
Abstract
Cathepsin B is a lysosomal cysteine proteinase that may participate in cancer progression. We compared localization of its protein and activity during progression of human colorectal cancer. In adenomas and carcinomas, protein expression and, particularly, activity were elevated compared with those in normal colorectal mucosa. In normal mucosa, cathepsin B protein expression was moderate in stroma and variable in epithelium, whereas activity was mainly present in distinct areas of stroma directly underneath the surface of the colon and in epithelium at the surface of the colon. Stroma in adenomas and carcinomas contained moderate to high protein levels but little activity except for areas of angiogenesis, inflammation, and necrosis, in which activity was high. In adenomas and the majority of well-differentiated carcinomas and moderately differentiated carcinomas, cathepsin B protein and activity were found in granular form in the epithelium, close to the basement membrane. Protein and activity levels were low and diffusely distributed in cancer cells in the remainder of the well-differentiated and moderately differentiated carcinomas and in all poorly differentiated carcinomas. Invasive fronts in most cancers contained moderate protein levels but high activity. We conclude that (a) activity localization is essential to understand the role of cathepsin B in cancer progression, and (b) cathepsin B activity in human colon is associated with invasion of cancer cells, endothelial cells, and inflammatory cells, and in cell death, both apoptotic and necrotic.
Keywords
I
Proteinases can facilitate invasion and metastasis in a number of ways, including detachment of individual cells from the primary tumor, penetration of surrounding tissues and the vascular system by degradation of basement membranes and extracellular matrix (ECM), adhesion, and detachment by shedding adhesion molecules (Kohn and Liotta 1995; Basbaum and Werb 1996; Hooper et al. 1997). Although upregulation of proteinase expression facilitates the invasive process, control must be retained, otherwise there would be unrestricted proteolysis (Liotta et al. 1991; Kohn and Liotta 1995; Basbaum and Werb 1996; Hooper et al. 1997). Part of the control may be activation of proteinases or association to the plasma membrane of either cancer or stromal cells after secretion into the extracellular environment (Johnsen et al. 1998; Brünner et al. 1999).
Cathepsin B is the major representative of the cysteine proteinases and is present in lysosomes of all types of cells under normal conditions (Kirschke et al. 1995). It is involved in lysosomal turnover of proteins, including collagen, fibronectin, proteoglycans, elastin, and basement membrane-specific components (Everts et al. 1985; Maciewicz et al. 1990; Van Noorden and Everts 1991; Buck et al. 1992). The enzyme exhibits broad substrate specificity at both acid and neutral pH (Maciewicz et al. 1990; Buck et al. 1992; Werle et al. 1997) and functions both as an exopeptidase and as an endopeptidase (Keppler and Sloane 1996). The enzyme is synthesized as a precursor, which is enzymatically inactive (Mort and Recklies 1986; Nishimura et al. 1988; Frosch et al. 1999). Many human cancer cell lines with invasive potential have been proved to exhibit qualitative and quantitative differences in the regulation of cathepsin B expression, including increased synthesis, vesicular localization (Frosch et al. 1999), secretion of the inactive proform into the extracellular environment (Sloane et al. 1986; Maciewicz et al. 1989; Frosch et al. 1999), and association with the plasma membrane (Sloane et al. 1986; Erdel et al. 1990; Rozhin et al. 1994; Spiess et al. 1994; Strohmaier et al. 1997; Van Noorden et al. 1998a; Frosch et al. 1999). This membrane-bound form was proved to be active (Sloane et al. 1986; Rozhin et al. 1994; Spiess et al. 1994; Van Noorden et al. 1998a; Demchik et al. 1999; Frosch et al. 1999), suggesting proteolytic activity at the cancer cell surface. Cathepsin B has been implicated in the direct degradation of the ECM by normal cells (Reddy et al. 1995; Afonso et al. 1999) and by cancer cells during invasion (Maciewicz et al. 1990; Buck et al. 1992; Coulibaly et al. 1999; Webb et al. 1999), and by endothelial cells during angiogenesis (Keppler et al. 1996; Kostoulas et al. 2000). It is also conceivable that cathepsin B is engaged in proteolytic cascades to activate other proteinases which, in turn, mediate ECM degradation (Kobayashi et al. 1991; Sivaparvathi et al. 1995; Van Noorden et al. 1998a; Ramos–DeSimone et al. 1999). In vivo inhibition of cathepsin B activity delays the onset of growth of primary tumors (Van Noorden et al. 2000) and downregulates metastasis of several types of cancer in vitro and in animal models (Kobayashi et al. 1992; Redwood et al. 1992; Navab et al. 1997; Van Noorden et al. 1998a).
The role of cathepsin B in progression of colorectal cancer has been studied but the results are contradictory. This is particularly caused by the application of different techniques. For example, Campo et al. (1994) and Keppler et al. (1988) localized immunohistochemically the total amount of cathepsin B protein, including pro-enzyme, active enzyme, and enzyme complexed with inhibitor. It was found in human colorectal carcinomas that expression of cathepsin B protein correlates with tumor progression and shortened patient survival. On the other hand, cathepsin B mRNA levels and activity were determined in homogenates of human colorectal carcinomas by Murnane and co-workers (Sheahan et al. 1989; Murnane et al. 1991; Shuja et al. 1991; Iacobuzio-Donahue et al. 1997; Del Re et al. 2000). These authors found the highest levels of mRNA and activity in early stages of tumor progression. However, information on activity in relationship to specific sites in the tumors, such as regions of invasion, were not obtained in these studies. Cathepsin B activity has been determined in microdissected areas of invasive human colon cancer. These areas showed a two-to three-fold increase over the activity in normal epithelium (Emmert–Buck et al. 1994). However, only a few cases were studied and a correlation with tumor progression could not be made. Therefore, the objective of the present study was the establishment of the exact localization of both protein and activity of cathepsin B in serial sections of human colorectal carcinoma biopsies with the use of immunohistochemistry, enzyme histochemistry, and image analysis to link cathepsin B protein and activity distribution patterns with tumor progression.
Materials and Methods
Tissue Specimens
Forty samples of human colorectal carcinomas with adjacent normal mucosa were obtained from the Pathology Department at Kφbenhavns Kommunes Hvidovre Hospital (University of Copenhagen; Hvidovre, Denmark). Four biopsies of well-differentiated carcinomas (10%), 25 biopsies of moderately differentiated carcinomas (62%), and 11 poorly differentiated carcinomas (28%) were collected. Four samples of tubular adenomatous polyps and three samples of normal mucosa taken at least 5 cm from the sites of the carcinomas were obtained from the Pathology Department (University of Maastricht; The Netherlands). Moreover, normal mucosa adjacent to tumors was analyzed. After resection, the tissue samples were immediately frozen in liquid nitrogen and stored at −80C until used.
Preparation of Cryostat Sections
Serial cryostat sections (6 μm thick) were cut at a cabinet temp of −25C on a motor-driven cryostat (Bright; Huntingdon, UK) at a low but constant speed to minimize variation in section thickness. The sections were picked up on clean glass slides and stored at −80C until used.
Histopathology
Pathological examination was performed with the use of hematoxylin–eosin (H&E)-stained sections of each specimen. For orientation in the sections and to facilitate analysis of serial sections, high-resolution composite images of entire H&E-stained sections of all biopsies were taken. Digital images were captured using a light microscope (BH2; Olympus, Tokyo, Japan) with a ×2 SPlan FL 2 objective and a CCD black-and-white video camera (Sony; ATV Horn, Aalen, Germany). The camera was coupled via a frame grabber (maximal size 786 × 512) to an image analysis system with the KS 300 software package (Kontron; Eching, Germany). Camera signal and set up were adjusted according to the recommendations of Chieco et al. (1994). Tissue sections were viewed with the use of white light. Furthermore, microphotographs were taken of the H&E-stained sections at higher magnification using a Vanox light microscope (Olympus) with ×10 and ×25 objectives. Consecutive sections were used for immunohistochemical and enzyme histochemical analysis of cathepsin B expression and cathepsin B activity, respectively.
Immunohistochemistry
A rabbit polyclonal antibody against both the pro-form and the active form of human cathepsin B (Calbiochem; Cambridge, UK), which recognizes an epitope of the heavy chain of cathepsin B, was applied to localize cathepsin B with the use of the immunogold-silver labeling technique. Sections were air-dried. Blocking of endogenous peroxidase activity was performed in methanol containing 0.45% hydrogen peroxide for 15 min. Sections were rinsed three times for 5 min in 50 mM Tris rinsing buffer containing 0.05% Tween-20 (Merck; Darmstadt, Germany). To reduce nonspecific binding, sections were rinsed in Tris buffer containing 3% fat-free milk powder, 0.1% fish gelatin, and 0.05% saponin (incubation buffer). The antibody was used at a concentration of 1 μg/ml. After rinsing in Tris buffer containing 0.05% (v/v) Tween-20, sections were incubated with the secondary anti-rabbit IgG conjugated with 0.8 nm colloidal gold (Aurion; Wageningen, The Netherlands) at a dilution of 1:30 in incubation buffer for 60 min at 20C. After rinsing in Tris buffer, additional stabilization of the immunogold complex was performed by a fixation step in 2% glutaraldehyde in Tris buffer for 15 min, followed by three rinses in bidistilled water for 5 min each. Sections were stained in the dark for 9 min in silver developer, freshly prepared according to Danscher and Nörgaard (1983). After rinsing in bidistilled water for 3 min, sections were fixed for 3 min in a solution of 2.5% (w/v) Na2S2O3 in bidistilled water and then stained in Gill's hematoxylin for 15 sec and in eosin for 30 sec before mounting in Euparal. For negative controls, the primary antibody was replaced by an irrelevant rabbit polyclonal antibody (against salmon vitellogenin) at identical dilutions.
Images of the immunohistochemically stained sections were taken with an Axioskop light microscope (Zeiss; Oberkochen, Germany) using epipolarized light for the visualization of the immunogold-silver reaction product and a ×40 water-immersion objective.
Enzyme Histochemistry
The incubation medium to demonstrate cathepsin B activity consisted of 100 mM phosphate buffer (pH 6.0), 1.3 mM EDTA (disodium salt), 1 mM dithiothreitol, 2.67 mM
The image analysis system has been described in detail by Jonker et al. (1997) and consisted of the following components: (a) a fluorescence microscope (Dialux 20; Leitz, Wetzlar, Germany) with a ×10 objective or a ×40 water-immersion objective and excitation at 385–425 nm and emission at >460 nm; (b) a CCD camera with an 8-bit resolution (4910; Cohu, San Diego, CA) to capture images; (c) a frame grabber (LG-3; Scion, Frederick, MD) to convert and transport images from the camera to the computer; (d) a Power Macintosh 8100/110 computer (Apple; Cupertino, CA) and the public domain NIH imaging software program (version 1.57; written by Wayne Rassband and available via internet from http://rsb.info.nih.gov) for processing and analyzing of the images; settings of camera and frame grabber were according to Jonker et al. (1997); and (e) two monitors to visualize images to directly compare images of protein expression and activity with H&E images.
Results
Localization of Cathepsin B Activity
Cathepsin B activity was present in all specimens (Figure 1a). Activity was optimally localized after 15 min of incubation. Incubations in the presence of FMK did not produce any yellow fluorescence, indicating the specificity of production of fluorescence by cathepsin B activity (Figure 1b).
Immunolocalization of Cathepsin B
The polyclonal antibody demonstrated cathepsin B in epithelium and stroma of colorectal mucosa (Figure 1d). Control reactions with an irrelevant rabbit antibody raised against salmon vitellogenin at the same dilution resulted in low levels of positivity (Figure 1f). Some-times, precipitate in the lumen of the colon and rectum reacted nonspecifically (Figures 1d and 2d). Positivity in the lumen was ignored in all further studies.
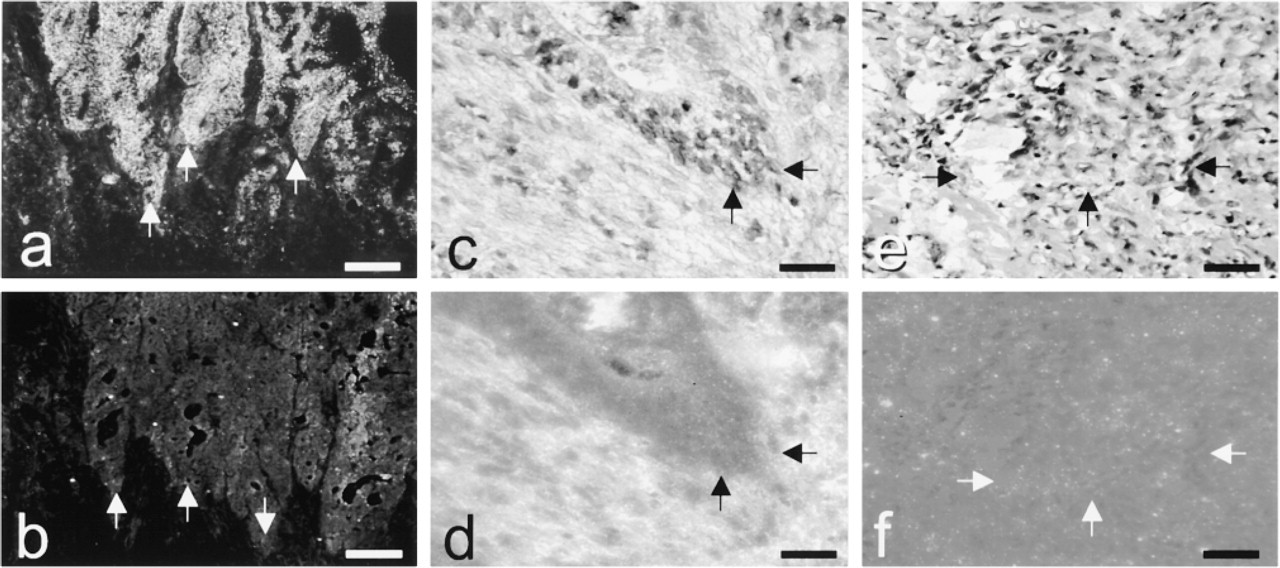
Digital photomicrographs of sections of invasive fronts (arrows) of a moderately differentiated carcinoma after staining for cathepsin B activity in (
Cathepsin B in Colorectal Cancer
Table 1 summarizes the semiquantitative analysis of localization patterns of cathepsin B protein and activity in relation to colorectal cancer.
Cathepsin B in Normal Colorectal Mucosa
Cathepsin B protein was present in variable amounts in epithelium of the crypts. In most cases, epithelium contained moderate amounts of cathepsin B protein (Figure 2f), but sometimes epithelium in crypts was almost devoid of cathepsin B (Figure 2d). Stroma also contained moderate amounts of cathepsin B that was homogeneously distributed (Figures 2d and 2f).
Normal colorectal epithelium showed cathepsin B activity that was heterogeneously localized. Activity was mainly observed in epithelium at the colorectal surface, whereas epithelium in the crypts showed hardly any activity (Figures 2b and 2c). Stroma was usually negative except for distinct areas that were mainly found directly underneath the colorectal surface (Figures 2b and 2c).
When the localization patterns of cathepsin B protein and activity are compared, it can be concluded that (older) epithelium at the colorectal surface expressed both cathepsin B protein and activity. When epithelium in the crypts contained cathepsin B protein, it was not active. Stroma contained moderate amounts of cathepsin B protein that was homogeneously distributed, but it was active only in certain areas that were mainly found directly underneath the colorectal surface (cf. Figures 2c and 2d). These localization patterns of cathepsin B protein and activity were similar in all normal mucosa investigated, whether or not it was adjacent to tumors.
Cathepsin B in Adenomas
Cathepsin B protein was mainly localized in tubule adenomas at the basal side of epithelium adjacent to the basement membrane (Figure 3b). This pattern was distinctly different from that in normal epithelium (cf. Figures 2d and 2f). The entire epithelium of all adenomas contained cathepsin B protein. Cathepsin B protein expression in stroma was similar to that in normal colorectal tissue. Activity was elevated in epithelium of all adenomas compared with normal epithelium (cf. Figures 2c and 3a). It was localized along its basal side. Stroma contained only little activity (Figure 3a).
When protein and activity distribution patterns were compared, we conclude that a large fraction of cathepsin B was localized in granules at the basal side of epithelium and that this fraction was active (Figures 3a and 3b). Cathepsin B in stroma did not show distinct alterations in adenomas compared with normal colorectal tissue.
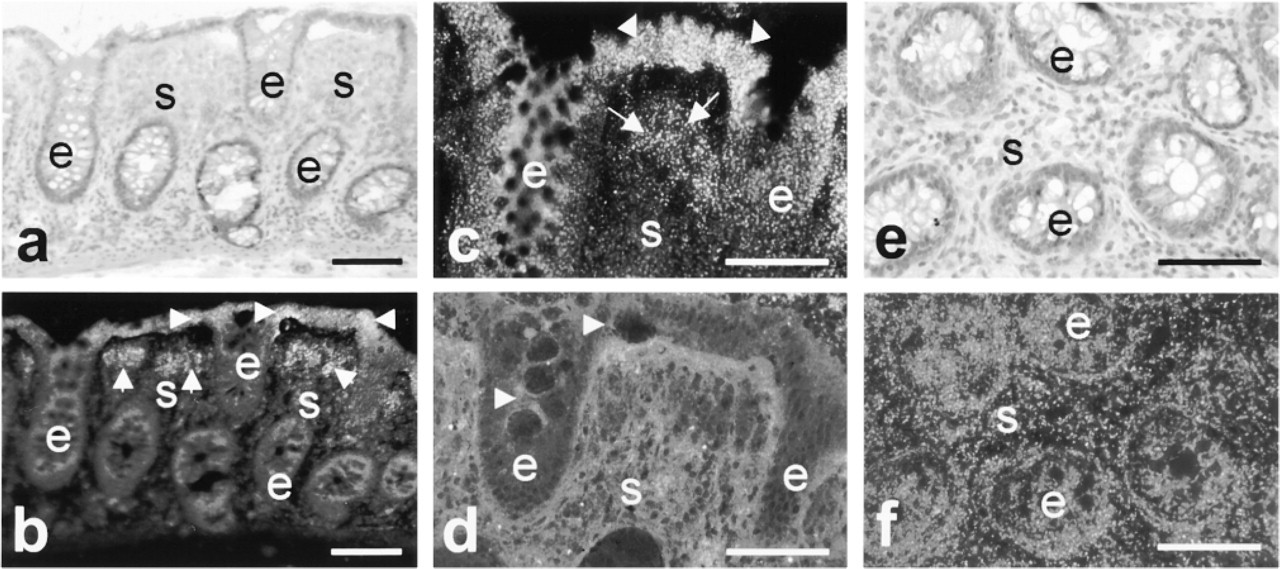
Digital photomicrographs of adjacent sections of normal human colon tissue stained with H&E (
Cathepsin B in Well-differentiated Carcinomas
Epithelium in well-differentiated tumors showed an expression pattern of cathepsin B protein and activity that was largely similar to that in adenomas, whereas stroma contained higher amounts of cathepsin B protein than in adenomas (Figures 3c and 3d). Furthermore, a transition was observed in this stage in specific sites of four tumors towards a localization of both protein and activity over the entire cytoplasm of cancer cells (Figures 3c and 3d), as was observed in half of moderately and in all poorly differentiated carcinomas (see below).
Comparison of the localization patterns of cathepsin B protein and activity showed that these were in agreement with each other, except that stroma contained large amounts of inactive cathepsin B.
Cathepsin B in Moderately Differentiated Carcinomas
Expression patterns of cathepsin B protein and activity in moderately differentiated carcinomas were similar to those in well-differentiated carcinomas (Figure 3e). Epithelial cells in 50% of the tumors contained cathepsin B protein (not shown) and activity at the basal side of cells (Figure 3e), and in 50% it was diffusely distributed throughout their cytoplasm with a tendency towards localization in apical regions of the cells (Figure 3f). Stroma was strongly positive for protein (not shown), but in most cases not for activity (Figure 3e). However, in specific sites of some tumors, stroma was strongly positive for both protein (not shown) and activity (Figure 3f), similarly to poorly differentiated carcinomas (see below).
Cathepsin B in Poorly Differentiated Carcinomas
Epithelium in poorly differentiated carcinomas contained low levels of cathepsin B protein that was diffusely distributed in cancer cells, whereas stroma contained high cathepsin B levels (Figure 3h). Activity was significantly decreased in cancer cells in poorly differentiated carcinomas compared with that in moderately and well-differentiated colorectal carcinomas (Figure 3g), except for satellite-like protrusions of cancer cells that showed high activity (Figure 4). When cathepsin B activity was present, it was diffusely distributed over the cytoplasm of the cancer cells.
Cathepsin B and Invasive Fronts, Angiogenesis, Necrosis, and Inflammation
Invasive fronts and satellites of cancer cells showed moderate protein levels of cathepsin B, but its activity was high (Figures 1 and 4).
When activity was present in stroma, it was in defined tissue compartments where angiogenesis (Figures 5a and 5b), necrosis (Figures 5c and 5d), or inflammatory cells (Figures 5e and 5f) were present. Lymph follicles were frequently observed, which did not show any activity.
Expression of cathepsin B protein as detected immunohistochemically and cathepsin B activity as detected enzyme histochemically in different tissue compartments of human colorectal mucosae and colorectal tumors; various tumor biopsies contained regions with epithelial cells in different stages of carcinogenesis, and therefore numbers of the different stages of cancer that were analyzed are greater than the numbers of biopsies that were available a
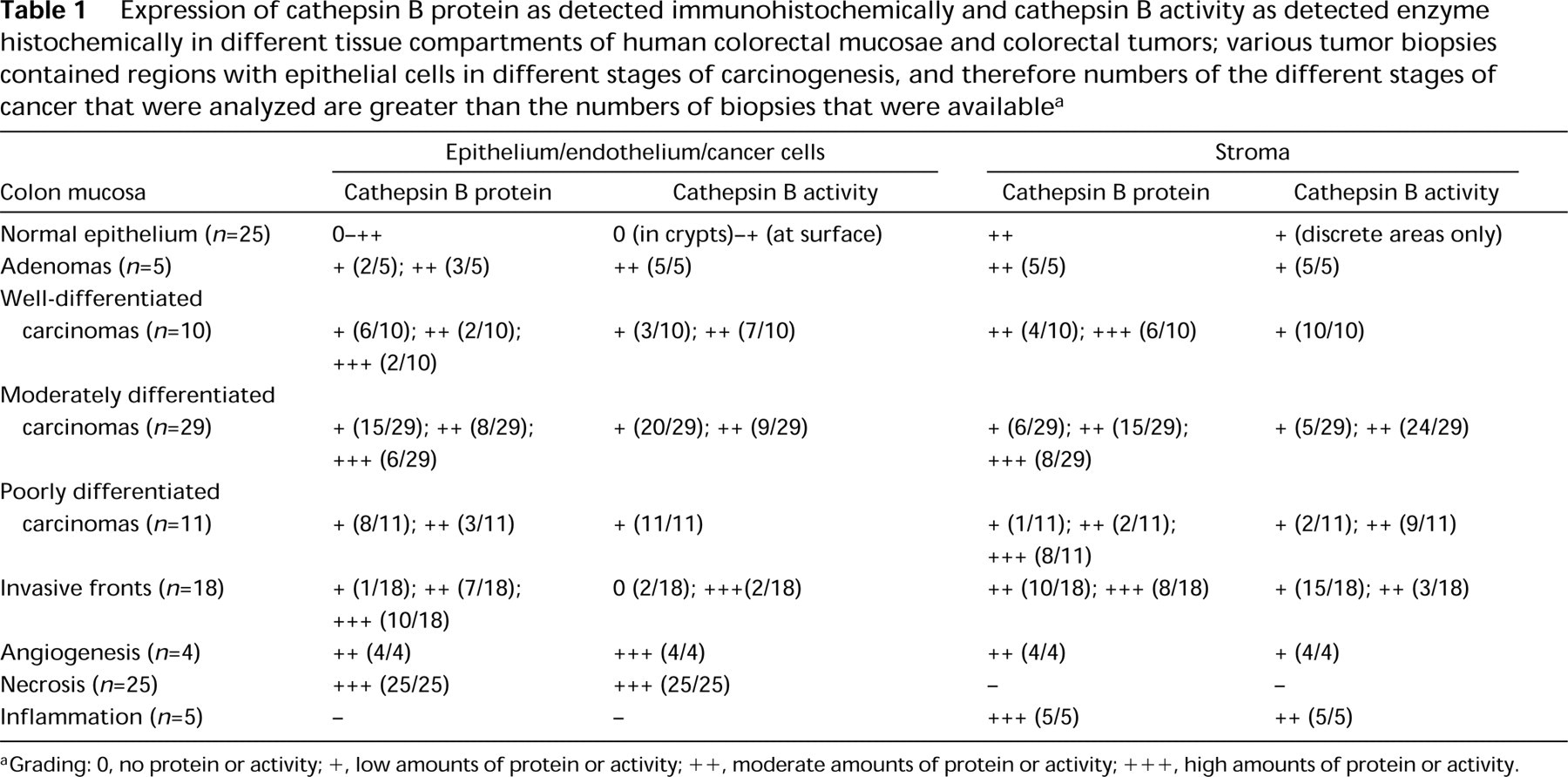
aGrading: 0, no protein or activity; +, low amounts of protein or activity; + +, moderate amounts of protein or activity; + + +, high amounts of protein or activity.
Discussion
Localization of cathepsin B protein and activity in serial sections revealed a distinct pattern in relation to colorectal carcinogenesis. In normal colorectal mucosa, cathepsin B protein was present in the larger part of the epithelium but was active only in the older cells at the colorectal surface (Bleeker et al. 2000). It may play a role in apoptosis in these cells as it does during regression of prostate and mammary glands (Guenette et al. 1994; Tenniswood 1997). In tubule adenomas, the intracellular localization pattern was changed significantly. The entire epithelium contained active cathepsin B. It was localized at the base of cells close to the basement membrane. Whether this localization pattern is a general phenomenon in all adenoma types remains unknown. However, this distribution pattern did not change in well-differentiated carcinomas and half of the moderately differentiated carcinomas. In a few cases (four) of well-differentiated, half of the moderately differentiated carcinomas, and all of the poorly differentiated carcinomas, the intracellular localization of protein and activity was diffuse over the cytoplasm. Similar distribution patterns were found by immunolocalization of cathepsin B by Keppler et al. (1988) and Campo et al. (1994). This diffuse form is either the truncated cytoplasmic form of cathepsin B (Campo et al. 1994; Qian et al. 1994; Keppler and Sloane 1996; Frosch et al. 1999) or a vesicular form (Frosch et al. 1999), or both. It appeared to be active in our study. These forms of cathepsin B may well be involved in extracellular proteolytic processes related to invasion, because all satellite-like protrusions and invasive fronts that contained high activity showed a diffuse distribution pattern of activity all over the cancer cells. It may well be the form that is secreted or built into the plasma membrane (Sloane et al. 1986; Erdel et al. 1990; Qian et al. 1994; Spiess et al. 1994; Keppler and Sloane 1996; Strohmaier et al. 1997; Ulbricht et al. 1996, 1997; Van Noorden et al. 1998a; Frosch et al. 1999). Biopsies of poorly differentiated carcinomas were characterized by low cathepsin B protein and activity in cancer cells, except for satellite-like protrusions that contained cathepsin B protein but particularly high levels of cathepsin B activity.
Stroma also showed distinct changes in cathepsin B expression during tumor progression. It contained higher amounts of cathepsin B protein in malignant tumors than in adenomas and normal colonic mucosa. In general, stromal cathepsin B protein expression was highest in poorly differentiated carcinomas. Whereas cathepsin B protein is present rather homogeneously in stroma, cathepsin B activity is localized in distinct patterns throughout tumor progression from normal mucosa to poorly differentiated carcinomas. Activity in stroma was particularly linked with angiogenesis, necrosis, and inflammation.
Our study links previous seemingly contradictory studies of cathepsin B in colorectal cancer. We found the inverse relationship between differentiation grade and cathepsin B activity that Murnane and co-workers (Sheahan et al. 1989; Murnane et al. 1991; Shuja et al. 1991; Iacobuzio–Donahue et al. 1997; Del Re et al. 2000) found in homogenates. A disadvantage of the biochemical analysis was the use of homogenized tissues in which the exact localization of cathepsin B activity was lost. Undoubtedly the tissues contained stroma that contains highly variable cathepsin B activity. High cathepsin B activity was found particularly in areas of invasion, angiogenesis, necrosis, or inflammation. Angiogenesis can be considered as a controlled invasive process in which endothelial cells produce a number of proteolytic enzymes, such as cathepsin B (Keppler et al. 1996). Cathepsin B is an important enzyme in inflammatory cells, such as macrophages, and is involved in breakdown of inflamed tissue (Reddy et al. 1995). Our findings that high cathepsin B activity in tumors is related to necrosis suggest that cathepsin B is activated in necrosis as part of autolysis, probably in a similar way as it is activated in apoptotic cells in normal epithelium.
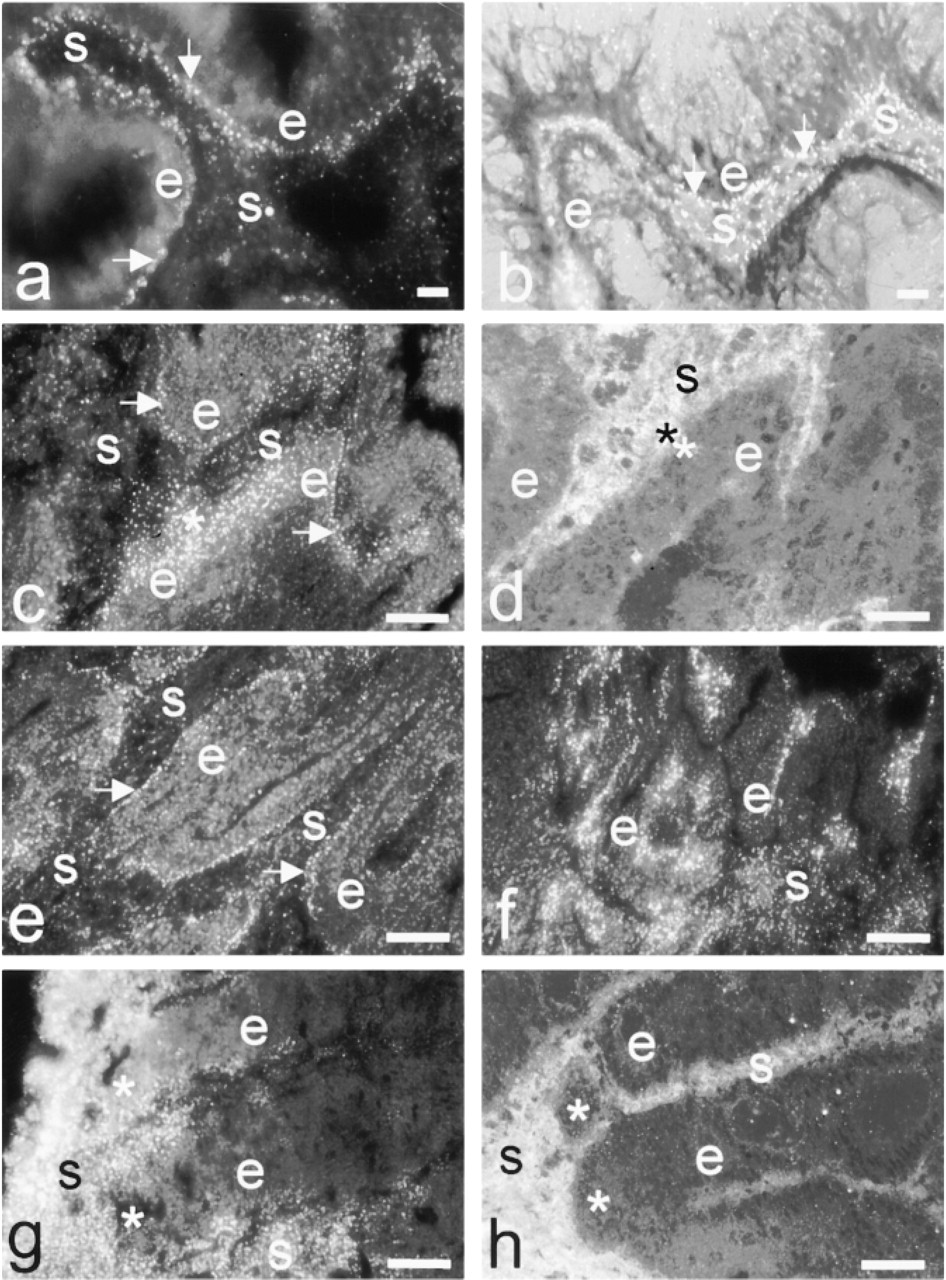
Digital photomicrographs of adjacent sections showing (
The high cathepsin B activity in invasive fronts of the tumors correlates well with the link between cathepsin B protein expression and shortened survival as was found by Sloane and co-workers (Campo et al. 1994; Emmert–Buck et al. 1994) using immunohistochemistry or microdissection and subsequent activity measurements. A relationship between cathepsin B and cathepsin L levels and survival of breast cancer patients was reported recently by Foekens et al. (1998) but not by Lah et al. (2000). Prognostic significance of cathepsin B levels in colorectal cancer was not found despite the link between cathepsin B protein expression and metastasis (Herszenyi et al. 1999; Hirai et al. 1999).
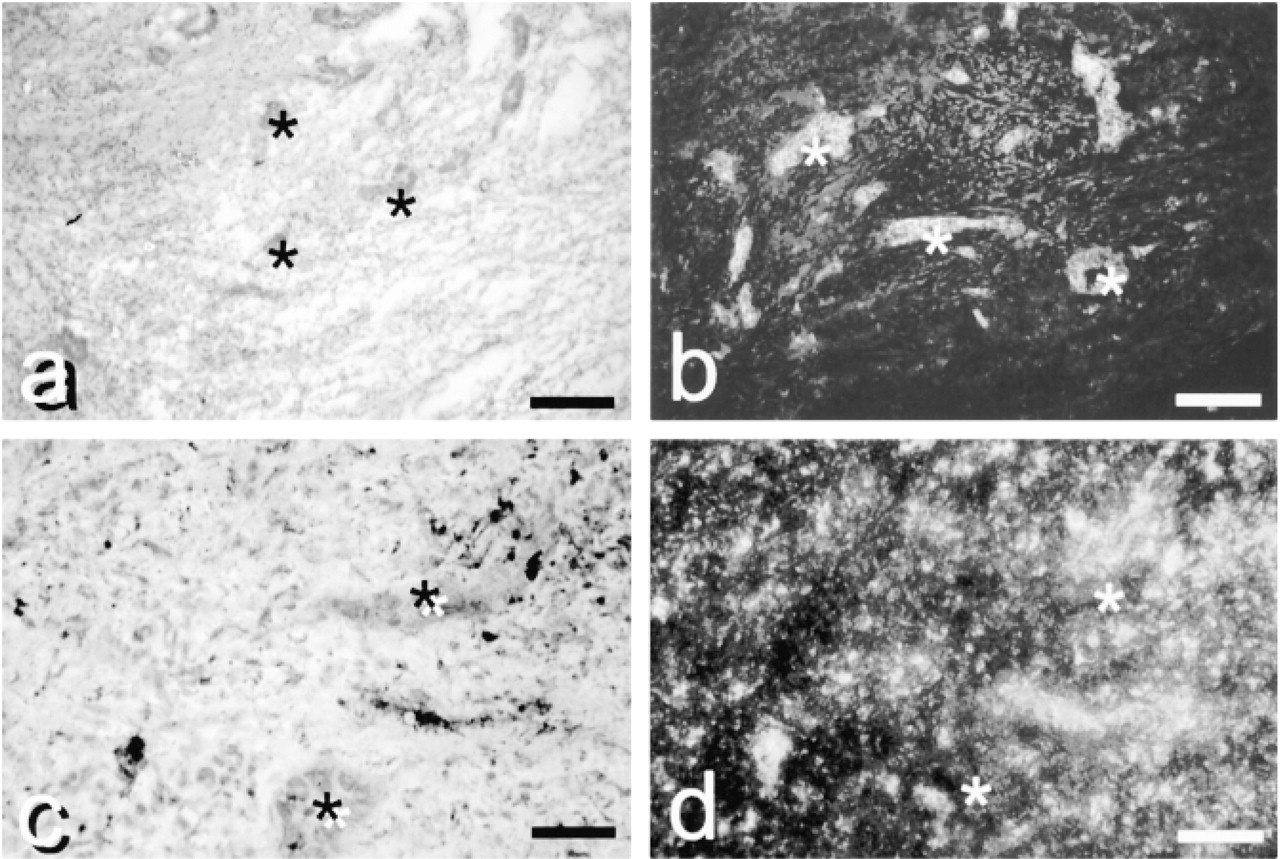
Digita photomicrographs of invasive satellite-like fronts (asterisks) in a moderately differentiated carcinoma stained with H&E (
In conclusion, our study combining localization of protein and activity of cathepsin B indicates that cathepsin B expression in colorectal cancer shows a distinct pattern. Its activity is markedly linked to processes such as apoptosis in normal epithelium and necrosis in tumors, and to invasion of cancer cells, angiogenesis, and inflammation. In particular, the link with invasion and angiogenesis and the fact that cathepsin B can be expressed extracellularly in these processes make the enzyme an attractive target for therapeutic strategies using selective inhibitors of the extracellular form of cathepsin B. In this way, the enzyme in the lysosomes is kept intact for its normal function in protein degradation. This approach was effective in a rat model of colon cancer metastasis in the liver, in which oral treatment of the rats with a water-soluble selective inhibitor of cathepsin B reduced metastasis (Van Noorden et al. 1998a).
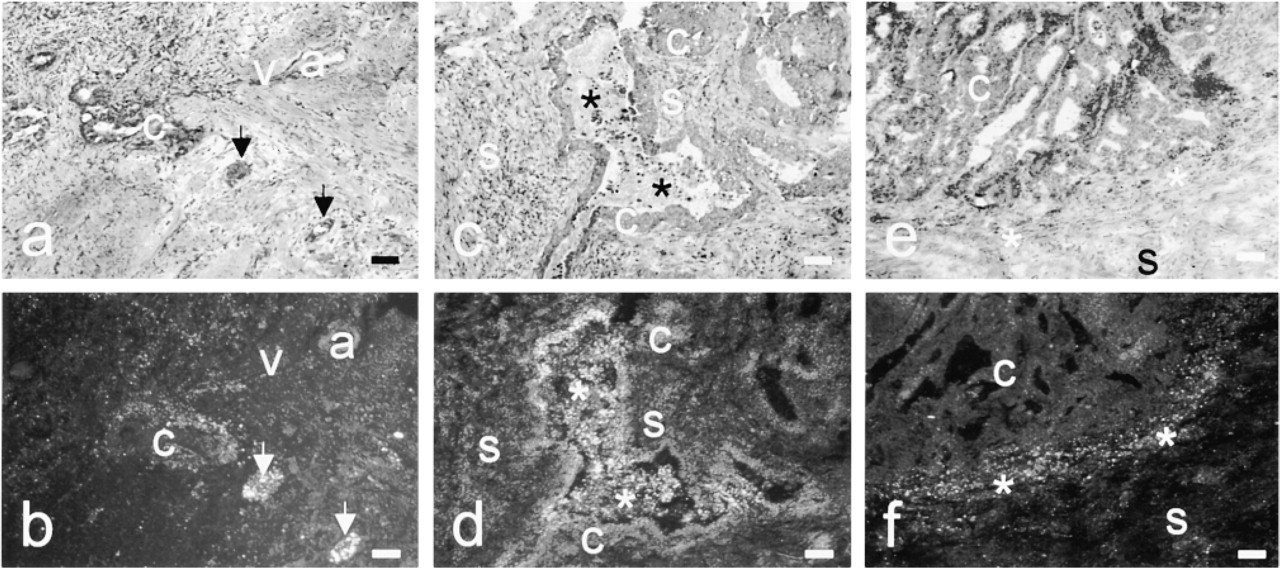
Digital photomicrographs of serial sections showing angiogenesis (arrows) in stroma around poorly differentiated colon carcinoma cells stained with H&E (
Footnotes
Acknowledgements
We are grateful to Mr Jan Peeterse for preparation of the digital images and to Ms Trees M.S. Pierik for preparation of the manuscript.