, 3C, and 3D), and less intense expression of c-Jun and in fewer cells (Figure 3B; Table 1), but most intense in the new bone trabeculae that begin to form at the edges of the biopsies. All proteins were intensely revealed in preosteoblasts, osteoblasts, and osteocytes (Figures 3E–3L). In parallel, in the control fractured but non-distracted repair tissues (of a total regeneration time of 20 days), similar strong immunostaining was revealed in all the bone-associated cells and the mesenchyme-like cells that are still recruited to the newly formed trabeculae but, interestingly, no staining could be observed in the fibrous zone that separates the edges of the bone trabeculae (Figures 4A and 4B). When rabbit non-immune serum was substituted for the primary antibody, no positive staining could be observed (Figure 4C). A semiquantitative evaluation of the results of the immunostaining of all the proteins tested in this study in distracted tissues is summarized in Table 1.
Relative immunostaining of the AP-1 family proteins in distracted tissuesa
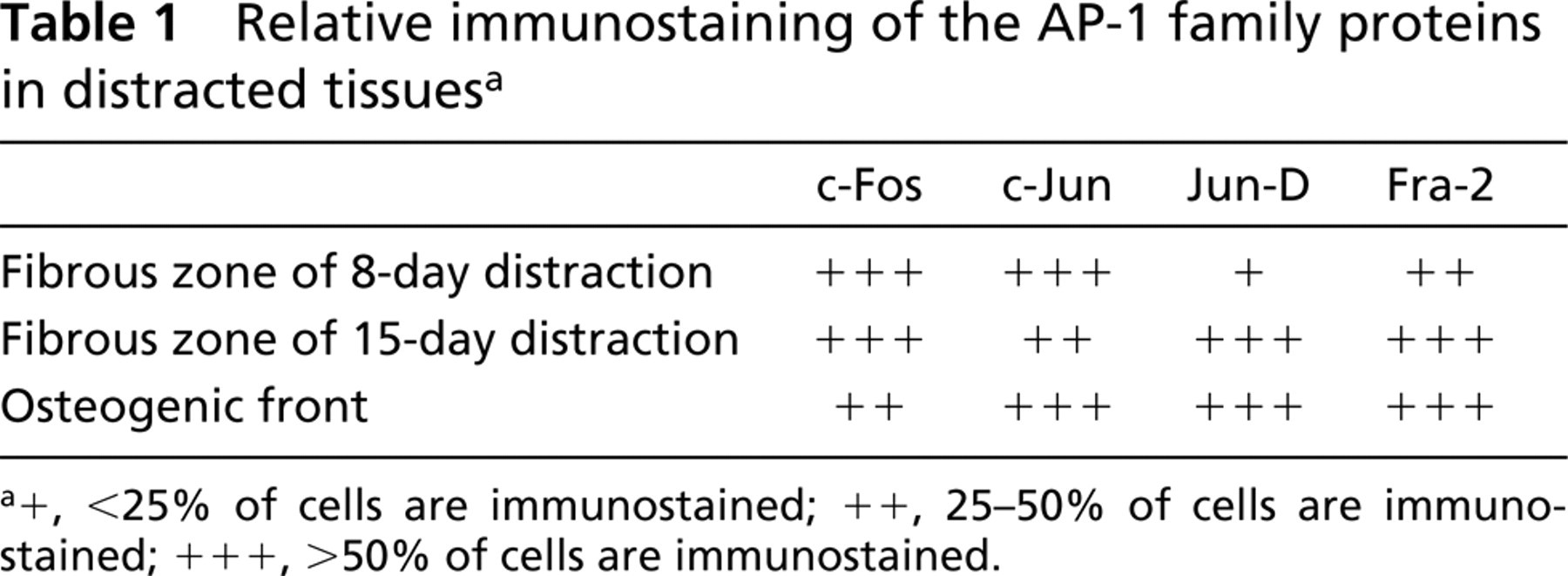
+, <25% of cells are immunostained; ++, 25-50% of cells are immunostained; +++, >l50% of cells are immunostained.
Discussion
Bone formation by osteoblasts is essential not only for skeletal growth and bone remodeling but also for bone healing and repair. Several hormones and growth factors that are implicated in the regulation of bone physiology are now known to upregulate the expression of proteins of the AP-1 complex (Liang et al. 1999; Palcy et al. 2000; Stanislaus et al. 2000; Varghese et al. 2000). Several genes coding for bone-associated proteins, such as collagen type I, alkaline phosphatase, osteocalcin, collagenase-3, and parathyroid hormone (PTH)/PTH-related peptide receptor contain an AP-1 response element in their promoter (Katai et al. 1992; Lian et al. 1998; Varghese et al. 2000). Moreover, one of the AP-1 proteins, c-Fos, has been implicated in transduction of mechanical stimulation to bone cells (Lean et al. 1996; Peverali et al. 2001).
In the sheep model of the present study, a continuous mechanical strain/stress is exerted on the regenerating callus beginning 5 days after an osteotomy and external fixation of the maxilla (which is a component of the craniofacial complex). It is assumed that the distraction force, which distracts both edges of the fractured bone one from the other for 1 mm daily, maintains the strain force as a result of the external fixation until the next distraction on the following day. Several studies as well as our own have confirmed the very early observations of Ilizarov that the distraction force enhances and maintains proliferation of the cells that constitute the callus replacing the initial hematome (Ilizarov 1988; Aronson et al. 1995; Li et al. 1997; Rachmiel et al. 2002). Except for a few in vivo studies in which vertebrae were loaded by compression (Inaoka et al. 1994; Lean et al. 1996) or by employing a hydraulic chamber (Moally et al. 2000), all the other studies observing that mechanical force induces the expression of early-response genes of the AP-1 transcription factor family, and c-fos in particular, employed in vitro systems (McCabe et al. 1995; Fitzgerald and Hughes–Fulford 1999; Wozniak and Hruska 2001; Granet et al. 2002; Kletsas et al. 2002). The strength of our study is in offering an insight into an in vivo system in a large mammal by imitating a newly applied clinical practice for correcting facial aberrations and reconstructions after trauma or metaplasia. Clarifying the molecular events by using such an animal model will help to develop procedures that might enhance and ameliorate the osteogenic process.
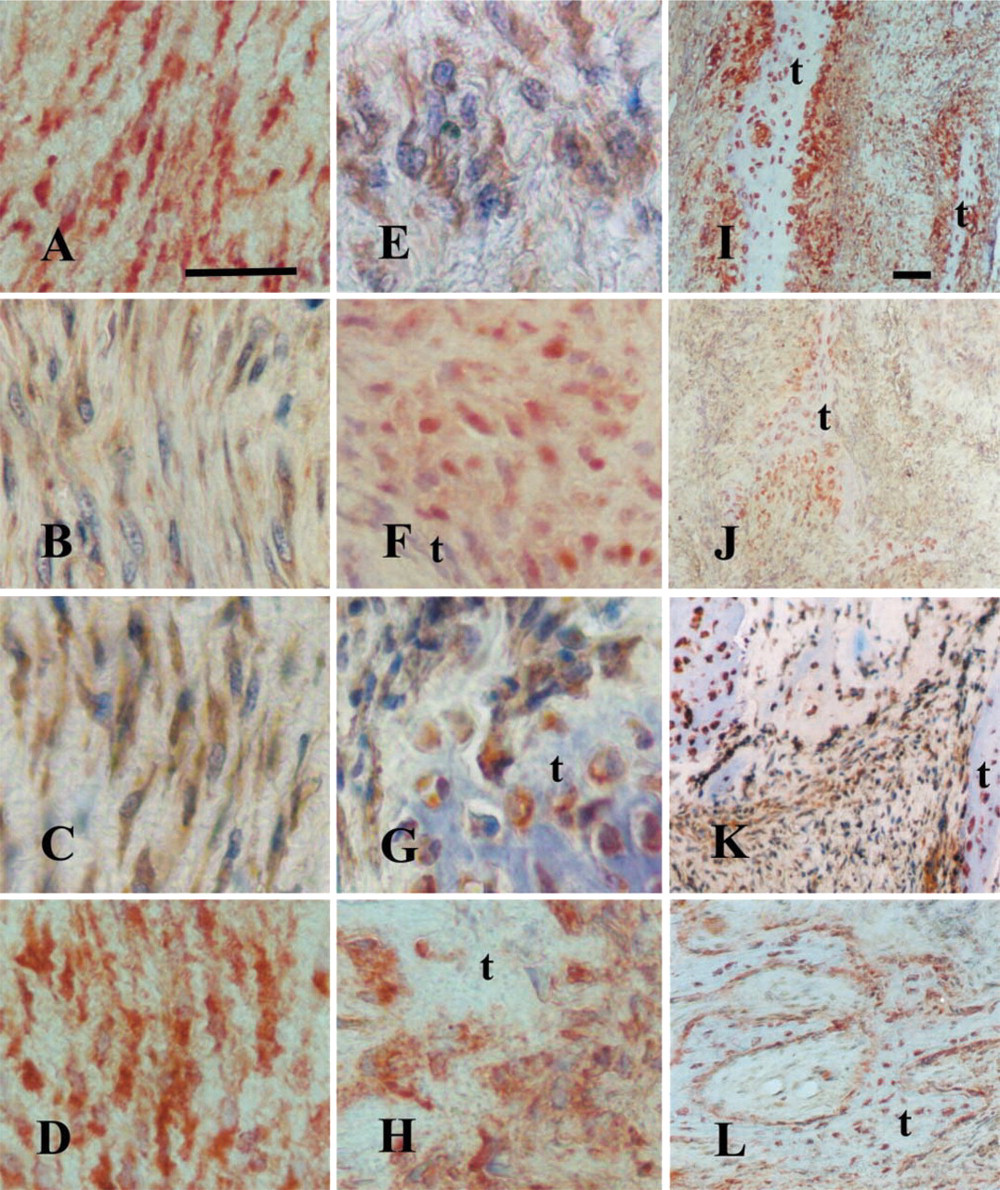
Fifteen-day distraction: c-Fos (
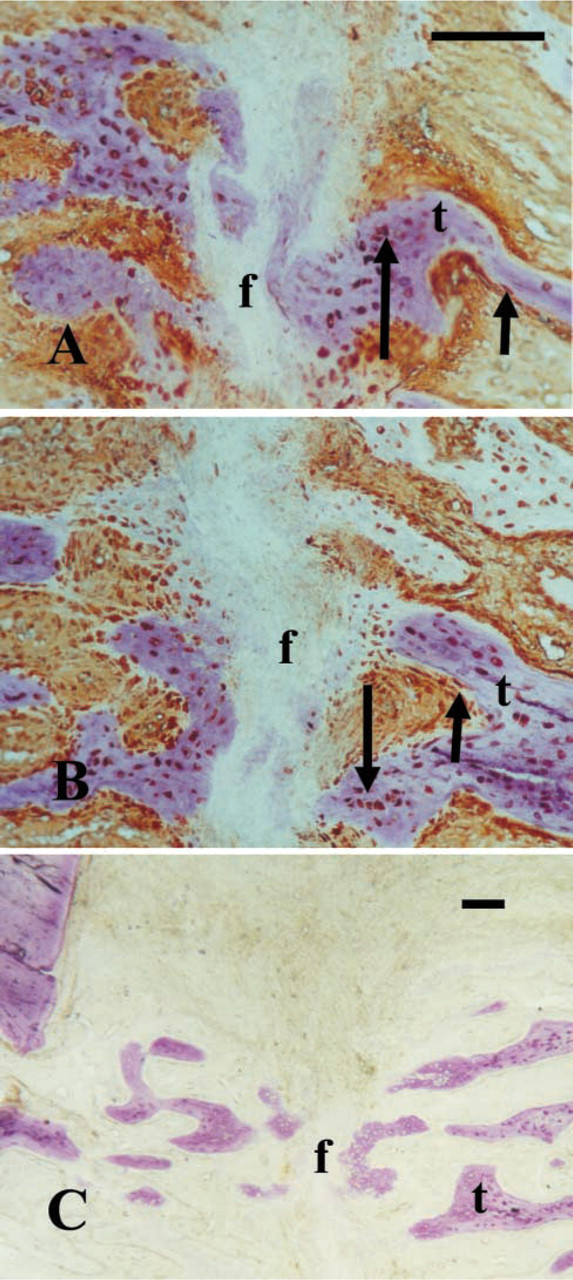
Control repair tissue of osteotomized but not distracted fracture of 20 days: (
We show an increase in the expression of c-fos and c-jun both on the mRNA level and, by IHC, on the protein level. The latter technique offers the advantage of identifying the specific cells that synthesize the proteins and their intracellular localization. Although AP-1 family proteins are usually regarded to be nuclear, we found that most of the immunostaining was cytoplasmic (Granet et al. 2002). This corresponds to the data described by Wozniak and Hruska (2001) in mechanically strained human osteoblasts, where the AP-1 stimulated proteins were localized to the endoplasmic reticulum, compared with nuclear localization in non-strained osteoblast progenitors. This increase was evident after 8 days of distraction in distracted calluses compared with the non-distracted control tissues. At this stage there are still no signs of overt bone formation in both conditions of repair. Therefore, the increase in c-fos and c-jun expression and in protein synthesis relates to the mechanical stimulation applied. This increase is observed even though in normal fracture repair that is externally fixated, a small freedom of movement is also experienced. Distracted tissues continued to show expression throughout the period of the study, which was 15 days of distraction. When bone trabeculae start to form from the edges of the fractured bone (starting usually between the ninth and tenth days of distraction) and grow in the direction of the central zone of the callus, c-fos is expressed not only in mesenchymal cells but also in the fibroblast-like cells that comprise the fibrous zone and can be regarded as osteoblast progenitors because they already express Cbfa-1 (Lewinson, unpublished data). The latter are the largest component of the distracted callus at this stage. In the control non-distracted repair tissue, the respective fibrous component is relatively very small (because it does not undergo distraction), comprising the area between the newly formed bone trabeculae, and does not express c-fos, c-jun, or fra-2. On the other hand, bone-associated cells that lie adjacent to the osteoid or embedded in it (osteoblasts and osteocytes) show intense immunostaining with the tested AP-1 antibodies, irrespective of whether or not the regenerating tissue was stretched. Therefore, we suggest that c-fos and c-jun are constitutive genes of bone correlating with fully differentiated bone cells, but not with cells that comprise the prebone. There, these genes are turned on only when mechanical force is applied. On the other hand, fra-2 could be visualized with a similar intensity both in distracted and non-distracted tissues even in the short-term biopsies. This might indicate that fra-2 is a constitutive gene of not fully differentiated cells independent of mechanical stimulation. This stands in contradiction to other findings in the literature that associate fra-2 with differentiated bone cells (McCabe et al. 1995,1996). This discrepancy could be explained by the very different systems used in both studies, i.e., regenerating callus vs primary osteoblast cultures of fetal calvarial cells. Recently, the importance of DeltaFosB, which is a naturally occurring truncated form of FosB, for bone formation has been reported (Sabatkos et al. 2000; Sims et al. 2002). It will be interesting to check this important AP-1 family member in our in vivo system.
In conclusion, we have demonstrated that mechanical stimulation of regenerating bone by daily distraction and under normal physiological conditions stimulates the expression of early-response genes of the AP-1 family of transcription factors.
Footnotes
Acknowledgements
Supported in part by the Lazaroff Fund for Research in Maxillofacial Surgery and by a grant from the Ministry of Health, grant number 3785, Israel.